 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 1, January 15, 2008, pg. Overexpression of the pineapple fruit bromelain gene (BAA) in transgenic Chinese cabbage (Brassica rapa) results in enhanced resistance to bacterial soft rot Yu-Jin Jung1 , Chang-Sun Choi2 , Jin-Heui Park3, Hee-Wan Kang4, Je-Eul Choi5, Ill-Sup Nou6, Soon Youl Lee*7, Kwon-Kyoo Kang*8 1 Department of Horticulture,
Hankyong National University,
67 Sukjong-dong, Ansung city,
Kyonggi-do 456-749, South Korea,
Tel: 82 31 670 5107,
Fax: 82 31 670 5333
E-mail: yuyu1216@naver.com Financial support: This work was supported by a grant from the Agricultural R&D Promotion Center (Project No. 204068-3). Received January 26, 2007 / Accepted May 20, 2007 Code Number: ej08007 Abstract Bromelain is a crude protein extract obtained from pineapple stems, which comprises a variety of proteolytic enzymes. It exhibits potential therapeutic activities against trauma, inflammation, autoimmune diseases and malignant disorders. In this study, we cloned BAA1 (the gene encoding fruit bromelain) into a plant expression vector that was then used to transform Brassica rapa and overexpress BAA1 under the control of the cauliflower mosaic virus (CaMV) 35S promoter. We demonstrate that constitutive overexpression of BAA1 in B. rapa confers enhanced resistance to the soft rot pathogen Pectobacterium carotovorum ssp. carotovorum. These results suggest that it could be utilized for protecting plants from attack by bacterial pathogens. Keywords: Brassica rapa, fruit bromelain, soft rot resistance, transgenic plant.
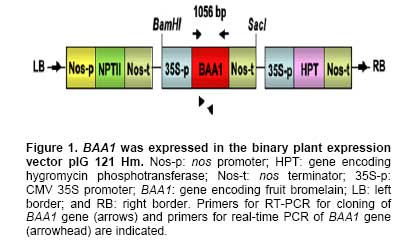
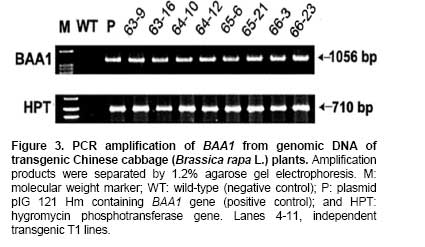
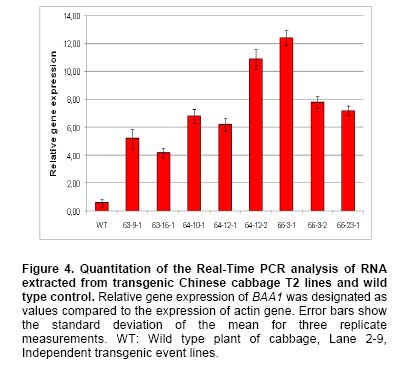
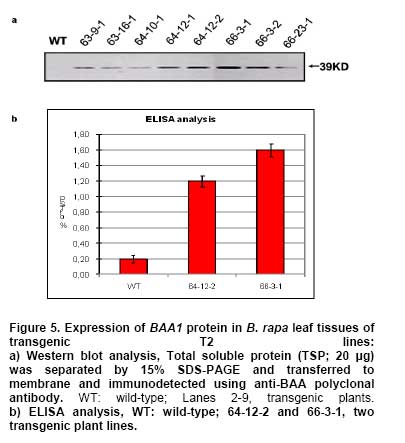
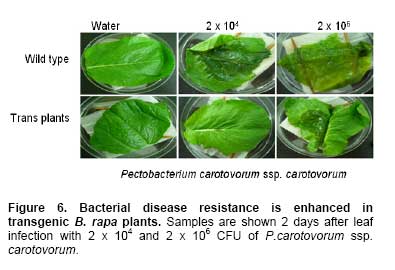
Bacterial soft rot is a common disease caused by Pectobacterium carotovorum ssp. carotovorum, which causes serious damage and economic losses in many vegetable crops including carrot, radish, potato and all types of Brassica. It is considered one of the most destructive diseases of Chinese cabbage (Brassica rapa), which is cultivated throughout China, Japan, and Korea (Kikumoto, 1981; Li, 1981). Plants employ a complex array of defense mechanisms to protect against invading phytopathogenic microorganisms. For example, plants will fortify their cell walls as a physical barrier and synthesize antimicrobial compounds such as phytoalexins (Dixon, 1986), pathogenesis related (PR) proteins (van Loon and van Kammen, 1970; Linthorst, 1991; Yun et al. 1997), thionins, ribosome-inactivating proteins, defensins and nonspecific-lipid transfer proteins (Broekaert et al. 1997). Bacterial soft rot disease is hard to prevent due to the broad host range and latency in crop waste. Since chemical control is not developed yet, traditional cultural practice is still applied to prevent soft rot disease in Chinese cabbage. However, there are some reports showing increased resistance to soft rot by conventional breeding methods. When interspecific hybridization was performed either by sexual cross or somatic cell fusion of Chinese cabbage plus Kale, the progeny showed increased resistance to soft rot (Yoshikawa et al. 1989; Yamagishi et al. 1990). Recently, transgenic plants expressing several genes were reported to show resistance to the soft rot-causing bacterial pathogen. The presence of pflp (ferredoxin-like protein) gene in the transgenic plant showed the increased resistance to bacterial pathogen (Yip et al. 2007). In addition, transgenic potatoes expressing heterogeneously ATP/ADP transporter gene (Linke et al. 2002) and bacterial pheromone N-acyl-homoserine lactone synthesis gene (Toth et al. 2004) showed much improved