 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 2, April, 2008, pp. Bt protein rhizosecreted from transgenic maize does not accumulate in soil Ezequiel Margarit1 , Martín I. Reggiardo2 , Hugo R. Permingeat*3 1Centro de Estudios Fotosintéticos y Bioquímicos,
Suipacha 570, 2000 Rosario,
Argentina
Tel: 54 341 437 1955
Fax: 54 341 437 0044
E-mail: margarit@cefobi.gov.ar Financial support: This work was supported by Rotary Club Ciudad de Buenos Aires. Received March 14, 2007 / Accepted August 13, 2007 sc Code Number: ej08018 Abstract The persistence of CryIAb protein rhizosecreted in soil is important in the assessment of its environmental risk. Here we report that CryIAb protein from transgenic maize does not accumulate at high levels in soils. Levels of CryIAb protein rhizosecreted by three maize transgenic events (BT11, MON810 and 176) were studied in hydroponic cultures and found only in the MON810 and BT11 events but not in event 176 or control plants. Under field conditions, the cryIAb gene and a basal level of CryIAb protein was detected in soils from plots cultivated with transgenic and non-transgenic maize, possibly from Bacillus thuringiensis present in the soils. Keywords: cryIAb gene, CryIAb protein, ELISA, genetically modified organisms, PCR, Zea mays (L.).
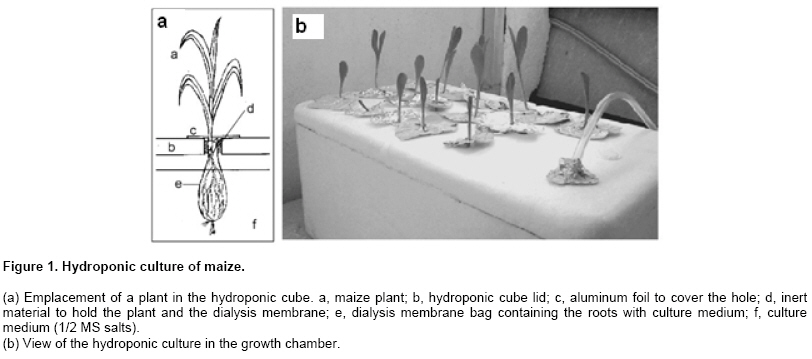
The area covered with transgenic crops is increasing yearly worldwide. In 2005, 90 million hectares were cultivated with genetically modified soybean (60% of total soybean acreage), maize (24%), cotton (11%), and canola (5%). Resistance to herbicides and insects were the traits introduced into these species (71% and 18%, respectively). The two dominant genetically modified (GM) crop/trait combinations in 2005 were herbicide-tolerant soybean (54.4 million ha, 60% of the global acreage, grown in seven countries) and Bt maize (9.1 million ha, 13% of the global acreage, grown in nine countries) (James, 2005). Despite the increasing adoption of GM crops by farmers, the public perception of transgenic crops is critical. Two main topics are the center of a protracted debate in society: environmental impacts and health risks of growing and eating GM organisms and derivates. Different regulatory systems were developed and implemented to assess these risks in countries where transgenic crops are grown. The major goals of these systems include actions necessary to ensure safe products and to engender public trust (Jaffe, 2004). Three events of Bt transgenic maize have been authorized for commercialization in Argentina and Europe. These events (176 from Novartis, MON810 from Monsanto, and BT11 from Syngenta) contain different sequences of a synthetic cryIAb gene (AGBIOS, 2006). Root exudation is a phenomenon contributing to the communication and interaction between plants and soil organisms. Root exudates have been grouped into low and high molecular weight compounds: e.g., amino acids, organic acids, sugars, phenolics, mucilage (polysaccharides), proteins, respectively (Walker et al. 2003). One of the compounds secreted by transgenic maize plants is the CryIAb protein (Saxena et al. 1999; Saxena and Stotzky, 2000; Saxena et al. 2002a), which remains active in soil by binding to clays and humic acids for at least 234 days, the longest time studied (Tapp and Stotzky, 1998), and moves vertically to different depths according to the clay content (Saxena et al. 2002b). The CryIAb protein derived from transgenic crops may be introduced to soil by root exudates, pollen during tasseling, and plant residues after harvesting (Saxena et al. 1999). However, this insecticidal protein has no apparent effect on earthworms, nematodes, protozoa, bacteria, and fungi in soil (Saxena and Stotzky, 2001a). The degradation of CryIAb proteins in soil has been determined by immunology and bioassays with susceptible insect species (Tapp and Stotzky, 1998; Herman et al. 2001; Head et al. 2002; Hopkins and Gregorich, 2003; Zwahlen et al. 2003), as well as degradation of Bt maize biomass (Flores et al. 2005). Gene transfer from the environment to different organisms has been widely described (reviewed by Bertolla and Simonet, 1999), including heterologous DNA fragments present in transgenic plants. This also addresses the importance of assessing the presence of CryIAb gene on soil, since horizontal gene transfer to soil microorganisms or even plants cannot be completely excluded. In this paper, we quantified the amount of CryIAb protein rhizosecreted by different events of transgenic maize grown under hydroponic condition and studied the presence and origin of CryIAb protein and DNA in soil cultivated with transgenic and nontransgenic maize. Materials and Methods Soil samples from plots cultivated with transgenic maize (32G63MG -event MON810- and Chalten TD -event 176-), and their respective nontransgenic near-isolines (hybrids 30F82 and Pucara, respectively) were examined for the presence and persistance of cryIAb genes and CryIAb protein. Samples were taken from plots (10 Ha-each) in a 40-ha field near San Genaro (Depto. San Jerónimo, Santa Fe, Argentina, 32º S Lat and 61º W Long). The soil was a Typical Argiudol Clason Serie (25.3% clay, 72.8% silt, 1.9% sand, 2% organic matter). This field had never been cultivated with transgenic maize before. Triplicate samples from the first 15 cm from the soil surface and 20 cm in diameter, centered in the maize plant, were collected from each plot at 3-month intervals (in February -before harvesting the crop-, May, August, and November, 2002, and in March, 2003) and pooled before carrying out the analyses. The samples were cleaned from visible plant tissue residues and homogenized to a fine powder in a mortar to extract DNA and proteins for PCR and ELISA analyses, respectively. DNA isolation, and PCR analysis Ground soil (15 g) was mixed with 15 mL of extraction buffer (100 mM Tris-HCl, pH 8, 100 mM sodium EDTA, pH 8, 1.5 M NaCl) in centrifuge tubes (SS34, Sorval) and incubated at room temperature for 20 min, with manual stirring every 3 to 5 min. Then, 1.5 mL of 20% SDS was added to each sample, which was homogenized, incubated at 65ºC for 1 hr with gentle shaking every 5 min, and centrifuged at 5,000 x g for 10 min. Supernatants were collected, and the soil pellet was reextracted with another 5 mL of extraction buffer, incubated at 65ºC for 10 min, and centrifuged as above. Supernatants were transferred to SS34 tubes and 1 volume of precipitation solution (30% Polyethylen glycol –PEG- 8000, 1.6 M NaCl) was added. The mixture was incubated at room temperature for 2 hrs and centrifuged at 5,000 x g for 20 min. The pellets were resuspended in 100 µL of TE (10 mM Tris-HCl, pH 8, and 1 mM sodium EDTA, pH 8). To purify the DNA from proteins and polysaccharides contamination, 7.5 M potassium acetate was added to a final concentration of 0.5 M. Samples were kept in ice for 5 min and centrifuged at 16,000 x g for 30 min at 4ºC. The supernatant was transferred to a new tube and extracted twice with 24:1 chloroform:isoamylic alcohol. DNA was precipitated by adding 0.6 volumes of isopropanol and incubated at 4ºC for 1 hr. DNA