 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 3, July 15, 2007, pg. 417-424 RESEARCH ARTICLE Induction of in vitro roots cultures of Thypha latifolia and Scirpus americanus and study of their capacity to remove heavy metals María del Socorro Santos-Díaz*1,María del Carmen Barrón-Cruz2 , María Catalina Alfaro-De la Torre3 1Centro de Investigación y Estudios de Posgrado
Facultad de Ciencias Químicas Universidad Autónoma de San Luis
Potosí Manuel Nava 6, San Luis Potosí CP 78210, México
Tel: 444 8262440 Fax: 444 8262372 E-mail: ssantos@uaslp.mx *Corresponding author Financial support: Code Number: ej07040 We have established the conditions to obtain in vitro root
cultures of Thypha latifolia and Scirpus americanus and have
investigated their capacity to remove Pb(II), Mn(II) and Cr(III) from the culture
medium. The best conditions for the in vitro culture growth were: an
inoculum of Keywords: heavy metals, in vitro root cultures, phytoremediation, Scirpus americanus, Thypha latifolia. Abbreviations: 2,4-D: 2,4-dichlorophenoxiacetic acid Heavy metals are toxic pollutants that have serious adverse effects on human health. They are toxic because can replace other essential metals in pigments or enzymes, disrupting the function of these molecules (Manios et al. 2003). Also because they may cause oxidative stress, especially transition metals as Fe2+/3+ and Cu+/2+ (Rivetta et al. 1997). The removal of metals from solution using plants offers an attractive alternative, because it is solar driven and can be carried out in situ, minimizing cost and human exposure (McCutcheon and Schnoor, 2003). Plants have developed different mechanisms of tolerance to the metals and to the metal accumulation. Some plants excrete organic acids, as malate and citrate, that act as metal chelators and decrease the rhizospheric pH increasing the bioavailability of metals for phytoextraction (Pivetz, 2001). Organic acids can also inhibit metal uptake because they complex the metal outside the root (phytostabilization). For example, citrate inhibits Al and Cu uptake in some plant species (De la Fuente et al. 1997; Murphy et al. 1999). The high metal tolerance may be in part due to the highly efficient intracellular compartmentalization. The uptake of metals requires their transport across the root cell membrane to the symplast. This process involves specific membrane transporter proteins as heavy metal ATPases (HMAs), the natural resistance-associated macrophage proteins (Nramps), and cation diffusion facilitators (CDFs) (Hall and Williams, 2003). Once inside the cell, the metals could be translocated to their final destination by membrane metal transporters, and metal-binding proteins as metallothioneins (Goldsbrough, 2000). Metal chaperones are a different class of proteins that bring metals to specific targets in the cell (Himelblau et al. 1998). For the storage in the vacuole (sequestration), certain metals may be complexed by phytochelatins. These compounds are synthesized enzymatically from glutathione. Complex of metals bound by glutahione or phytochelatins are shuttled to the vacuole by an ATP-binding cassette (ABC) type transporter protein in the tonoplast. Other