resistance to soft rot. Bromelain (EC 3.4.22.4) was isolated from the stem of pineapples and was characterized as a complex of cysteine proteases (Taussig and Batkin, 1988; Maurer, 2001). Cysteine proteases are involved in various physiological and developmental processes in plants including programmed cell death during organ senescence and tracheary element differentiation (Beers et al. 2000; Subbaiah et al. 2000; Lam, 2005; Beyene et al. 2006). The precise roles of proteinases in these and other plant programmed cell death processes are under investigation (Beers et al. 2000). Bromelain is reported to have a number of potential therapeutic applications, including treatment of trauma, inflammation, autoimmune diseases, enhancement of immune response, and malignant disorders (Maurer, 2001; Orsini, 2006). When used in conjunction with antibiotic therapy, bromelain has been shown to increase antibiotic effectiveness and absorption (Luerti and Vignali, 1978; Tinozzi and Venegoni, 1978). In addition, bromelain is known to be relatively safe and not to show side effects, toxicity and resistance (Mynott et al. 2002). However, the mechanisms underlying these pharmacological effects remain not understood. Tinozzi and Venegoni, 1978). In addition, bromelain is known to be relatively safe and not to show side effects, toxicity and resistance (Mynott et al. 2002). However, the mechanisms underlying these pharmacological effects remain not understood. The 1056 bp BAA1 gene encoding pineapple bromelain was cloned and the gene product was grouped to be a peptidase C1A subfamily, composed of cysteine peptidases (CPs) similar to papain, including the mammalian CPs (cathepsins) (Muta et al. 1993). Even though several protease and cathepsins were reported to be involved in the host-defense mechanism (Tani et al. 2001; Shafer et al. 2002) and to show antimicrobial activity, the exact physiological role of the BAA1 in the plant defense system is not well characterized yet. In this study, we wanted to know whether bromelain protease has an antimicrobial activity when expressed in plants since the role of bromelain has been reported in a number of potential therapeutic applications and to increase antibiotic effectiveness as described above. We isolated BAA1, which encodes bromelain cysteine protease from pineapple (Ananas comosus L. Merrill) and over-expressed this gene in B. rapa. We then investigated the resistance of the resulting transgenic plants to bacteria using a Pectobacterium carotovorum ssp. carotovorum bioassay. Materials and Methods Transgene construction, preparation of explants, bacterial strain and transformation We isolated BAA1 cDNA from total RNA extracted from pineapple fruit using the primers BAA-F (5’-ATG GCT TCC AAA GTT CAA CTC GTG-3’) and BAA-R (5’-TCA AGT TTC AGA AAC CAT CTT-3’), which were based on the published sequence (GenBank: GI2342495). The BAA1 cDNA was cloned into a transgene cassette under the control of the cauliflower mosaic virus 35S promoter (CaMV 35S promoter), then subcloned into the binary Ti-plasmid vector pIG 121 Hm (Clontech, Palo Alto, CA, U.S.A), which contains both kanamycin and hygromycin resistance genes for bacterial and plant transformation selection, respectively (Figure 1). For sterilization, seeds were submerged in 70% ethanol for 2 min, then in 1.5% NaOCL for 10 min. They were then rinsed eight times with sterilized water and plated onto MS medium (Murashige and Skoog, 1962) and incubated under light conditions for 6 days. Following germination, the hypocotyls were cut into ca. 5-7 mm segments and pre-cultured for 2 days on MS solid medium containing 1 mg/L 6-Benzyladenine (6-BA) and 1 mg/L α-naphthaleneacetic acid (NAA). Explants