was pelleted by centrifugation at 16,000 g for 15 min and resuspended in 75 µL of TE buffer. DNA was quantified fluorometrically with the fluorochrome, Hoesch H33258 (Ausubel et al. 1994). DNA used as positive and negative controls for analysis was isolated from transgenic and non-transgenic kernels, respectively, according to Lipp et al. (1999). End-point PCR was carried out in a Perkin-Elmer 2400 (Applied Biosystem, Foster, CA) or a Sontec (Buenos Aires, Argentina) thermocycler for amplification of cryIAb and maize zein genes. The zein gene was also amplified as an internal control in parallel reactions where maize DNA was added at 10 ng per tube (Permingeat et al. 2002). Reactions took place in a final volume of 25 µL, containing MgCl2 (1.5 mM for cryIAb and 2 mM for zein), 0.8 mM dNTPs, 0.5 µM primers, 1.25 units of Taq Polymerase, and 1 X PCR buffer. All reagents were from Promega (Madison, WI) except for the primers, which were synthesized by Biosynthesis Inc. (Lewisville, TX), and 3-10 ng of DNA from soil samples. Conditions for amplification were: denaturing of DNA at 94ºC for 5 min, 40 cycles of 30 sec at 94ºC, 30 sec at 58ºC for cryIAb gene or 60ºC for zein gene, 30 sec at 72ºC, and a final extension at 72ºC for 5 min. Amplification products were electrophoresed in 2% agarose gels for 1 hr at 40 mA, and stained with ethidium bromide for visualization. Primers used for the amplification fragments of cryIAb fragment amplification were 5’- ATG GAC AAC AAC CCC AAC ATC –3’ and 5’- AAA GAT ACC CCA GAT GAT GTC –3’ (Permingeat et al. 2002); for amplification fragments of zein, the primers were 5’- GCT TGC ATT GTT CGC TCT C -3’ and 5’- CGA TGG CAT GTC AAC TCA TTA -3’ (Studer et al. 1997), giving fragments of 204 and 447 bp, respectively. The PCR analyses were performed in triplicate. Soluble total proteins (STP) were extracted from 1 g of soil with 1.25 mL Extraction/Dilution Buffer (EnviroLogix, Inc.) according to the protocol indicated by Saxena et al. (1999). The mixture was incubated at 4ºC for 90 min with shaking, centrifuged (10,000 x g), and the supernatant was recovered. One 0.5-5 µL aliquot of the extract was used to quantify STP spectrophotometrically by the Bradford method, using bovine serum albumin for the standard curve. Seeds from maize hybrids 32G63MG (event MON810), Chalten TD (event 176), Chalten TD Max (event BT11) and AX82 (nontransgenic) were surface sterilized with 70% ethanol (1 min) and 2.5% NaClO (20 min) and then washed three times with sterile distilled water. Seeds were germinated between cotton and paper in sterile Petri dishes. The hydroponic cube was made from a styrofoam box (35 cm long, 20 cm wide, and 15 cm high) previously disinfected as were the seeds plus a 2 hrs exposure to a UV lamp (254 nm). Holes of 1-cm diam. were made in the lid of the box. Roots of 15 5-6 days-old intact plantlets were bagged in a 6-8 kDa cut-off dialysis membrane (Spectrapor, Sectrum Medical Industries, Inc.) (Figure 1a). Plantlets with bagged roots were placed in the holes of the lid and immobilized with a piece of foam at the hypocotile level. The bags and the cube were filled with sterile medium containing one half Murashige and Skoog salts, pH 6. Medium was added weekly to the cube to keep the initial level, but not exchanged. A fish-tank air insuflator was used to prevent root anoxia. Incoming air was filtered through a sterile 50 mL-Falcon tube containing autoclaved cotton. Plants were cultured for 3 and 5 weeks in a growth chamber (26ºC, 16:8 hrs photoperiod) under aseptic conditions as shown in