 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 3, July 15, 2007, pg.436-443 RESEARCH ARTICLE Molecular description and similarity relationships among native germplasm potatoes (Solanum tuberosum ssp. tuberosum L.) using morphological data and AFLP markers Jaime Solano Solis1,Daniza Morales Ulloa2 , Leonardo Anabalón Rodríguez*3 1 Laboratorio de Biotecnología y Mejoramiento
Vegetal Escuela de Agronomía Universidad Católica de Temuco Longitudinal
Norte s/n Temuco, Chile Tel: 56 45 205527 Fax: 56 45 205540 E-mail: jsolano@uct.cl *Corresponding author Financial support: The present work has been done in the framework of a research programme DIPUCT (Grant DIPUCT 2004-2-02) of the Universidad Católica de Temuco. Code Number: ej07042 Chile is considered to be a sub-center of origin for the cultivated potato, with native and introduced genetic material coexisting in the country. Thus, the different varieties present in Chiloe Island are characterized by a rich diversity of forms, sizes, colours and phenological characteristics. In the present work, the level of polymorphism and the genetic relationship were studied by means of molecular markers using the amplified fragment length polymorphism (AFLP) technique and twenty-seven morphological characters. Twenty varieties of potatoes from the Chiloe Island were analyzed. The commercial variety Desirée and one specie from the Etuberosa series, Solanum fernandezianum, collected in the Juan Fernandez Island were included as controls. A similarity tree-diagram was made, based on all the AFLP bands generated in the range between 65 and 290 base pairs. With these tools, it was possible to identify molecular differences and similarities that might be associated with important morphological traits such as the predominant forms of the tuber, flower colour and resistance to disease. Keywords: cluster analysis, molecular markers, Solanum tuberosum ssp. tuberosum. Abbreviations: AFLP: amplified fragment length polymorphism The potato is one of the four most important crops in the country and constitutes a basic food in the diet of the population. Chile is considered to be a sub-center of origin for the cultivated potato (Spooner et al. 2005b). In this context, the native potatoes of Chiloe are characterized by a rich variety of forms, sizes, colours and phenological characteristics. This rich genetic patrimony needs to be described and individualized in order to be conserved and used. Unfortunately, more of the native potatoes of Chiloe are being lost every day, because of their replacement by introduced varieties and their phytopathological deterioration among other causes. As a result, their conservation must be ensured by means of germplasm banks collections. The conservations and use of this native genetic material will ensure that it does not disappear, but enjoys a projection for the future with new uses. In addition, potato improvement programs require a basic knowledge of the morphology and genetic nature of the main parts of the plant. In this context, new advances in molecular biology with the use of more sensitive molecular markers able to detect changes in the genotype of the individuals, greatly contributed to the generation of this important information. There is a remarkable concentration of cultivated and wild forms of potato in the south of Chile with the greatest number of native varieties being located in the island of Chiloe, and they are still preserved in the fields of small farmers. The peculiar characteristics of Chiloe, its natural conditions and its isolation, have allowed the proliferation of a great number of native varieties, of varying qualities and suitable for cultivation at different times in the farming calendar, as well as a number of different forms of preparation and consumption (Contreras et al. 1981). Until the mid 1960s, the markers used in genetic studies and improvement were those controlled by genes associated with morphological characteristics in easily identified phenotypes (Hijmans and Spooner, 2001). Potato germplasm has been described using morphological elements (Huamán et al. 1977; Ortiz and Huamán, 1994). Since 1975, it has been known in Europe and North America that potato cultivars could be identified by their protein and enzyme patterns (Stegemann and Loeschecke, 1976). With the modern technologies provided by molecular