were then used for transformation. Glycerol stocks of Agrobacterium tumefaciens LBA4404 were used to inoculate liquid YEB medium (pH 7.2) containing 5 g/L tryptone, 5 g/L yeast extract, 5 g/L sucrose, 2 mM MgSO4 and 50 mg/L kanamycin, then incubated overnight at 27-28ºC with constant shaking (200 rpm). At mid-log phase of OD600 value of 0.9, 0.1% of the culture was transferred to fresh medium and cultivated to OD600 = 0.4. Cells were collected by centrifugation and re-suspended in sterilized water. Explants were immersed in the bacterial suspension for 2-3 min with constant shaking. The infected hypocotyls were then blotted onto filter paper and then transferred on the MS plate without any antibiotics and was incubated for 2 days at 25ºC under dark. It was then transferred on callus induction medium (CIM; B5 salts, 1 mg/L 2,4-D, 3% (w/v) sucrose, 0.8% (w/v) agar, 500 mg/L carbenicillin) and incubated for 14 days at 25ºC under 16 hrs light / 8 hrs dark photoperiod. Finally, it was moved onto differentiation medium (DM; B5 salts, 3 mg/L BAP, 1 mg/L zeatin, 3% (w/v) sucrose, 0.8% (w/v) agar, 50 mg/L hygromycin, 500 mg/L carbenicillin) and grown for 14 days. It was subcultured 3 more times every 14 days. Differentiated shoots were moved to maturing medium (MM; B5 salts, phytohormone free, 3% (w/v) sucrose, 0.8% (w/v) agar, 50 mg/L hygromycin, 500 mg/L carbenicillin) for 3 weeks. Roots were inducted on MS medium with 1 mg/L IBA (Indole-3-butyric acid) and 50 mg/L hygromycin. Eventually transformed seedlings were planted in pots for further growth. All the media above contained 3% (w/v) sucrose (pH 5.8) and all explants were cultured under conditions of 16 hrs light (2000 Lux) / 8 hrs dark, at 23-25ºC. Total DNA was extracted from various independent transgenic lines using a DNeasy Plant Kit (Qiagen, Germantown, MD, U.S.A.). Independent BAA- and HPT-transgenic ‘T0’ and ‘T1’ lines were screened by polymerase chain reaction (PCR) using genomic DNA of the transgenic plants as template DNA and BAA1 gene-specific primers and hygromycin-resistance gene specific primers. The reaction conditions for the PCR were: 1 cycle of 94ºC for 1 min and 30 cycles of 94ºC, 30 sec; 55ºC, 30 sec; and 72ºC, 1 min and 1 cycle of 72ºC for 10 min. PCR amplifications were performed in a Minicycler (MJ Research, Waltham, MA, U.S.A.) using the following primers, HPT specific primers (5’-GCG TGA CCT ATT GCA TCT CC-3’ and 5’-TTC TAC ACA GCC ATC GGT CC-3’) and BAA1-gene specific primers used in the cloning of the gene. Total RNA was isolated from Chinese cabbage using the TRIzol method (Invitrogen, Carlsbad, CA, U.S.A.). In all cases, RNA was treated with RNase-free DNase and the DNase removed according to the manufacturer's instructions (AMBION, Austin, TX, U.S.A.). RNA was quantified in a spectrophotometer at 260 nm. Total RNA (5 µg) was reverse transcribed in 20 µl reactions using random hexamers (GE Healthcare Bio-Sciences Corp., Piscataway, NJ, U.S.A.), RNaseH-free reverse transcriptase (Fermentas Life Sciences, Burlington, ON, Canada) and GeneAmplimer pAW 109 RNA (2.5 x 105 copies; Applied Biosystems, Foster City, CA, U.S.A.). The reaction was diluted 50-fold and 5 µl added to a 25 µl PCR amplification reaction containing the BAA gene-specific primers BAA-F and BAA-R, as described above. Amplification of the constitutively-expressed gene encoding actin was used as a loading control (actin primers: 5’-ATG GTT GGG ATG GGT CAA AAA-3’ and 5’-ACG GAG CTC GTT GTA GAA AGT-3’). Amplification of GeneAmplimer pAW 109 RNA (primers: 5’-CAT GTC AAA TTT CAC TGC CTT CAT C-3’ and 5’-TGA CCA CCC AGC CAT CCT T-3’) was used as a positive control for the RT-PCR (reverse transcription PCR) reaction efficiency. PCR