 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 10, No. 4, October 15, 2007, pg. 604-617 New strains obtained after UV treatment and protoplast fusion of native Trichoderma harzianum: their biocontrol activity on Pyrenochaeta lycopersici Ximena A. Besoain*1,Luz María Pérez2 , Antonio Araya3 , Llacolén Lefever4, Melina Sanguinetti5 , Jaime R. Montealegre6 1Laboratorio
de
Fitopatología
Facultad
de
Agronomía
Pontificia
Universidad
Católica
de
Valparaíso
Quillota,
Chile
Tel:
56
32
274522
Fax:
56
32
274570
E-mail: xbesoain@ucv.cl Financial support: This research was funded by FONDECYT 1040531/04 grant. Code Number: ej07059 The obtainment of 30 new strains from native Trichoderma harzianum after UV light irradiation (UV-A and UV-C), and of 82 strains resulted from protoplast fusion were accomplished. The new strains, initially selected for their growing rate under low temperature and high pH conditions, as well as for their innocuousness on tomato plants, were tested for in vitro inhibition of Pyrenochaeta lycopersici in dual cultures and due to secretion of volatile and diffusible metabolites. All the UV-A and UV-C selected candidate mutants were innocuous to tomato plants, but none of them showed improvement in their biocontrol activity on P. lycopersici. Th12A20.1 increased 1.3 and 1.9 fold the total fresh weight of Fortaleza tomato plants when compared to its parental strains Th12 and Th11, respectively. The selected candidate mutants obtained through protoplast fusion were also innocuous to tomato plants, but only ThF1-2 and ThF4-4 inhibited 1.3 fold (in dual cultures) and 5 fold (due to secretion of volatile metabolites) the growth of P. lycopersici, respectively, in relation to the mean inhibitory effect of both parents. Therefore, these candidate mutants could be included in experiments under field conditions. Keywords: corky root, mutagenesis, tomato diseases, Trichoderma, volatile and diffusible metabolites.
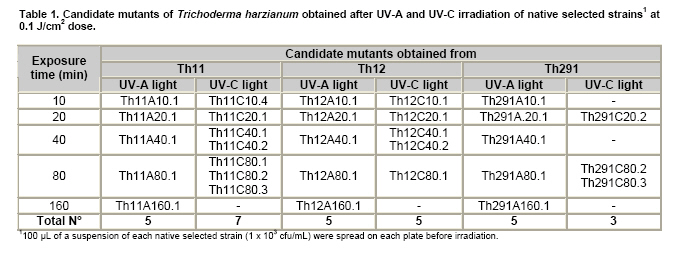
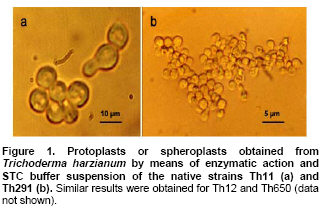
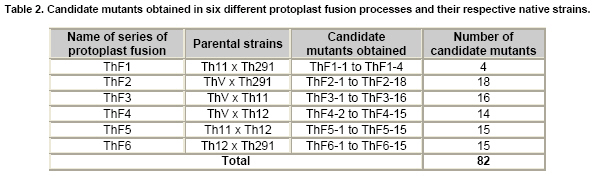
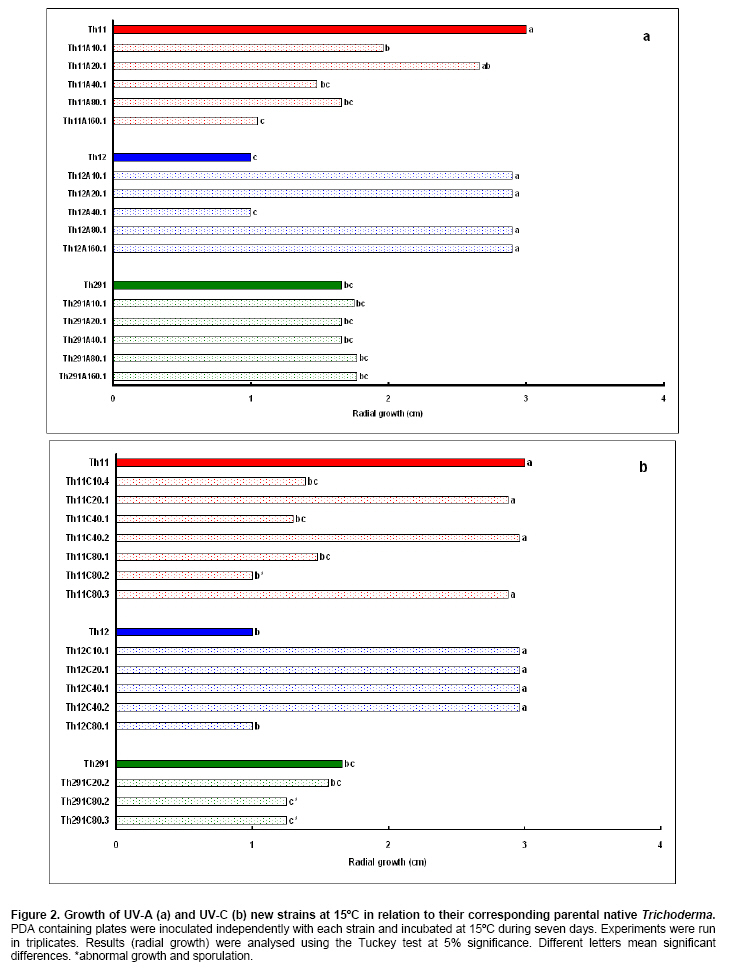
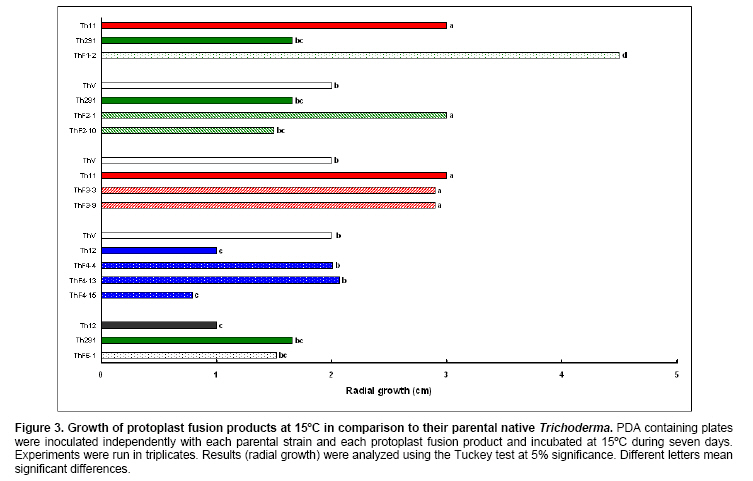
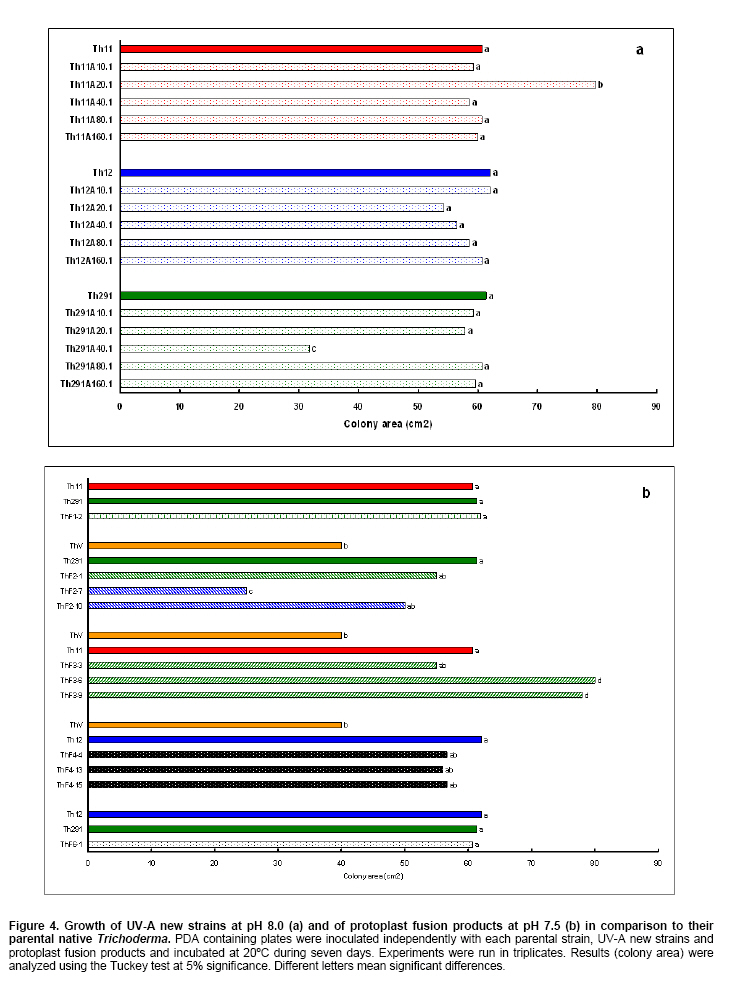
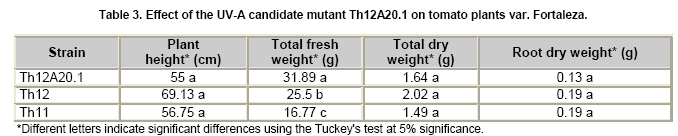
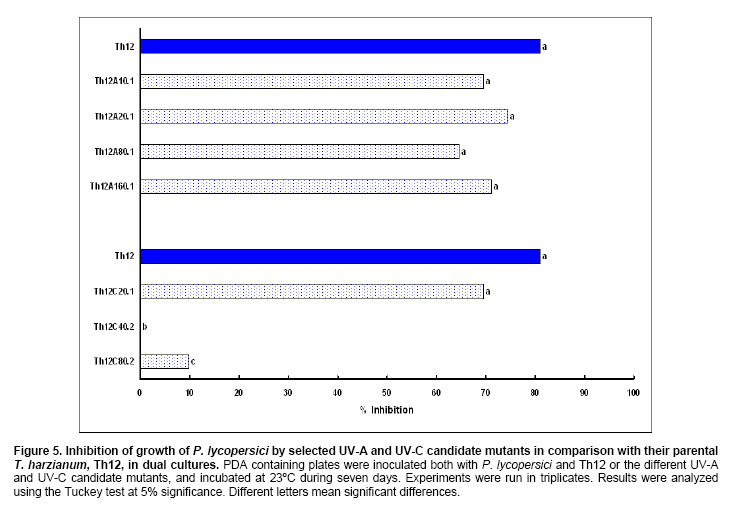
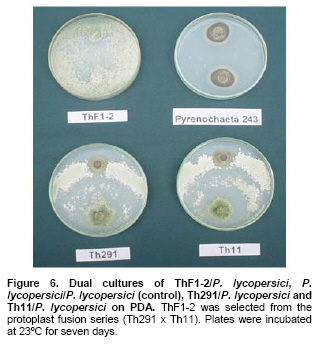
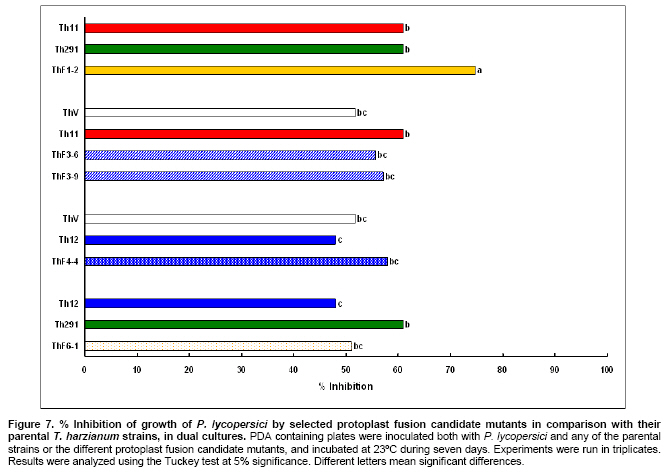
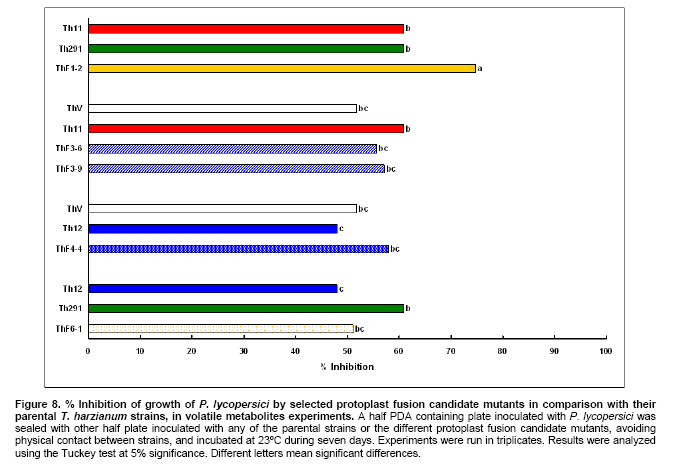
Tomato crops (Lycopersicon esculentum Mill.) have turned to be in Chile and worldwide, the economically most important horticultural product. When grown under unheated greenhouse and in monoculture conditions, a severe disease pressure occurs. Among these, the corky root caused by P. lycopersici Schneider & Gerlach is observed. This fungus is one of the most important pathogens of tomato plants and other vegetable crops both in protected and field crops all over the world (Fiume and Fiume, 2003). The disease results in rotting of tomato fine roots, browning and suberization of main roots and, in extreme conditions, canker at the crown level of the plant. Tomato diseases are normally controlled by means of selected fungicides and soil fumigation with methyl bromide (MeBr). Although the use of MeBr-chloropicrin eradicates many of the fungi involved in root rot complex, it has been shown that reinvasion by fast growing fungi may lessen the effectiveness of fumigation in the later growing season (Pinkerton et al. 2002). The control of P. lycopersici in Chile is accomplished through fumigation with this chemical, which contaminates the environment, affects the ozone layer, destroys the soil microflora, and must be applied every season because of its null residual activity and the rapid re-colonization of soils by the phytopathogens (Ristaino and Thomas, 1997; Pinkerton et al. 2002). Several disease management practices have been proposed to replace MeBr (Ristaino and Thomas, 1997; Fiume and Fiume, 2003). The natural control of several phytopathogens is based on the presence of suppressive soils where different biocontrol microorganisms are detected, such as those belonging to Trichoderma, Gliocladium, Pseudomonas and Bacillus genera, among others (Weller et al. 2002; Montealegre et al. 2003; Guo et al. 2004; Huang et al. 