metal-binding molecules that are involved in the metal complexation in the vacuole are organic acids (Kramer et al. 2000) and sulphides, in the particular case of cadmium (Cobbett, 2000). Additional mechanisms of tolerance are the assimilation of metals into organic molecules by metal-modifying enzymes (e.g. selenate is metabolized to dimethylselenide) or the changes in the oxidation state of metals that reduce their toxicity (e.g. toxic Cr VI is reduced to the non toxic Cr III; Lytle et al. 1998; De Souza et al. 2000). The plants are efficient in the removal metals from their solid or liquid environment as also the plant cell cultures do. In vitro culture of plant organs (roots and shoots) allow indefinite propagation and experimentation using tissues derived from the same plant, thus avoiding the effects of variability between individual specimens. Axenic conditions in culture prevent microbial symbiosis disguising the metal uptake characteristics of plants grown in soil. Experiments using separately cultured organs also allow the metal accumulation properties of each organ to be identified. Some examples include the removal of Sr2+ by shoots of Solanum laciniatum (Kartosentono et al. 2001), and hyper-accumulation of Cd by hairy roots of Thlaspi caerulescens (Nedelkoska and Doran, 2000). Another advantages of in vitro cultures is the facility to obtain variants with different tolerance to several biotic stresses (Ben-Hayyim, 1987; Santos-Díaz and Ochoa-Alejo, 1994). Some plants that have potential on phytoremediation are Thypha latifolia (reed mace) and Scirpus americanus (american tule). These species are able to grow on water bodies contaminated with heavy metals formed as result of industrial and domestic effluents (Montante-Montelongo et al. 1995) or in sludge compost watered with metaliferus water (Manios et al. 2003). This work is focused on T. latifolia and S. americanus plants
growing on an artificial lake contaminated with Pb, Cr, Fe, Ni, Cu and Mn,
known as Tenorio Tank. This tank has an extension of On the basis of this observations, the aim of this research was to develop a protocol to establish the in vitro roots cultures of T. latifolia and S. americanus and later on to investigate if they have the capacity to remove Pb(II), Mn(II) and Cr(III). Their concentrations are the most important in the water column of Tenorio Tank. Because phytoremediation and phytomining requires growth of the plants, the experiments were carried out in nutrient media under conditions supporting the root growth. The T. latifolia and S. americanus roots were
collected from plants growing on the Tenorio Tank. Segments of roots (5 to The roots were cultured in 250 ml flasks on a orbital shaker
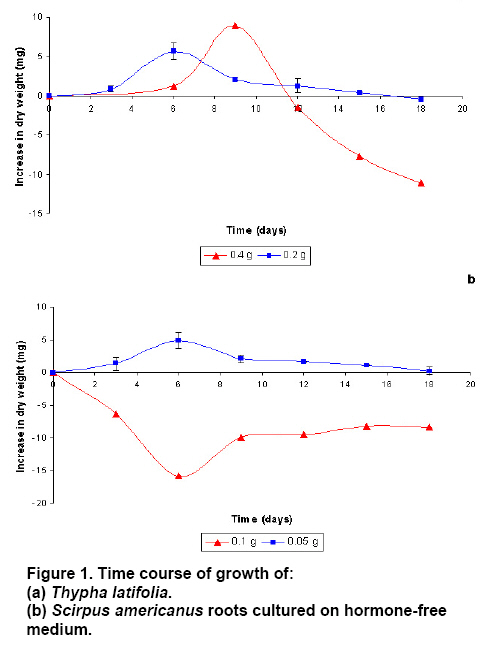
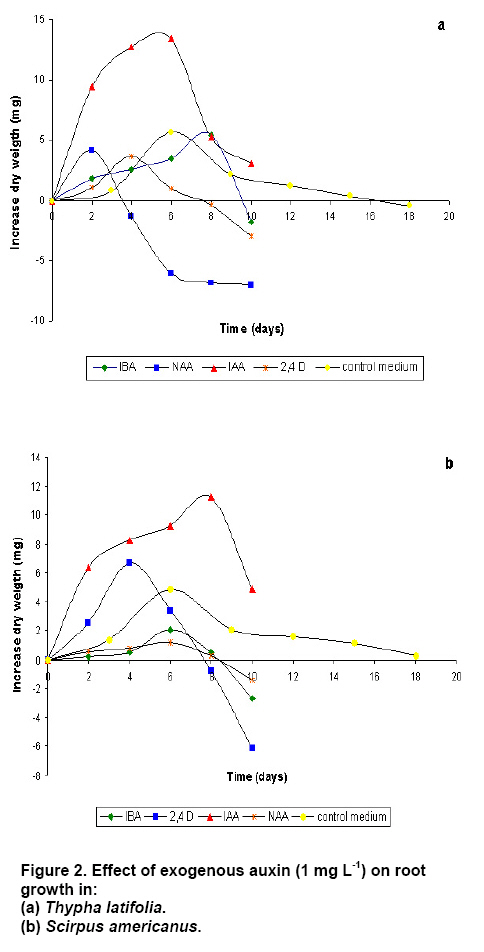
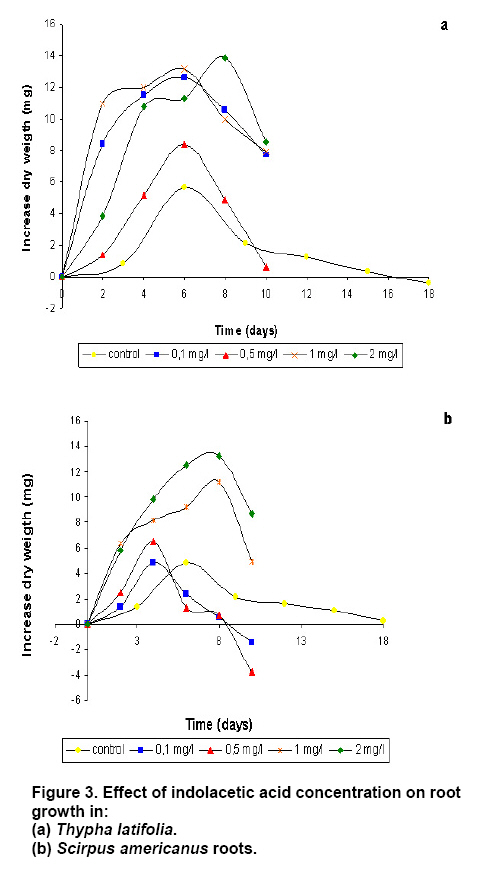