reactions were separated by agarose gel electrophoresis and stained with ethidium bromide. In order to obtain semi-quantitative results, the number of cycles used for PCR amplification of each gene was adjusted so as to obtain barely-visible bands on agarose gels. The expression of BAA1 gene was quantitated using actin gene as a reference gene. The cDNA was diluted 100-fold for real-time quantitative PCR experiments (RT-qPCR) using SmartCycler II (Takara, Shiga, Japan) and SYBR RT-PCR kit (Perfect Real Time, Takara, Shiga, Japan). The genes encoding BAA and actin were amplified using gene-specific primers designed using the Primer Express 1.5 software package (Applied Biosystems, Foster City, CA, U.S.A.). The primers for BAA (5’-CTC TAT CGG AGC AAG AAG TTC TCG-3’ and 5’-GCC ACA CCG TTG TTA GAT ATG ATG-3’) and actin (primers described above) amplified 103-bp and 167-bp fragments, respectively. The SYBR Green I incorporated into amplified DNA was detected and the relative expression was calculated based on the standard curve from the actin gene expression. The reactions were performed three times and the mean value and standard deviation were used for the result. The expression level of the actin was considered as 1 and the relative amount of the expression level of BAA1 gene to that of actin was designated in the Figure. Preparation of antibody and immunoblotting Polyclonal antibody was prepared against N- and C-terminal peptides of BAA (MAEYGRVYKDNDEKMRRFC and CSYVRSNDESMKYA-NH2, respectively) by subcutaneous immunization of a rabbit with 200 µg peptide in Freund’s complete adjuvant. Subsequently, two booster injections containing 150 µg peptide in Freund’s incomplete adjuvant were administered at 2-week intervals. The antibody titer was estimated by indirect ELISA. Briefly, total protein and BSA (200 µg/ml) were coated in 96 well and incubated for 2 hrs at RT. After washing three times and incubated with blocking buffer overnight and washed three times. 100 µl of diluted (1:500) anti BAA-antiserum was incubated for 2 hrs and washed three times. 100 µl of diluted HRP-conjugated goat anti-mouse IgM was incubated and the excess antibody was washed out. The color development was performed by incubating with substrate for 30 min and was determined by ELISA reader at 415 nm. Total protein was extracted from transgenic lines and used for the Western blot analysis. Protein extracts from untransformed plants were used as negative controls. Protein samples were separated by electrophoresis on 15% SDS-polyacrylamide gels, followed by electrotransfer onto nitrocellulose membranes (Hybond ECL, GE Healthcare Bio-Sciences Corp., Piscataway, NJ, U.S.A.). The immunoblots were then hybridized with antibody against BAA and alkaline phosphatase detection performed, as per the manufacturer's instructions (GE Healthcare Bio-Sciences Corp., Piscataway, NJ, U.S.A.). Bioassay for transgenic plants An in planta bacterial bioassay was performed on 20 seedlings from each transgenic line as well as non-transgenic control plants at 25 days after germination. P. carotovorum ssp. carotovorum was obtained from the Korean Agricultural Culture Collection (KACC, http://kacc.rda.go.kr/eng/) at the National Institute of Agricultural Science and Technology, RDA, Suwon, Korea. Bacteria were grown for 1 or 2 d in 85-mm plates containing NA agar medium (5 g peptone, 3 g beef extract, 2 g yeast extract and 15 g Bacto-agar per L) at 28ºC. Sterile water (2 mL) was added to each plate and freshly-cultivated bacteria were scraped gently from the agar medium with a sterile glass rod. After centrifugation at 5,000 x g, the bacterial pellet was resuspended in sterile water and adjusted to OD600 = 