2005; Montealegre et al. 2005). Considering the difficulty to generate suppressive soils similar to the natural ones, the use of selected biocontrol microorganisms could provide an alternative to the use of chemical fungicides. Trichoderma spp. has proved to be useful in the control of phytopathogens affecting different crops (Benítez et al. 2004). Specifically, the use of Trichoderma harzianum Rifai native strains as biocontrol agents (BCAs) in tomato crops could be a useful alternative in the control of soilborne diseases, to be applied under monoculture conditions. Nevertheless, it seems necessary to improve the already positive biocontrol effect of T. harzianum (Pérez et al. 2002; Besoain et al. 2003), especially for its use on tomato crops developed under conditions where P. lycopersici easily produces the corky root disease. In addition, the high alkaline pH found in soils where this crop is cultivated in the central zone of Chile along with low temperatures, should be considered as challenging conditions in the obtainment of improved T. harzianum strains. Taking into account that previous results obtained after testing more than 35 native isolates from different Trichoderma species (T. harzianum, T. koningii, T. piluliferum and T. polysporum) on the development of P. lycopersici in conditions that favored the phytopathogens growth, allowed us to select isolates Th11, Th12, Th291 and ThV (Besoain et al. 2003). Therefore, we decided to improve the biocontrol activity of these selected Trichoderma strains using already described procedures for the obtainment of mutants. Thus, this paper describes the use of UV-A and UV-C radiation and protoplast fusion of native T. harzianum strains that have proved to behave as good BCAs, (Pérez et al. 2002; Besoain et al. 2003), for the obtainment of improved strains; their selection under low temperature and alkaline pH conditions, and the biocontrol effect of the selected new strains on P. lycopersici, under laboratory and glasshouse conditions. UV-A and UV-C irradiation. Th11, Th12 and Th291, native T. harzianum strains, previously selected from suppressive soils where tomatoes were grown for more than 15 years without the use of MeBr, that show a good behavior as BCAs of P. lycopersici (Pérez et al. 2002; Besoain et al. 2003), were used. This isolates were kept lyophilized and stored at -20ºC, for a five-year period. These were recovered in Potato Dextrose Agar (PDA) plates, and conidia were obtained as described (Pérez et al. 2002). Later, 100 µl of suspensions of each strain, containing 1 x 103 cfu/mL, were spread on plates containing corky root medium (CRM) (Shishkoff, 1992), and irradiated with 0.01-0.1 J/cm2 doses of UV-C light (254 nm) or with UV-A light (320 nm), during different time periods (10, 20, 40, 80 and 160 min). After irradiation, plates were incubated at 23ºC for 7-10 days. Grown colonies were purified, lyophilized and stored at -20ºC for further tests. Protoplast fusion. New strains from Th11, Th12, ThV and Th291 native T. harzianum were obtained, using a combination of the Bae and Knudsen (2000), the Stasz et al. (1988) and the Sivan et al. (1990) techniques. Briefly, 3 x 105 conidia of each native strain were seeded in flasks containing 100 mL of sterile yeast extract broth. Th11, Th12, ThV and Th291 were combined in order to obtain six different series: ThF1 (Th11 x Th291), ThF2 (ThV x Th291), ThF3 (ThV x Th11), ThF4 (ThV x Th12), ThF5 (Th11 x Th12), and ThF6 (Th12 x Th291). Flasks were incubated for 23 hrs at 28ºC in constant agitation. The mycelium obtained was collected and incorporated to 10 mL of a cell wall lytic enzymatic solution (L3768, Sigma), and incubated again with agitation during four hrs at the same temperature. Finally, the suspension was filtered through four layers of cheesecloth and centrifuged at 100 x g during 10 min to collect protoplasts or spheroplasts. These were washed in STC buffer (1.2 M sorbitol, 10 mM Tris-HCl pH 7.0, 10 mM CaCl2, Autoclave) to remove the enzyme solution, centrifuged again at the same speed and time, suspended in 1 mL of STC buffer. Protoplasts or spheroplasts were counted in a Bauer hemacytometer. 