 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 2, April, 2008, pp. Design and expression of a retro doublet of cecropin with enhanced activity Mauricio Díaz1 , Gloria Arenas2, Sergio H. Marshall*3 1Laboratorio de Genética e Inmunología
Molecular,
Instituto de Biología,
Pontificia Universidad Católica de Valparaíso,
Av. Brasil 2950, Valparaíso, Chile,
Tel: 56 32 273119
Fax: 56 32 596703
E-mail: mauricio_diazruiz@yahoo.com Financial support: This work was supported by the Copec-UC foundation and MECESUP. Received July 27, 2007 / Accepted January 9, 2008 Code Number: ej08020 Abstract Novel doublet molecules of cecropin A from Drosophila melanogaster were designed and constructed combining the regular (CECdir) with the inverted (CECret) coding sequence of the standard CEC A1 gene resulting in the following configurations: CECdir-CECret and CECret-CECdir. These two recombinant molecules were generated using a three-primer driven PCR reaction yielding composite single functional aminoacidic molecules with the coding sequences of CECdir linked in frame with the coding sequence of CECret and vice versa. In order to obtain these constructions, a retropeptide DNA-coding sequence was chemically synthesized to match the expected polarity of the newly generated CECret sequence. Both doublet antimicrobial peptides (drAMPs) were cloned in the T7 promoter driven expression plasmid pET27b+ and expressed in E. coli BL21 without any fusion protein. Only the former recombinant peptide was expressed and purified from cell extracts and its specific activity against two different bacteria showed to be higher than those displayed by their monomer parental counterparts. Keywords: antimicrobial peptides, Escherichia coli, expression.
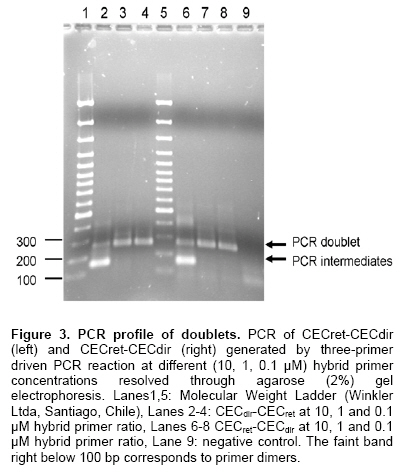
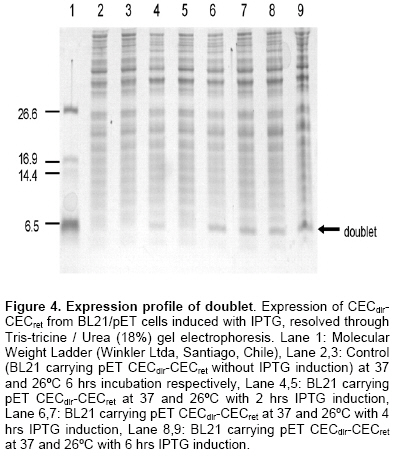
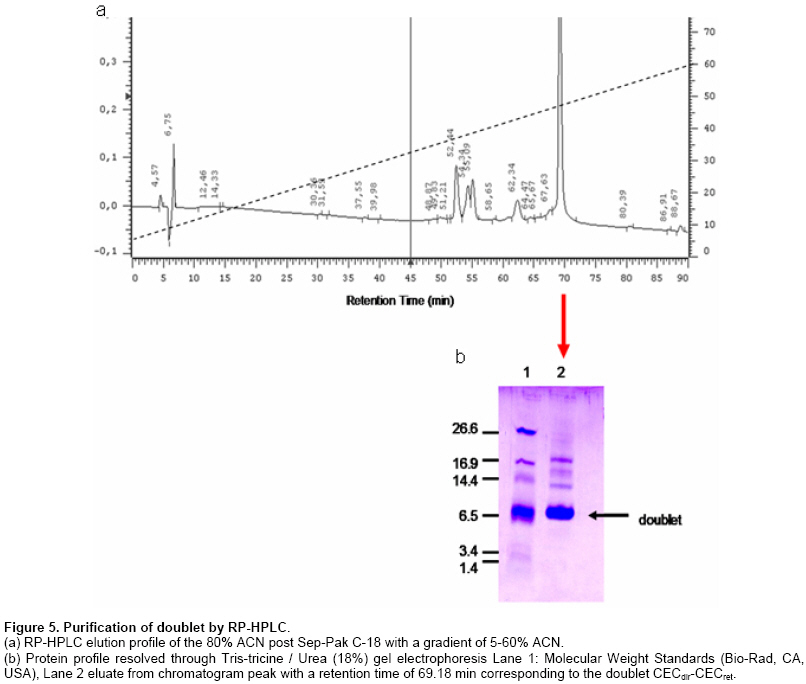
The discovery of natural peptides possessing a broad range of antimicrobial activities, synergy with conventional antibiotics and rapid killing action, has drawn attention as potential therapeutics (Zasloff, 2002). However, up to now the classical way to have access to these molecules has been their isolation from host organisms, which requires large amount of material with very low yields of functional peptides (Mercado et al. 2005). Other alternatives explored have been their chemical synthesis (Andreu et al. 1983) or their expression in heterologous systems following their cloning from origins (Piers et al. 1993; Zhang et al. 1998). Most antimicrobial peptides (AMPs) act by penetrating bacterial membranes and promoting cell lysis, mechanism of action which is known to be critically dependent on their cationic and amphipathic nature (Yount and Yeaman, 2005). It has also been suggested that in some cases above a sufficiently high concentration of adsorbed peptides the mechanism depends on a peptide dimerization or oligomerization while they interact with the membrane of bacteria, which in turns results in, pore formation (Zemel et al. 2005). Studies using cross-linked disulfide-dimerized melittin and magainin analogssupport the idea that an interfacially adsorbed, self-associated dimer acts as a precursor or a meta-stable intermediate in the pore-formation mechanism (Hristova et al. 2001; Dempsey et al. 2003). The spontaneous formation of disulphide-linked dimers or the engineering of dimers through crosslinking (Tencza et al. 1999; Dempsey et al. 2003) showed enhanced membrane permeabilization activity compared with the corresponding monomer, especially at low concentrations, where pore formation by the dimerized peptide occurs