biology, various methods arise for the direct detection of genetic polymorphism at the DNA level using molecular markers. Molecular markers have contributed to a greater genetic knowledge of many vegetable species, including potato (Ritter et al. 2004). In addition, these markers have been used in the Solanum genus for the analysis of biodiversity and for phylogenetic studies (Ritter, 2000; Spooner et al. 2005a). Recently, efforts have been devoted to achieve a less subjective description of cultivars, based on Random Amplified Polymorphic DNA (RAPD) (Miller and Spooner, 1999; Isenegger et al. 2001; Sun et al. 2003), and microsatellites (Ashkenazi et al. 2001; Raker and Spooner, 2002). Interrupted Simple Sequence Repeats (ISSR) has also been used (Milbourne et al. 1998; Bornet et al. 2002). Amplified fragment length polymorphism (AFLP) analysis represents the most recent technology for obtaining a great number of molecular markers in the genomes of prokaryotes and eukaryotes (Savelkoul et al. 1999). The AFLP technique (Vos et al. 1995) generally produces between 50 and 100 scorable fragments per polimerase chain reaction (PCR) reaction (Maughan et al. 1996). This technique has been widely used in various crops, including cultivated potatoes (Powell et al. 1996; Milbourne et al. 1997; McGregor et al. 2000; Avrova et al. 2002; Straadt and Rasmussen, 2003; Furini and Wunder, 2004) and their wild relatives (Kardolus et al. 1998). In the last years, the AFLP technology was used to differentiate genotypes used as ancestors in potato genetic improvement programs. This allowed establish exactly the differences between the most important varieties used in these programs. In this context, Spooner et al. (2005b) used AFLP molecular data, suggest a single origin rather than multiple origins of cultivated potato. In this paper, we have analyzed the molecular diversity of Chilean native potato germplasm (Solanum tuberosum ssp. tuberosum L.) using AFLP markers and determined the genetic relationships existing among the genotypes included in the study. The analysis included twenty accessions of native potato collected on the island of Chiloe, one improved cultivar Desirée, and one accession from the Etuberosa series, Solanum fernandezianum collected in the Juan Fernandez Island, Chile. Accessions were chosen to represent the extensive genetic diversity that can be found in Solanum tuberosum (Table 1). This material was cultivated in the field during agricultural season 2003/2004. The culture in the field began October 2003 and ended with the harvest in May 2004. The varieties were planted to 80 cm between and 40 cm over the crop line. An experimental design of complete blocks in random order was used in the establishment of the varieties in the field, in which the treatments corresponded to the different varieties being evaluated.
Morphological characters analysis Twenty-seven characters, analyzed and described by Huamán et al. (1977) using potatoes varieties, and applied to “Chilean or native” varieties were numerically codified using a qualitative multi-status criteria (from 0 to 9, depending on the variables of each character). These were used to design a numbered-data matrix. Some characters included in this analysis were (Table 2): predominant tuber skin colour, general tuber shape, leaf characters, flower and fruit characters, phenologic data and reaction to fungi (Phytophtora infestans). Ten individuals were morphologically evaluated in each variety.
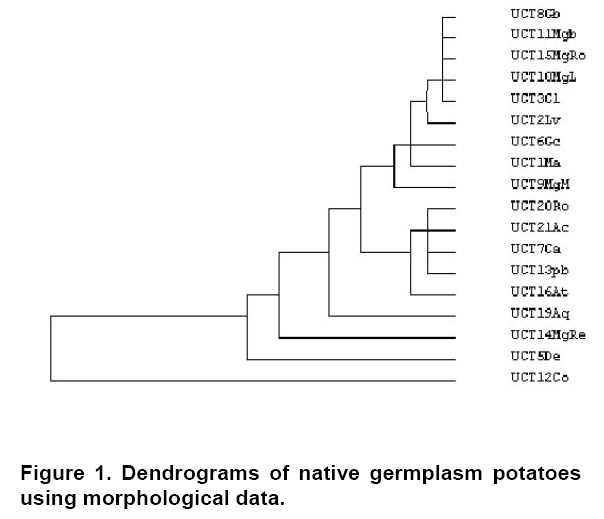
The morphologic data were subjected to cluster analysis. A standardization of the data by means of the Z-scores function was made. Average was applied to the technique of hierarchical conglomerate linkage using a matrix of quadratic similarity by means of Euclidean distances. The clustering method was made using the statistical software SPSS (statistical product and service solutions) version 12.0 for windows. S. fernandezianum was not included since it is a non-bearing species. Bruja, Michuñe negra and Clavela morada were not included because none of them bloomed and therefore several characters related to flowering and fructification could not be sufficiently registered. Approximately 100-200 mg of leaf tissue were freeze-dried and ground in liquid nitrogen with a mortar and pestle. Genomic DNA was isolated with Plant DNAzol® following