200 µl of a solution containing 60% (w/v) polyethyleneglycol (PEG), 10 mM CaCl2 and 10 mM Tris-HCl pH 7.5, were gradually added to the protoplast suspension (1 x 105 protoplasts/mL of STC), incubated for a few min and then, a second and a third aliquot of 500 µL of the same PEG solution were added. The mixture was incubated for 10 min at 30ºC, with agitation, to induce fusion. An extra volume of STC (2.2 mL) was added before centrifuging at 100 x g. Protoplasts were diluted in STC to obtain serial dilutions. Aliquots of 100 µL were spread on plates containing protoplast regeneration medium (PRM), (Stasz et al. 1988). Plates were incubated at 23ºC during a week for the obtainment and purification of strains. The preliminary selection of new strains was done on the basis of their growth at low temperatures and high pH, and on the innocuousness on tomato plant growth. The additional selection took into account % inhibition of P. lycopersici growth in dual cultures and due to volatile and/or diffusible metabolites production. Growth at different temperatures and pH. The selection of new strains obtained both after UV light irradiation or protoplast fusion, as well as the behavior of their corresponding native isolates, was carried out by seeding a mycelium plug on PDA plates and incubated at 10ºC and 15ºC for seven days. Growth at different pH values was estimated using PDA plates adjusted to pH 6.0, 6.5, 7.0, 7.5 and 8.0, after autoclaving, with KOH or HCl. Plates were incubated at 20ºC for seven days. Each experiment was run in triplicates. Results were analyzed using the Tuckey test at 5% significance. Innocuousness tests of native and new strains on tomato plants. (a) New strains obtained after UV irradiation. A sterile distilled water suspension (5 x 105 conidia/mL) of each native or new strain of T. harzianum obtained after UV irradiation was prepared, to be used as inocula for the evaluation of possible damages or benefits of these strains on tomato plants. Tests were run as follows: tomato fine roots were submerged in the conidia suspension, and once inoculated, seedlings were transplanted to pots containing a mixture of compost:sand:soil = 3:1:1 (v/v/v), and kept under controlled greenhouse conditions (controlled range of temperature: 12ºC (minimum) and 20ºC (maximum); 12 hrs natural light, irrigation: once a week). One month after transplantation, they were pulled out for evaluation. Each unit of treatment considered five plants of each of the four cultivars used (Fortaleza, R593, Naomi and Yonit), per each of the strains tested. Pots were arranged in a complete random design. Once the plants were removed, the potting mix was rinsed from the roots and the total fresh weight was measured. Then, plants were dried in paper bags for 48 hrs at 60ºC to obtain the total dry weight. Treatments considered a single new strain. Controls were run with the parental strains. Results correspond to mean and were analyzed by the Tuckey's test at 5% significance. (b) New strains obtained through protoplast fusion. Pellets that contained the different strains of Trichoderma were prepared according to Montealegre and Larenas (1997) and used as inoculums for the evaluation of possible damages or benefits on tomato plants varieties Yonit and Dalila (Zeraim Gedera) cultivated in a cold greenhouse in Quillota, V Region, Chile. Nine plants were used in each treatment, with a completely randomized design. Treatments considered the use of native isolates, Th291, Th11, Th12 and Th650, the new strains obtained through protoplast fusion F1-2, F4-4 and F5-8 the UV-C irradiated new strain Th11C80.3, and a commercial product based on T. harzianum (Orious, powder formulation). Controls were run without the addition of BCAs. Seedlings were transplanted into plastic bags containing natural infested clay soil with P. lycopersici, obtained from a monoculture tomato crop in the Quillota area. Recently prepared pellets containing new and native strains, treatments were used in 1-3 g doses, depending on the concentration of each BCA, previously determined by viability tests. Pellets were set in the planting hole before transplanting. Two months later after transplanting, the plants were pulled out for evaluation. Once the plants were removed, the potting mix was rinsed