operated at 130 rpm and maintained at Three replications per treatment and two independent experiments were included per analysis. In order to evaluate statistically and significant differences among mean values, an ANOVA and Tukey test were used at the significance level of 95%. To establish the in vitro root cultures As the auxins generally promote lateral root formation, we tested the effect of 1 mg L-1 IAA, IBA, NAA or 2,4-D. After 10 days, the growth of roots of T. latifolia onto the culture media containing NAA or IBA was lower in comparison with control medium. An increase in biomass was observed until day 4 onto the medium with 2,4-D. The IAA promoted the highest growth of roots reaching the maximum at day 8 (Figure 2a). The roots of S. americanus presented a similar behaviour. The growth of roots was lower on media with ANA and 2,4-D in comparison with control medium. The highest increase in growth was obtained again with IAA but at day 6 (Figure 2b). Once selected the auxin promoting the most important increase of growth, we analyzed the effects of different concentration of IAA on rooting. As expected, the exogenous IAA caused abundant lateral roots formation in T. latifolia compared with control medium. The increase in radical growth was proportional to the concentration of auxin until 8 day (Figure 3a). In the case of S. americanus roots a small increase in dry weight was observed with 0.1 and 0.5 mg L-1of IAA but just until day 4. However, using higher concentration of IAA (1-2 mg L-1) an increase between 3 to 3.5 fold was observed in comparison to control medium (Figure 3b). In summary, the optimal conditions to establish the in
vitro roots culture corresponded to an inoculum of To measure the Pb, Cr and Mn removalby roots the experiments
were conducted over a period of 8 days. The results shown that growth of T. latifolia and S.
americanus roots was essentially unaffected by the presence of Pb, Cr or
Mn onto medium (Table 1). The S. americanus and T. latifolia roots
removed from the culture medium about 50% of Pb at 4 hrs, period that was nominated
time 0. The metal was totally captivated at day 4 by S. americanus cultures
and about 95% at day 8 by T. latifolia roots. In the case of Cr its
removal was slower reaching only 34% (T. latifolia) and 47% (S. americanus) at
time 0. Close of 90% of metal was removed from the medium at day
Table 2 shows the maximum removal of metal from the solution related to the quantity of root biomass (gram of dried weight) that has growth in the culture medium. According to these data S. americanus cultures removed between 1.5-3 fold more metals than T. latifolia roots and in a shorter time.
The in vitro roots culture is an ideal system to study the process involved in heavy metals uptake because they can be propagated indefinitely, are very stable, and eliminate the effect of variations between individual seedlings (Pollard and Baker, 1996). Previous papers have demonstrated the effectiveness of in vitro roots cultures to remove metals (Nedelkoska and Doran, 2000). Yet, the metal concentrations in biomass grown in liquid culture are usually greater than those in soil-grown plants due to the greater bioavailability of metal ions in solution. In this work, we selected the T. latifolia and S.
americanus species since they are adapted to grow on presence of heavy
metals. To initiate the in vitro culture the roots were cultivated
in media with auxins because it is widely accepted that auxins have a central
role in adventitious root initiation (Blakesley, 1994).