Figure 1b. Every week, three plants were sampled to evaluate growth and to harvest the rhizosecreted proteins. Medium in the bags (approximately 10 mL) was centrifuged at 5,000 g at 4ºC for 30 min to remove suspended particles, and the supernatant was concentrated to 100-350 µL using a YM-30 Centricon (Amicon Bioseparations, Millipore) (cut off: 30 KDa). Proteins were quantified by the Bradford method, and the medium was lyophilized. Then, they were resuspended in 100 µL of extraction/dilution buffer from the EnviroLogix CryIAb/c Plate Kit. An enzyme linked immunosorbent assay (ELISA) kit (CryIAb/Cry1Ac Plate kit AP003, from EnviroLogix, Portland, ME) was used to quantify the CryIAb protein in soil extracts from Bt and non-Bt corn fields, and exudates from hydroponic-cultured transgenic and non-transgenic maize plants, according to the manufacturer protocol. The pH of the extraction buffer was 7. Neutral and slightly acidic pH values reduce the amount of humic acid and organic matter in the extracts (Murase et al. 2003) and also the solubility of the native Bt crystal toxin present in soil. Proteins measurements were made in a Dynex MRX plate reader (Dynex Technologies) at 450 and 630 nm. A standard curve of the OD of each known concentration of CryIAb provided with the kit (0.5, 2.5 and 5 ppb) was included in the assays. The result of each unknown sample was calculated as ng CryIAb protein/g of dry-weight soil or as pg CryIAb protein/plant, respectively. Each sample was analyzed in triplicate. Isolation and characterization of Bacillus thuringiensis from soil Distilled water (10 mL) was added to 2 g of soil from a non-Bt corn fields. The suspension was shaked, heated for 30 min at 60ºC, and allowed to stand for 1 hr at room temperature. An aliquot of the supernatant was diluted 10-fold, and 40 µL of the dilution were plated onto LB medium, and incubated overnight at 30ºC, as described in http://www-biology.ucsd.edu/labs/aroian/protocols/btspore.html. Isolation was performed in triplicate. Bacterium colonies were picked and resuspended in 10 µL of sterile destilled water. This dilution was used as template (2 µL) for PCR, in which the cryIAb native gene was recognized by the primers, 5’-CGG GAT TAG AGC GTG TAT G – 3’ and 5’- TGG CCC CGA AAA CCC TAC – 3’, which amplify a 356-bp PCR fragment. Conditions for PCR were: 95ºC for 3 min; 30 cycles of DNA denaturing at 95º for 30 sec, annealing at 51ºC for 30 sec, and extension at 72ºC for 30 sec, and a final extension at 72ºC for 3 min. Reagents were added at the following concentrations: 1.5 mM MgCl2 0.8 mM dNTPs, 0.5 µM primers, 1.25 units of Taq Polymerase, and 1X PCR buffer, in a final volume of 25 µL/tube. Results In a 3-week period of hydroponic culture, the CryIAb protein was detected and quantified immunologically in hybrids 32G63MG (event MON810) and Chalten TD max (event BT11), but not detected in the hybrid Chalten TD (event 176) and control plants. The total exudated protein from all genotypes was comparable (Table 1). The hybrid 32G63MG (event MON810) was kept in hydroponic culture for two more weeks, and the CryIAb protein increased in the rhizosecreted exudate in a linear manner, yielding 336 pg from about 67 ug of exudated protein per plant (Table 2). The CryIAb protein was also quantified in samples of root tissues from 3 weeks old plants from the three events grown in hydroponic culture, using the same commercial ELISA kit (EnviroLogix), and a positive correlation between the CryIAb rhizosecreted and CryIAb protein present in root tissue was found (Table 1).