from the roots and the total fresh weight was measured. Then, plants were dried in paper bags for 48 hrs at 60ºC to obtain the total dry weight. Treatments considered a single new strain. Controls were run with the parental strains. Results correspond to mean and were analyzed by the Tuckey's test at 5% significance. Dual cultures. The native isolates of T. harzianum, as well as the new strains obtained either after UV light irradiation or protoplast fusion, were spread on acidified PDA plates (1mL of 1M lactic acid per L of PDA medium), placing two equidistant 0.5 cm diameter agar disks, one containing young mycelium of BCA (Tewari and Bhanu, 2004) and the other, P. lycopersici mycelium. Controls were run placing two equidistant 0.5 cm agar disks of the BCA or the phytopathogen. Plates were incubated for 7 days at 23ºC. The area of the pathogen colony was measured and compared to the control. Each experiment was run in triplicates. The pathogen growth inhibition was calculated as follows: Pathogen growth inhibition (%) = 100 - % Control Control (%) = ACP/ACC x 100; (ACP: colony area of pathogen with biocontroler; ACC: colony area of pathogen with same pathogen) Production of diffusible metabolites. PDA containing plates were inoculated with a 5 mm disk of each of the native and the new strains of Trichoderma. The inoculum was placed on a cellophane paper disk (non water proof, 0.6 µm pore size as in Kosuta et al. (2003)) which covered completely the plate, as described by Whipps (1987). After incubation at 15ºC and before the mycelial growth reached the edges of the cellophane paper, it was removed along with the grown Trichoderma. The plate was inoculated again at the centre, but this time with a 5 mm disk of pure mycelia of P. lycopersici. The plates were further incubated at 15ºC for seven days and the pathogen growth was measured. Mocked inoculated plates with a 5 mm disk of PDA and inoculated with P. lycopersici on the cellophane paper as above, were used as controls. The experiment, considering a single Trichoderma strain, was run in triplicates. Results were analyzed using the Tuckey test at 5% significance. Production of volatile metabolites. In order to evaluate the presence of volatile metabolite in mutants or native isolates of T. harzianum, the methodology described by Whipps (1987) was used. Two PDA containing Petri dishes (half plates of the same diameter) were set against each other after inoculation, and were sealed with waterproof adhesive tape, to avoid leakage of metabolites. Inoculation was done as follows: one half dish was inoculated with a 0.5 cm diameter disk of young mycelium of the BCA, and the other half dish was inoculated with P. lycopersici, this last half dish was placed on top. Due to the slow growth of P. lycopersici, it was seeded three days before Trichoderma strains.The plates were incubated at 15ºC for four days and the growth of the pathogen was measured and compared to controls run using the same inoculum in both half dishes. The experiment, considering a single Trichoderma strain, was run in triplicate. Results were analyzed using the Tuckey test at 5% significance. A total of thirty colonies were developed after irradiation with UV-A and UV-C, plates containing the native isolates of T. harzianum Th11, Th12 and Th291 (Table 1). Fifteen of them grew after 10 to 160 min of UV-A irradiation, resulting in three new strains from each parental one. A 1% recovery for each time of irradiation was calculated from results. The fifteen new strains recovered after irradiation of Th11, Th12 and Th291 with UV-C, which grew after 10 to 80 min irradiation, ranged from 0.67% to 2%, depending on the UV-C exposure time and on the parental strain. No strain development was observed after 160 min exposure to UV-C of any of the parental strains. Seven, five and three new strains were recovered after UV-C treatment of Th11, Th12 and Th291, respectively (Table 1). The enzymatic treatment of native strains of T. harzianum allowed the obtainment of protoplasts or spheroplasts, such as those shown in Figure 1. A total of 82 new strains (candidate mutants) were obtained after the six protoplast fusion series (Table 2), which corresponds to a mean of 0.014% total