0.1. Leaves were inoculated with the bacterial suspension by syringe infiltration at the concentration of CFU (colony forming unit) of 2 x 104/mL and 2 x 106/mL. Inoculated plants were transferred to a growth chamber and incubated at 28ºC under continuous light, then checked 12 to 96 hrs after inoculation. Control plants were treated similarly using sterile water. All the bioassays were performed three times and the pathogenicity was determined by the diameter of the disease lesion on the leaves of the plants. The mean values of the diameter were calculated and grouped to 5 groups depending on the size of the diameter. When there was no lesion, we assigned as negative (-); for lesion diameter of 1 cm, as ±; for lesion diameter of 1 ~ 2 cm, as +; for lesion diameter of 2 ~ 3 cm, as ++; for lesion diameter larger than 4 cm, as +++ at 2 days after infection. The construct of BAA1 gene encoding bromelain protease under CaMV 35S promoter (Figure 1) was transformed into A. tumefaciens LBA4404 using electroporation and transgenic plants were generated by Agrobacterium-mediated transformation (Hélias et al. 2000). After Agrobacterium tumefaciens-mediated transformation, transformants were selected for hygromycin resistance. 19 hygromycin resistant calli were obtained after hygromycin selection, from which 8 independent plants were regenerated. Transformants started to produce callus after 10 days-culture in the induction medium, an amorphous mass of cells appeared from the callus during the late period of the second generation of the culture, and many multishoots appeared from the third generation of the culture (Figure 2a). After culture in the regeneration medium and when the length of the plantlets reached 7-8 cm, the shoots were induced to form roots (Figure 2b). The rooted individuals were transferred to the pot to induce flowering by treatment at 8-10ºC for 40 days. The flowered individuals were bud pollinated to produce T1 (Figure 2c and Figure 2d). The presence of BAA1 in the 8 independent T1 lines was confirmed by PCR of genomic DNA, which generated an approx 1 Kb fragment with BAA1 gene specific primers as expected, whereas no DNA could be amplified from untransformed (control) plants (Figure 3). The HPT specific primers produced a 710 bp fragment in the transgenic plants as well as the plasmid vector. Expression of BAA in transgenic plants Semi-quantitative RT-PCR was performed to measure the expression of BAA1 in the transgenic T1 lines. All T1 lines examined showed detectable expression of BAA1 gene (data not shown). Real-time PCR was also used to measure BAA1 mRNA expression levels in the leaves of transgenic plants and all the transgenic lines showed high-levels of BAA1 expression, especially in line number 64-12-2 and 66-3-1 as shown in Figure 4. Western blot analysis of these transgenic plants identified a band with a molecular mass of about 39KD, corresponding to the predicted size of BAA1, whereas no equivalent band was observed in the untransformed control (Figure 5a). Consistent with the real-time PCR result, transgenic lines, 64-12-2 and 66-3-1, showed the highest expression of BAA1 protein. Using anti-BAA antibodies, BAA1 protein levels in the leaf tissues of T2 transgenic plant lines B64-12-2 and B66-3-1 were determined by ELISA. The amount of plant BAA1 protein was estimated by comparing the relative light units (RLU) emitted by a known amount of a BAA oligopeptide-antibody complex with that emitted by a known amount of transformed plant total soluble protein (TSP). BAA levels were expressed as a percentage of total soluble plant protein (% TSP; Figure 5b). In the leaf tissues of the B66-3 transgenic line, BAA1 represented 1.6% of total protein. Western