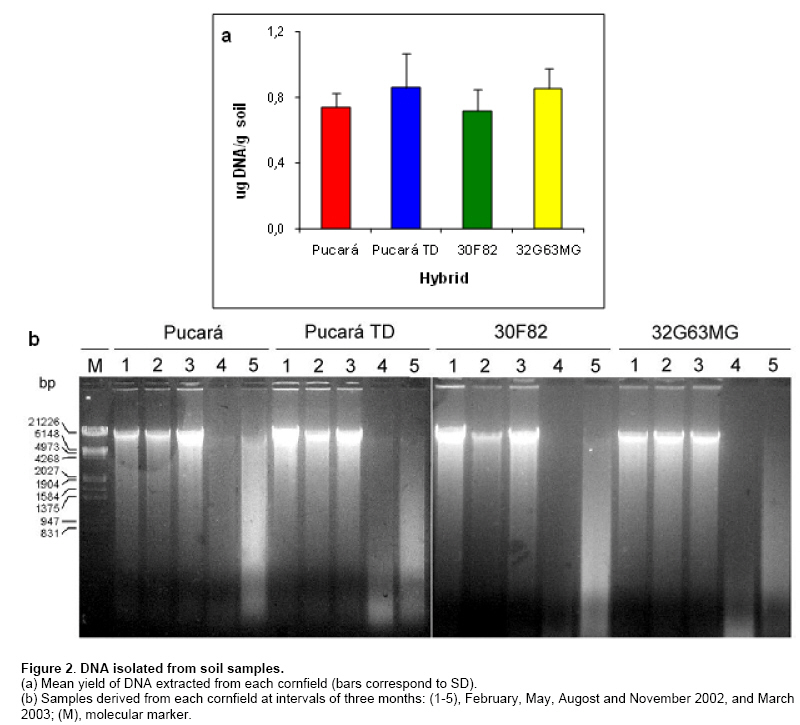
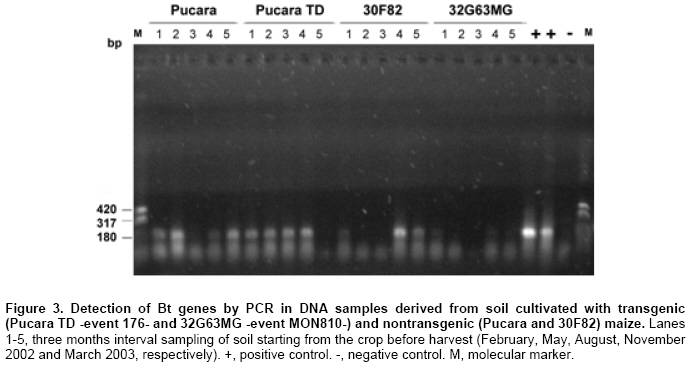
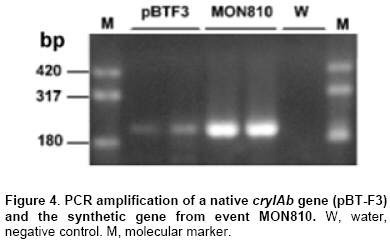
As mentioned in Materials and Methods, DNA and proteins were isolated from soil samples from transgenic and nontransgenic cornfields to study the presence and persitence of the cryIAb gene and CryIAb protein. The yield of DNA extracted from soil samples was low according to results previously reported (Douville et al. 2007). An average of 0.8 µg/g soil was fluorometrically quantified. A wide diversity of molecular weight DNA was observed for most of the DNA extracts, probably corresponding to mainly intact DNA from soil microorganisms and degraded DNA from plant residue and other organic matter (Figure 2). Other methods of DNA purification (ISO/TC 34 WG7 N23, 2001; Miller et al. 1999) were also tested with similar results (data not shown). Samples of DNA were first analyzed to assess their quality for PCR amplification. DNA (10 ng) from a maize kernel was added to an aliquot of each sample of soil DNA (intentional contamination), and zein amplification was carried out as previously reported (Permingeat et al. 2002). Sixteen of 20 samples gave the expected band corresponding to the zein gene either added or present in soil (not shown). Four soil samples (Pucará-3, Pucará TD-5, 30F82-3, 32G63MG-3) did not yield the zein amplification fragment, suggesting the presence of inhibitors of the Taq polymerase in the soil DNA sample. DNA extraction of good quality from soil samples is not easy due to the copurification of humic acids and organic matter. This fact was observed as a brown color of the solution containing the DNA and the inhibition of the Taq polymerase. A dilution (1:100) of the DNA extract was used for PCR, using 3-10 ng of DNA as template for the assays and diluting the inhibitors to find the band of amplification. The 20 DNA samples isolated from soils cultivated with either transgenic and nontransgenic maize were analyzed for the presence of the cryIAb gene. Fifteen of them gave the expected band of 204 bp, corresponding to the cryIAb gene, after PCR amplification (Figure 3). These results were confirmed by Southern blots of the amplification products (data not shown), where one more sample gave the expected amplicon. The four negative samples corresponded to those identified previously as inhibited. Inasmuch only one replicate was included in the figure, analyses were done per triplicate, and results were highly consistent. The amplification products corresponding to the cryIAb gene were obtained after amplification of DNA samples purified from soil cultivated with transgenic and non-transgenic maize. Why did the soil cultivated with nontransgenic maize have the Bt gene? To highlight this question, the same DNA samples were assayed for the presence of the zein gene from maize tissues. There were no bands in the gels or in the Southern blots (data not shown). To determine whether the primers used to amplify the synthetic cryIAb gene from transgenic maize events could also amplify the native gene of B. thuringiensis, a PCR experiment was performed. Figure 4 shows the amplification bands from both the native and the synthetic (from event MON810) genes. The native gene used as a template was previously cloned into plasmid pBT-F3 in our laboratory. A BLAST analysis of these primers revealed a limited number of hits with other organisms different than B. thuringiensis, and with a much higher E value. B. thuringiensis was also isolated from soil samples from plots cultivated with nontransgenic maize. A PCR assay using primers that recognize a fragment of the native cryIAb delta-endotoxin gene revealed that 4 out of 10 colonies analyzed yielded the expected fragment (data not shown), indicating the presence of the bacterium in this soil. The BLAST analysis of these oligonucleotides did not find any hit with other organisms than Bacillus thuringiensis. The protein extracts showed a slow browning, suggesting contamination with humic acids. The total protein extracted ranged between 18.8 and 29.1 µg/g of soil, and the CryIAb protein was detected with the ELISA kit (limit of detection = 0.5 pg), although at levels below the limit of quantification (50 pg). Although a 10-fold concentration of the soil protein extracts would have been required to fit in the standard curve for precise quantification. The multifactorial ANOVA (p ≥ 0,01) revealed no differences between plots cultivated with transgenic and nontransgenic maize or between plots cultivated with different transgenic maize events (176 and MON810), during the 15-month period of the experiment (Table 3).