yield. Growth at different temperatures and pH. All the new strains (Table 1) obtained after UV-A and UV-C irradiation of Th11, Th12 and Th291 grew the same as their parental strains at 10ºC. On the other hand, growth at 15ºC differed from the observed at 10ºC: while the UV-A and UV-C derived new strains Th12A10.1, Th12A20.1, Th12A80.1, Th12A160.1 and the five Th12 UV-C derived new strains grew significantly better than the corresponding parental ones; Th11A20.1, Th11C20.1, Th11C40.2, Th11C80.3, Th12A40.1, and the Th291A10.1, Th291A20.1, Th291C20.2, Th291A40.1, Th291A80.1, Th291A160.1 showed no significant differences in growth with the parental Trichoderma, after analysis of results by the Tuckey test at 5% significance (Figure 2). The remainder UV-A and UV-C new strains grew worse than their corresponding parental strains. The UV-C new strains Th11C80.2, Th291C80.2 and Th291C80.3 were discarded for further experiments because of their abnormal growth and sporulation at the temperatures already mentioned. On the other hand, the following 11 mutant candidates of the 82 obtained through protoplast fusion (Table 2), grew at 10ºC: ThF1-2, ThF2-1, ThF2-7, ThF2-10, ThF3-3, ThF3-6, ThF3-9, ThF4-4, ThF4-13, ThF4-15 and ThF6-1. From these, ThF4-4, ThF4-13 and ThF4-15 did not show significant differences in their growth at 10ºC when compared to their parental strains. In addition, ThF1-2, ThF2-7 and ThF6-1 grew better, and the remainder candidate mutants grew worse than their two parental strains. Analysis of behavior at 15ºC (Figure 3) showed that the same protoplast fusion products mentioned above were able to grow at this temperature, although significant differences could be observed for some of them: ThF1-2 and ThF2-1 grew better, ThF4-15 grew worse and the remainder protoplast fusion products grew the same when compared to their parental Trichoderma after analysis of results by the Tuckey test at 5% significance. No differences were observed in the growth of UV-C new strains at pH 7.5 and 8.0 when compared to their parental Trichoderma. The UV-A new strains Th11A10.1, Th11A80.1, Th11A160.1 and Th291A10.1 grew significantly worse at pH 7.5 when compared to Th11 or Th291, while the growth of all those derived from Th12 did not show significant differences with the parental strain. Growth at pH 8.0 showed that all UV-A new strains showed no significant differences with their parental Trichoderma, except Th11A20.1 that grew significantly better than Th11, and Th291A40.1, that grew worse than Th291 (Figure 4). The protoplast fusion products showed significant differences at pH 7.5 and 8.0 when compared to both parental Trichoderma. Growth experiments run at pH 7.5 showed that ThF3-6 and ThF3-9 grew significantly better than both parental isolates; ThF3-3, ThF4-4, ThF4-13 and ThF4-15 grew significantly better than one of their parental Trichoderma; ThF1-2, ThF2-1, ThF2-10 and ThF6-1 showed no significant differences and only ThF2-7 grew worse than any of their parental strains (Figure 4). Growth at pH 8.0 showed that the protoplast fusion products derived from Th11 x Th291 (ThF1) or from ThV x Th291 (ThF2) grew worse than any of their parental Trichoderma, while the remainder ThF3, ThF4 and ThF6 (Table 2) showed no significant differences with their parental strains. Innocuousness tests of native and candidate mutants on tomato plants. Pot and greenhouse trials showed that all the selected UV-A and UV-C candidate mutants were innocuous towards the tomato plants varieties tested: Fortaleza, R593, Naomi and Yonit. In addition, Th12A20.1 significantly increased the total fresh weight of Fortaleza tomato plants in relation to the native isolates Th11 and Th12 (Table 3). On the other hand, the selected protoplast fusion candidate mutants were also innocuous towards the different tomato plants varieties tested: Yonit and Dalila. None of the isolates produced a detrimental effect in contrast to control plants, grown in the absence of any BCAs. Since the adaptation to low temperature (10ºC-15ºC) and high pH (7.5-8.0) conditions are important factors in the biological control of the tomato corky root disease, the UV-A, the UV-C and the protoplast fusion candidate mutants that grew the same