the manufacturer's instructions. RNA was further eliminated by treatment with RNase. The quality and concentration of DNA was evaluated by agarose gel electrophoresis and spectrophotometry. Two independent extractions were performed on each accession. AFLP reactions were carried out using the AFLP Analysis System I kit (Invitrogen Life Technologies) according to the manufacturer's instructions. Each reaction was repeated at least once to verify the AFLP patterns generated. Approximately 500 ng of genomic DNA was digested for 2 hrs at 37ºC using 2 ul Eco RI/MseI restriction enzyme solution. The AFLP procedure (Vos et al. 1995) was carried out as described by Arens et al. (1998) with slight modifications. Briefly, the entire genomic DNA (400-500 ng) was digested with EcoRI and MseI, followed by ligation of the adapters. Pre-amplification was performed using a single adenine (A) selective nucleotide for each primer. For selective amplification, an EcoRI primer, with three selective nucleotides, was used in combination with a MseI primer with three selective nucleotides. For both the pre-amplification and selective amplification the following amplification profile was used: an initial cycle of 94ºC for 30 sec, 65ºC for 30 sec, 72ºC for 1 min, followed by 12 touchdown cycles in which the annealing temperature was reduced by 0.7ºC per cycle. The annealing temperature was then kept constant at 56ºC for the remaining 23 cycles. Amplification products were separated on a 6% polyacrylamide gel, and made visible by silver staining. Twenty-three primer combinations were tested for their ability to generate reproducible AFLP profiles that could be scored unambiguously. Reproducibility of the primer combinations was tested by comparing the AFLP profiles of four DNA samples collected from the same individual. Five combinations were specifically chosen for their ability to generate a large number of bands in order to increase resolution for the identification of possible identical plants (Table 2). For each primer combination, the presence or absence of a band in each sample was visually scored. Data were set in a binary matrix (Paul et al. 1997; Yee et al. 1999). Genetic similarities were calculated using the similarity coefficient and tree-diagrams obtained by clustering according to the unweighted pair group method with arithmetic averages (UPGMA) method using the NTSYS (numerical taxonomy and multivariate analysis system) pc 2.0.1 program (Applied Biostatistics Inc., NY, USA). The correspondence between the morphological and AFLP similarity coefficient matrices was tested on the basis of correlation analysis for Mantel test using the Mxcomp procedure. The morphologic data cluster analysis allowed the separation of two defined groups (Figure 1). The first group included potatoes with semi late vegetative cycle and elongate tubers, and skins of coloration dark purple. The stems of this group present angular sections and green colours and node of reddish and purples colours. The flowers show an intense red colour. Some varieties included in this are UCT8Gb, UCT11Mgb, UCT15MgRo, UCT6Gc, UCT9MgM and UCT1Ma. The second group presented semi early cycle and tubers of round forms, with skins principally purple colour and secondary white colour, distributed through all tuber or located around the eyes. This material presented dark green stems of angular sections and flowers intensely purple. This group was formed by the accessions UCT7Ca, UCT13MpB, UCT20Ro, UCT21Ac and UCT16At. Four accessions were not integrated in any of the previous groups. These are UCT19Aq, UCT5De, UCT14MgRe and UCT12Co, which morphologically are different. Comparatively these were less erect, more branchingand smaller plant. Their tubers were predominantly oval-elongate to round, with a greater number of principal stems by plant and a clear dominion of flowers of white coloration. The DNA samples were amplified with five primer combinations generating 281 AFLP markers positions, of which 253 (90.04%) were polymorphic (Table 3). The number of polymorphic markers per primer varied from 26 to 71. Primer pair EcoRI-ACC /MseI-CAT contributed the highest number of profiles obtaining 100% of polymorphic bands. These results concurred with those obtained by Savelkoul et al. (1999), who indicated that analysis using AFLP markers ensured a high level of polymorphic bands.