blot analysis was also used to estimate recombinant BAA1 yield, in a side-by-side comparison with LTB (lipid transfer protein) samples of known concentration. Direct comparison of band intensities indicated that transgenic samples contained ca. 320 ng BAA1 protein in 20 µg TSP, also corresponding to ca. 1.6%, which is in agreement with the results of the quantitative ELISA. Therefore, based on the ELISA and Western blot assays, 1 g of leaf tissue (fresh weight) from the transgenic Chinese cabbage plants contained 50-55 µg of recombinant plant-synthesized BAA1 protein. Transgenic BAA Chinese cabbage plants exhibit enhanced soft rot disease resistance In order to test the resistance to bacterial pathogen, we selected Chinese cabbages of transgenic lines, 64-12-2 and 66-3-1 that showed high expression of BAA1 protein. We used the soft rot pathogen P. carotovorum ssp. carotovorum, isolated from a wild-type Chinese cabbage cultivar, to infect transgenic and wild-type control plants at the 25 days after germination. Following infection with 2 x 104 CFU, disease symptoms appeared on controls but not transgenic plants 2 days after infection as shown in Figure 6. Following infection with 2 x 106 CFU, control plants exhibited severe soft rot disease symptoms by day 2 and died by day 4, whereas the transgenic lines showed much weaker symptoms compared to control plants (Figure 6, Table 1) by day 2 and the leaves were dead by day 4. We determined the severity of pathogenicity as diameter of disease lesion of the leaves of the plants two days after infection. The mean values of the diameter were calculated and grouped to 5 groups depending on the size of the disease lesion and the result was shown in Table 1. Figure 6 and Table 1 clearly shows that transgenic plants harboring BAA1 gene exhibited weaker or no symptoms of soft rot compared to control non-transgenic plants.
This study demonstrated that constitutive expression of BAA1 bromelain gene from pineapple stems in B. rapa confers enhanced resistance to bacterial soft rot disease caused by P. carotovorum ssp. carotovorum. At this moment we do not know the mechanism of the increased resistance to soft rot. The increase in the resistance to pathogenic infection might be a result of the process of programmed death as found in the developmental programmed cell death involving proteases (Beers et al. 2000). A number of molecular engineering strategies have been employed in the pursuit of increased plant resistance towards bacterial disease; these have included the introduction of non-plant foreign genes such as those encoding antibacterial properties, or hypersensitive cell death (HCD)-associated genes (Shen et al. 2000; Verberne et al. 2000). For example, overexpression of the R2R3 MYB-related gene or AtMYB30 leads to the induction of HCD and pathogen resistance (Vailleau et al. 2002). Similarly, overexpression of the serine/threonine kinase Pti1 in tobacco accelerates HCD and resistance to P. syringae pv. tabaci infection (Zhou et al. 1995). Similar resistance to bacterial pathogens has been observed in transgenic plants expressing expI (Fray et al. 1999; Mae et al. 2001) and hrap (Ger et al. 2002; Pandey et al. 2005). Whatever the mechanism is, our results suggest that BAA1 could be utilized for protecting plants from attack by bacterial pathogens. Further investigation is required to elucidate the cysteine protease interaction and identify other proteins involved in the defense system in B. rapa. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08007t1.jpg] [ej08007f5.jpg] [ej08007f2.jpg] [ej08007f1.jpg] [ej08007f4.jpg] [ej08007f3.jpg] [ej08007f6.jpg] | ||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}