Discussion The accumulation and persistence of the insecticidal toxins from B. thuringiensis in soil may result in environmental hazards, such as toxicity to non-target species and the selection of toxin-resistant target species (Stotzky, 2005). In addition, the risk for horizontal gene transfer to other soil microorganisms should not be completely excluded (Bertolla and Simonet, 1999). Several studies have determined that the insecticidal protein can be adsorbed to clay particles and humic acids (e.g., Tapp and Stotzky, 1995; Crecchio and Stotzky, 1998; Tapp and Stotzky, 1998), resulting in insecticidal activity for long periods (Saxena et al. 1999). Root exudation of insecticidal CryIAb protein from transgenic maize was first reported by Saxena and coworkers (Saxena et al. 1999; Saxena and Stotzky, 2000; Saxena et al. 2002a; Saxena et al. 2004). A major band of 66 kDa migrating in SDS-PAGE was reported in exudates from event BT11 but not from the near-isogenic non-Bt corn after 7 and 15 days in hydroponic culture (Saxena et al. 1999). The presence of CryIAb protein in soil cultivated with 12 transgenic corn hybrids, representing events 176, MON810, and BT11, was reported (Saxena et al. 2002a), whereas no toxin was detected in soils in which near-isogenic non-Bt corn was grown. In the same study, no differences were found in the level of exudation of the toxin (evaluated by bioassays and immunological assays) between hybrids containing different transformation events. However, we detected exudation from event MON810, at a level of 94 pg, and from event BT11 at a level of 19 pg of CryIAb protein per plant in three weeks, but not from event 176 in hydroponic culture. We found that the levels of CryIAb protein exudated correlate with the levels of Bt expression in root tissue. Expression of Bt in roots for event 176 was reported to be lower than 0.01 µg/g (AGBIOS, 2006), which is in agreement with our result (Table 1). The promoters used to control Bt expression in this event were pollen- and green tissue-specific. As no information was available in the literature for Bt expression in the roots of events MON810 and BT11, where the expression is controlled by the constitutive CaMV35S promoter controls the expression, we quantified it with an ELISA kit. We found evidence indicating a stronger expression in the roots (150-200 times) in these events than in event 176, which contains tissue specific (non-root) promoters Table 1. At the field level, B. thuringiensis insecticidal protein and DNA was found in most of the samples, regardless the time of collection or the hybrid present (either Bt or non-Bt). Our results suggest that the source of CryIAb protein and DNA found in soil samples was other than transgenic Bt plants, especially because the field used for these experiments had never been cultivated with a transgenic Bt crop, nor sprayed with Bt-based insecticidal formulations, which can be another source of Bt (Vettori et al. 2003). We provide evidence suggesting that CryIAb protein and DNA may arise from cells of B. thuringiensis already present in soils, even though we can not completely exclude the plant origin of the gene in soils with Bt and non Bt maize, as reported recently by Douville et al. (2007), using quantitative real time PCR. The negative PCR results obtained when an endogenous zein gene from maize in soil samples was amplified after extensive removal of plant residues suggested that free DNA from maize was either not present in