Furthermore, these results agree with those reported by Kim et al. (1998), who obtained a total of 84 polymorphic bands using a single primer combination among 12 potato cultivars. McGregor et al. (2002) indicate that AFLP is a very informative technique for difference potato germplasm, since they obtained 130 polymorphic DNA bands from two pair of combinations. Different combinations of primers achieved different degree of efficiency in the detection of polymorphism (Table 3). It was observed that the combination EcoRI-AGG/MseI-CTT displayed the lowest efficiency in the detection of polymorphisms with just 26 polymorphic bands (76.47%). The most efficient combination was EcoRI-ACC/MseI-CAT, which detected a total of 66 polymorphic bands (100%). Rouppe van der Voort et al. (1998), report that only one combination revealed sufficient information in an analysis of five genotypes of potato. These authors add that the number of monomorphic AFLP markers increases with the number of combinations of primers evaluated and the number of genotypes analyzed. Nevertheless, using different restriction enzymes, different pre-selective and selective nucleotide combinations increase the probability of finding useful polymorphisms; however, a greater number of selective bases lower the detection of polymorphisms. Our results indicated that all the matches analyzed differ in their degrees of similarity. The accessions "UCT5De" and "UCT10MgL" presented the highest genetic similarity (0.89), likewise the accessions "UCT9MgM" and "UCT13MpB" (Table 4). With respect to the accessions from Chiloe, the one, which displayed the least similarity to the rest, was "UCT1Ma". On the other hand, when comparing all accessions included in the study, it appears that they presented on average high genetic diversity (0,73) with similarities ranging between 0,32 and 0,89 (Table 4).
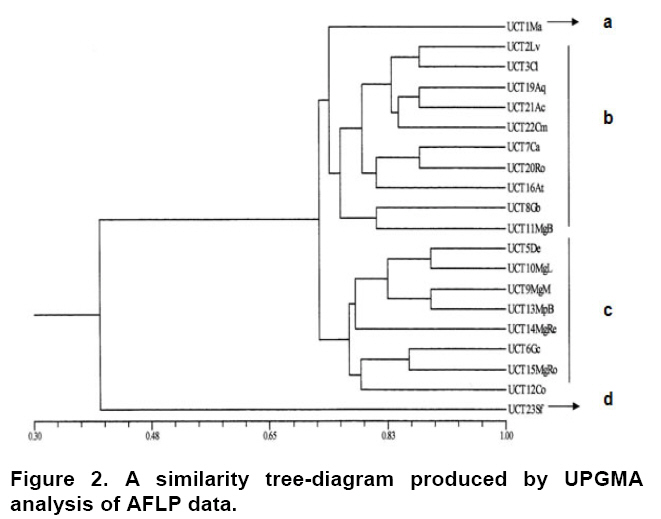
The similarity tree-diagram (Figure 2) derived from the register of all AFLP bands was generated by the use of combinations ACC/CAT, AGG/CTT, ACC/CAT, AAG/CAC and ACA/CTG. Nevertheless a single combination of primers (ACC/CAT) was sufficient to establish a level of optimal polymorphism. Four clusters were defined from the cluster analysis. The first cluster included only the accession UCT1Ma (a) (common name “michuñe azul”), which has a strongly restricted fusiform tuber of an intense blue colour. The second group (b) corresponded to tuber potatoes which have a generally oval form and a light-coloured skin, some with the presence of secondary colours. The flowers of this group are blue or pale-blue in colour. The third group (c) was made up of varieties which are long and smooth in shape, with strong pigmentation. This group presented flowers with strong colours, predominantly purple and intense red. Finally, the last group included the control diploid species, Solanum fernandezianum (d), which confirms the fact that this species belongs to the Etuberosa series. Comparison between morphological and AFLP data The Mantel test showed quite low correlations between the morphological and the molecular (AFLP) dendograms obtained (r = -0,09). Both the morphological and genetic analysis allowed to separate defines groups of native potato germplasm. No significant concordance between AFLP and morphology cluster analyses was observed. In the morphologic analysis, the commercial variety Desiree appeared like an independent variety whereas in the AFLP analysis it integrated group c. The morphological traits of this group are not present in variety Desiree; however, AFLP data indicating that exists a high similarity with the native material of potato of the Chilote group. This result agrees with the hypothesis of the existence of ancestral genes in improvement cultivars of potato. This absence of concordance also has been observed by Xu et al. (2000), Federici et al. (2001), Roldán-Ruiz et al. (2001), Martínez et al. (2003), in varieties of grapevines, weedy rice, ryegrass and azuki beans respectively. Finally, Spooner et al. (2005b), report that techniques of DNA fingerprinting are better than morphological data for discrimination of related genotypes and in the analysis of the genetic similarity. Our results confirm that DNA analysis by AFLP is an efficient method for the exploration of genetic diversity in potato populations. According to our background this is the first study using AFLP markers in Chile to assess the great variability that existing among Chilean germplasm of potato.
Note: Electronic Journal of Biotechnology is not responsible
if on-line references cited on manuscripts are not available any more after
the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07042f2.jpg] [ej07042f1.jpg] |
| |||||||||

{kind=link}
{kind=link}