In T. latifolia and S. americanus root cultures the auxin IAA
promoted the highest root formation. As it is a natural endogenous hormone
probably it was rapidly catabolised to a suitable level in cells under strict
metabolic regulation (Vuylsteker et al. 1998). It has been
described that IAA improves the number and quality of roots in several species
because it stimulates the cellular division of radical primordium, promotes
the synthesis of specific proteins and increases the sugar release into the
phloem ( The T. Latifolia and S. americanus root cultures were able to remove Pb and Cr near to 100% and 71-100% of Mn from the medium solution during the 6-8 days of experimentation.The mechanisms of metals removal seems to be characterized by a rapid then by a slower metal concentration decrease in the solution. The rapid decrease could be related to a sorption process at the root surface. Plant cells have an abundance of negatively charged-sites on their walls, so ion-exchange and other interactions between metals and carboxyl, sulphate, amino and other groups are likely to occur (Kratochvil and Volesky, 1998). The sorption of Pb, Mn and Zn to the root surface has been described in wetland plant (Phalaris arundinacea) by microtomography and X-ray microprobe images (Hansel et al. 2001). Adsorption and precipitation at the root surfaces seems to be the mechanism. Other mechanism of short-term removal, such a chelation and covalent binding cannot be discounted. The slower metal concentration decreasing can be due to an absorption process that should involve transport across the biological membrane and the internalization of the metal. This response could be a critical defensive strategy of the plant, providing time for the development of intracellular mechanism of metal complexation with specific proteins or sequestration onto the vacuole. In the particular case of Thlaspi caerulescens the analysis of root cell wall fractions revealed that the hairy roots partitioned virtually all the Cd uptake by the biomass in the cell wall fraction for 7 to 10 days before allowing passage into the symplast (Nedelkoska and Doran, 2000). The mechanism of Cd hyperaccumualtion in this species seems to be related to an efficient antioxidative defense, particularly an enhanced catalase activity (Boominathan and Doran, 2003). Further studies will be required to define the specific mechanisms of tolerance to Pb, Cr and Mn in S. americanus root cultures. It will be necessary to determine if the metal removal is the result of an adsorption and/or absorption process and which molecules are participating (glutathion, glutathion S-transferase, organic acid, fitoquelatins, methalothioneins, etc.). On the other hand, the plants have been divided in three arbitrary categories according to the ability to remove metals: normal plants, accumulators and hyperaccumulators. In the particular case for Cr, Pb and Mn metals, an accumulator concentrates more than 50 µg Cr g-1, 100 µg Pb g-1 and 2000 µg Mn g-1 dry weight while an hyperaccumulator concentrates 1000 µg g-1 of Cr and Pb, and 10,000 µg g-1 of Mn (Reeves and Baker, 2000). Considering the quantity of roots used and the metal removed from solution we have calculated the metal concentration removed per gram of tissue. Taking into account these data, the T. latifolia species correspond to the category of normal plants and S. americanus could be considered as an accumulator for Pb (157.73 µg g-1), Cr (55.6 µg g-1) and Mn (5000 µg g-1). To date there are few reports of plants that accumulate Mn but there are no known Pb hyperaccumulator plants (Lasat, 2002). Even when the in vitro root cultures of S. americanus were not a hyperaccumulator it is an interesting model due to its tolerance to several metals. It has been described that plants able to accumulate more than one metal, present some mechanisms that includes the induction of specific enzymes or phytochelatins, and the participation of specific transporters. For example, phytochelatins complexes with Cd, Ag and Cu has been identified on Arabidopsis thaliana, Silene vulgaris, Holcus lanauts, Agrostis castelana and Thlaspi caerulescens (Cobbett, 2000; Schat et al. 2002). The enhanced tolerance to Cd, Co, Cu, Mg, Ni, Pb and Zn in transgenic tomato plants was related to an increase in 1-aminocyclopropane-1-carboxilic acid (ACC) deaminase activity (Grichko et al. 2000) and the tolerance to Cd, Co and Zn in Thlaspi goesingense is related to a CDF present in vacuoles (Hall and Williams, 2003). The root cultures present an additional advantage to study the mechanisms of tolerance to heavy metals. It is well known that the in vitro production of secondary metabolites can be increased several times by modifying the culture conditions (Charlwood et al. 1990). This strategy could also be used to increase the metal tolerance on S. americanus root cultures. There are many examples from the literature describing plants which synthesize and accumulate secondary metabolites (glyceollins) upon treatment with metals. The mechanism proposed suggests that heavy metals induce an oxidative stress. The lipid oxidation process generate oxilipins, signaling molecules responsible for the heavy metal-induced defense response, this is, enzymes, glyceollins, proteins, transporters and/or quelators (Mithöfer et al. 2004).
Note: Electronic Journal of Biotechnology is not responsible
if on-line references cited on manuscripts are not available any more after
the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07040f3.jpg] [ej07040f1.jpg] [ej07040f2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}