the soil or it was too degraded to be amplified by PCR, even in samples collected postharvest. Degradation of plant tissue was shown by Ceccherini et al. (2003), who reported that most of the DNA is degraded inside plant cells during the senescence of the tissue while nucleases retain their activity. Different results have been reported about the persistence of the CryIAb protein in soils. Although some authors reported extended persistence and insecticidal activity after addition of purified protein, commercial insecticidal preparation or plant tissue from Bt maize (e.g., Tapp and Stotzky, 1998; Vettori et al. 2003; Zwahlen et al. 2003) others reported a rapid degradation of the CryIAb protein in soils cultivated with transgenic crops using a bioassay (Herman et al. 2001) and both ELISA and bioassays (Head et al. 2002). The long persistence of CryIAb protein was reported by Zwahlen et al. (2003) in soils from Switzerland, where plant residues of Bt maize were incorporated. They conclude that low temperatures play an important role to diminish the degradation rate, while rains do not seem to have a strong influence. Saxena and Stotzky (2001b) reported that all three transformation events (176, MON810, and BT11) have a higher lignin content than their near-nontransgenic isolines. This fact may contribute for a longer persistence of plant residues derived from transgenic maize in soils, as it was reported by Flores et al. (2005). On the other hand, the CryIAb protein not bound on surface-active particles seems to be labile to microbial degradation (Koskella and Stotzky, 1997). Baumgarte and Tebbe (2005) reported that immunoreactive CryIAb protein was detected infrequently in soil, especially in the first year of assay with a Bt maize, but they did not detect this protein in control plots with nontransgenic cultivars. Recently, Sun et al. (2007) reported that the CryIAb protein persisted in soil for at least 56 days after incubating a silty loam soil with Bt cotton tissues. We detected CryIAb protein immunologically in control plots at 30 pg/g of soil, which can be considered a basal level. In contrast, we found no accumulation of CryIAb protein during the growing season of Bt maize or one year after harvest. A recent report indicated that Bt maize does not accumulate in soil even after 3 years of sustained Bt corn cultivation (Dubelman et al. 2005). Our results indicate that there was no significative accumulation of CryIAb protein in soils under the conditions of this study. Moreover, the amount of CryIAb protein detected seemed to derive from B. thurigiensis already present in soils. The basal level of the bacterium in soil was not reported and discussed by authors that evaluated the exudation of CryIAb protein (Saxena et al. 1999; Saxena et al. 2002a) and its degradation (Herman et al. 2001; Head et al. 2002). Considering that B. thuringiensis is a common inhabitant of most soils around the world (Martin and Travers, 1989), a sensitive method, such as ELISA, allowed us to detect this basal level. To confirm this result, colonies of B. thuringiensis were isolated and identified by PCR from soil never cultivated with a Bt crop, nor sprayed with Bt insecticidal formulation. In summary, we quantified the level of rhizosecreted CryIAb protein from transgenic maize in hydroponic culture and found that it correlated with the level of expression in the roots driven by the promoters present in the transcriptional units of the transgenic events. Despite this, no differences in the level of immunoreactive CryIAb protein were found in soils cultivated with transgenic and nontransgenic maize up to one year after harvest. However, a basal level of CryIAb protein was detected, which may have resulted from B. thuringiensis already present in the soil. Further studies are needed to confirm that the bacterium already present in soil produces enough protein to be detected. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08018f2.jpg] [ej08018f1.jpg] [ej08018f4.jpg] [ej08018f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}