in one condition (Tº or pH) and the same or better in the other condition (Tº or pH) than their corresponding parental Trichoderma, and were innocuous to tomato plants, were selected for further experiments. Therefore, the selected UV-A candidate mutants were Th12A10.1, Th12A20.1, Th12A40.1, Th12A80.1 and Th12A160.1; the selected UV-C candidate mutants were Th12C20.1, Th12C40.2 and Th12C80.1, and the selected protoplast fusion candidate mutants were ThF1-2, ThF3-6, ThF3-9, ThF4-4 and ThF6-1. Inhibition of P. lycopersici growth in dual cultures. The % inhibition of P. lycopersici growth by UV-A selected candidate mutants showed no significant differences when compared to the 82% inhibition caused by Th12. A similar behavior was observed with the UV-C selected candidate mutant Th12C20.1, while Th12C40.2 and Th12C80.1 did not inhibit and poorly inhibited (10%) the growth of the phytopathogen, respectively (Figure 5). The ThF1-2 candidate mutant inhibited the growth of P. lycopersici (Figure 6) significantly better than any of its parental strains, being 1.3 fold more effective than Th11 or ThV (Figure 7). The other selected candidate mutants from the protoplast fusion products (series ThF3, ThF4 and ThF6) did not show significant differences in the % inhibition of P. lycopersici growth. Inhibition of P. lycopersici growth due to the production of diffusible metabolites. Analysis of the effect of diffusible metabolites secreted by the different UV-A, UV-C and protoplast fusion selected mutant candidates showed no significant differences in % inhibition of P. lycopersici growth (Data not shown). Inhibition of P. lycopersici growth due to the production of volatile metabolites. The % inhibition of P. lycopersici growth by UV-A or UV-C selected candidate mutants showed no significant differences when compared to the 9.0% inhibition caused by volatile metabolites secreted by Th12 (Data not shown). On the opposite, significant differences were observed for the ThF4-4 candidate mutant that showed 51% inhibition of P. lycopersici growth, effect which is much higher than the observed for the rest of the selected candidate mutants and native strains (Figure 7). The obtainment of the same number of new strains of T. harzianum from Th11, Th12 and Th291 after irradiation with UV-A (Table 1), showed that this treatment allowed the survival of a low percent of strains (a total of 1% from the three parental T. harzianum), where possible mutation(s) did not alter their ability to develop in the same culture media of their parental T. harzianum, but also showed no differences in the sensitivity of the parental strains to UV-A light. These results are consistent with previous reports on the sensitivity of T. harzianum to UV irradiation (Paul et al. 2005), and also suggest that early damage of most of the T. harzianum conidia population occurred because percent recovery did not change after the different time periods of irradiation. Other research groups have been successful in obtaining Trichoderma mutants by means of UV-mutagenesis (Szekeres et al. 2004; Hatvani et al. 2006); nevertheless, those UV mutants were obtained using only UV-C irradiation. Our results showed that UV-C affected differently each parental strain of T. harzianum (Th11, Th12 and Th291), because different number of new strains was recovered from each parental T. harzianum after treatment, being the new strains obtained from Th11 the most abundant. In addition, an opposite to UV-A irradiation, long exposure time to UV-C (160 min, Table 1) resulted in no recovery of strains from any of the parental ones, a fact that suggest that long time exposure to this wavelength produced a lethal effect on the surviving conidia at lower time exposure periods. Papavizas and Lewis (1983) evaluated the effect of irradiating V-8 agar plates containing 1 x 105 conidia under UV-C for 80 min. In those conditions, these authors obtained biotypes (R3 and R4) that showed better antibiotic secretion in comparison to the original isolate T-1, suggesting that it is possible to induce new stable biotypes of Trichoderma spp. with long exposure time to UV-C irradiation. Other authors used different experimental conditions for the obtainment of UV-C mutants from those used by Papavizas and Lewis (1983), or the used in this work: UV mutants of T. harzianum T334 were obtained after irradiation for 25-30 sec with a UV-C lamp at 23 cm distance, with 10% yield after selection using p-fluorophenylalanine (Szekeres et al. 2004), or UV mutants of T. harzianum T66 and of T. atroviride T122 were obtained after irradiation for 30 sec with a UV-C lamp at 40 cm distance. This latter work included 50 µg/mL tecobunazole for selection of mutant, but no yield data was provided (Hatvani et al. 2006). Our UV-C new strains developed and sporulated normally at 10ºC and 15ºC (Figure 2b), after 80 min irradiation suggesting a higher resistance to UV-C radiation than the strains used by Szekers et al. (2004) or Hatvani et al. (2006), excepting for Th11C80.2, Th291C80.2 and Th291C80.3 where abnormal growth and sporulation was observed. For these reasons, they were discarded for further tests. It is probable that mutations altering growth and sporulation were produced, being these parameters important for biocontrol activity. A mean yield of 14 x 10-3% candidate mutants was obtained after protoplast fusion (six different series; Figure 1 and Table 2), methodology previously used by Stasz et al. (1988) and by Hatvani et al. (2006). Our yield was much higher than the obtained by Hatvani et al. (2006) for mutants resistant to 12.5 µg/mL of tebuconazole and 1.5 µg/mL MBC, where yield was of 0.75 x 10-4% for T66 mutants and of 1.25 x 10-4% for T122 mutants. Nevertheless, it is important to mention, that the number of our new mutants was obtained without any further growth restriction. When temperature conditions were used for selection, our yield decreased to 1.9 x 10-3%, result that is still higher to the one obtained by Hatvani et al. (2006) at lower concentrations of the fungicides (0.15 x 10-3% for T66 and 0.25 x 10-3% for T122). Several UV-A, UV-C and protoplast fusion mutant biotypes (Figure 2, Figure 3 and Figure 4) behaved better or the same at low temperatures or at alkaline pH when compared to their parental strains, approach that was used by other groups to select Trichoderma strains that could control phytopathogens affecting different crops at temperature values of 10ºC (Antal et al. 2000; Kredics et al. 2000; Kredics et al. 2004). Antal et al. (2000) describe that the different Trichoderma strains tested at 10ºC biocontroled Rhizoctonia solani and Fusarium oxysporum f. sp. dianthi, while T. aureoviride and T. viride were more effective against Pythium debaryanum than T. harzianum at this same temperature. Pyrenochaeta, as described by Shishkoff (1992), that causes corky root in the absence of another pathogen, develops at soil temperatures around 14ºC; therefore, the Trichoderma mutants obtained in this work, and selected for their growth and sporulation at 10ºC and 15ºC, may act similar to those described by Antal et al. (2000), in the biocontrol of P. lycopersici in the weather and soil conditions found in Chile. As opposed to this, all isolates of P. lycopersici described in Italy by Infantino et al. (2003) showed an optimal growth temperature of 23ºC; therefore, temperature is a fact that must be taken into account for the isolation and obtainment of new biocontrol strains, because development of breeding programs for resistance against tomato corky root of tomato must considered weather and temperature conditions where BCA could develop in optimal conditions. The development of P. lycopersici is not affected by pH in the range of 4.0 to 8.0, while different native Trichoderma isolates have shown a more vigorous growth at low pH than at high pH and at temperatures between 25ºC and 30ºC (Besoain et al. 2003). This work also describes that soils found in Quillota, V Region, Chile, combines a high pH and low temperature, conditions that are optimal for P. lycopersici attack on off-season grown tomatoes. Therefore, and due to characteristics already mentioned, the biocontrol on P. lycopersici obtained by the native Trichoderma isolates was not the one expected, because they needed temperatures higher than those optimal for P. lycopersici development to grow (Pérez et al. 2002; Besoain et al. 2003). Thus, the obtainment of candidate mutants that grew the same or better both at alkaline pH and low temperatures when compared to the native strains, allowed us to select the UV-A strains Th12A10.1, Th12A20.1, Th12A40.1, Th12A80.1 and Th12A160.1; the UV-C strains Th12C20.1, Th12C40.2 and Th12C80.1, and the selected protoplast fusion candidate mutants ThF1-2, ThF3-6, ThF3-9, ThF4-4 and ThF6-1, to test their innocuousness on tomato plants previous to run inhibitory tests on P. lycopersici development. The innocuousness and non detrimental effect on tomato plants of all the selected UV-A, UV-C and protoplast fusion products was proved, suggesting that any of them may be used in the biocontrol of tomato pathogens. In addition, Th12A20.1 (Table 3) showed a growth promoter activity, as described for other Trichoderma strains (Altomare et al. 1999; Harman, 2000; Howell, 2003; Benítez et al. 2004). ThF1-2 (Figure 6 and Figure 7), the only candidate mutant that produced an 80% inhibition of P. lycopersici growth in dual cultures, most probably acquired genes that improved the biocontrol activity of its parents Th11 and Th291, suggesting that DNA transfer and/or recombination occurred. Hatvani et al. (2006) obtained three fusion products resistant to both tebuconazol and MBC, but these were recovered from UV mutants obtained from the same parental strains, one resistant to MBC and the other to tebuconazol. Early experiments run by Whipps (1987), showed that P. lycopersici development in dual cultures experiments, could be inhibited 33% and 39% by T. harzianum isolates a and b respectively, only when Tap Water Agar (TWA) was used and with no significant results when PDA was the culture media. In contrast, our results were obtained using the latter media obtaining 80% inhibition of the phytopathogen. None of the selected candidate mutants improved the inhibition of P. lycopersici growth due to the secretion of diffusible metabolites, as has been described for other Trichoderma strains (Benítez et al. 2004), suggesting that this biocontrol mechanism is not present in the new strains, or was changed or lost during UV irradiation or protoplast fusion, as has been described for mutants from T. viride (Chet et al. 1997). The five fold inhibitory effect on P. lycopersici due to the secretion of volatiles of ThF4-4, the fusion product of Th12 and ThV (Figure 8), suggests that also in this case, an effective fusion occurred. The behavior of ThF4-4 is significantly different from its parental strains, as opposite to results obtained with all other UV-A, UV-C or protoplast fusion candidate mutants. It is well known that T. harzianum can use different mechanisms such as secretion of volatile and diffusible metabolites, and hydrolytic enzymes among others (Altomare et al. 1999; Harman, 2000; Howell, 2003; Benítez et al. 2004; Montealegre et al. 2005). Th11 and Th12, previously selected strains for their control of P. lycopersici (Pérez et al. 2002; Besoain et al. 2003) did not produce UV-A or UV-C mutants with improved inhibitory activity on P. lycopersici due to volatile metabolite secretion; therefore, this biocontrol mechanism could be absent in the new strains, or was changed or lost during UV irradiation. Finally, we may conclude that the candidate mutants Th12A20.1, ThF1-2 and ThF4-4 should be included in future experiments, being conscious that in vitro or glasshouse experiments run to control phytopathogens does not necessarily correlate with the field performance experiments (Brunner et al. 2005). Thus, it seems important to test these selected BCAs under field conditions before including them in an integrated management program. The authors thank Dr. Eduardo Oyanedel, Department of Horticulture, Facultad de Agronomía, Pontificia Universidad Católica de Valparaíso, for critical revision of this manuscript. The researchers are grateful to Mrs. Gladys Andrade and Mr. Ivan Cortés for their technical assistance.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2007 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej07059f5.jpg] [ej07059f8.jpg] [ej07059f3.jpg] [ej07059f4.jpg] [ej07059t2.jpg] [ej07059f6.jpg] [ej07059t3.jpg] [ej07059f7.jpg] [ej07059f1.jpg] [ej07059t1.jpg] [ej07059f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}