well below the monomers threshold concentration. It was suggested that membrane perturbation resulting from the formation of the peptide dimer is the essential event that triggers the change in peptide orientation and the formation of a pore (Hristova et al. 2001). Evidence also shows the existence of natural homo and heterodimers like distinctin from the frog P. distincta (Batista et al. 2001) and cryptdin-related sequence peptides in mouse intestinal tissue (Hornef et al. 2004). On the other hand, it has also been observed that a mixture of AMP classes do interact with each other in a synergic way enhancing their specific activity (Luders et al. 2003). Cecropins are linear peptides, isolated initially from insects, which present a strongly basic NH2 terminal region, enriched in lysine, and a large COOH-terminal hydrophobic region (Holak et al. 1988; Boman, 1991). Their basal structure is alpha-helix and contains a strongly cationic region that permits interaction with the microorganism membrane and a large hydrophobic tail that induces the microbial lysis by alteration of the permeability of the membrane (Durell et al. 1992; Silvestro et al. 1997). Cecropin A from Drosophila melanogaster is a polycationic peptide that is active against a wide range of both Gram-positive and especially over Gram negative bacteria (Zasloff, 2002; Boman, 1998). Studies on molecular evolution of cecropins (Ramos-Onsins and Aguadé, 1998) demonstrated the importance of this multigene family in different species of invertebrates. Based upon these facts we designed and constructed a number of doublets of cecropin hoping to achieve, in a single molecule, the best efficiency of their parental counterparts and their recombinant expression in E. coli, considering that knowledge regarding the relationship between peptide structure and function as well as their mechanism of action is being applied in the design of antimicrobial peptide variants as potential novel therapeutic agents (Jenssen et al. 2006). Materials and Methods Bacterial strains, plasmids and growth conditions The pET27b+ (Novagen) vector was used for the expression of doublets CECdir-CECret, CECret-CECdir, as well as monomers CECret and CECdir. In these plasmids transcription of genes is controlled by the isopropil-β-D-thio-galactopyranoside (IPTG)-inducible T7/Lac promoter. E. coli TOP10 (Invitrogen) was routinely used as the host for propagation and maintenance. E. coli BL21(DE3) “codon plus” (Stratagene) was used as the host strain for protein expression. E. coli strains were grown at 37ºC in Luria-Bertoni (LB) broth or agar, and 50 µg/ml of Chloramphenicol and 75 µg/ml of Streptomycin was added to the media for strain BL21(DE3). Kanamycin (50 µg/ml) was added to the media when TOP10 and BL21(DE3) were transformed with the expression plasmids. Pathogenic bacterial strains Vibrio anguillarum and Vibrio ordalii were grown at 26ºC in Trypticase Soy Broth (TSB) and peptone-NaCl (PSB) respectively. Construction of CECret, CECdir-CECret and CECret-CECdir expression plasmids Previous two doublet constructions, a retropeptide-coding sequence of Cecropin A was designed to match the expected polarity for each coding triplets yielding the novel CECret gene and was later synthesized by GenScript. The resulting “retropeptide” is made up of L-amino acids in which the amino acid residues are assembled in opposite direction to the native peptide (Merrifield et al. 1995). Doublets with the configurations [(NH2)head-tail(COOH)]▼[(COOH)tail-head(NH2)] like CECdir-CECret and [(COOH)tail-head(NH2)]▼[(NH2)head-tail(COOH)] like CECret-CECdir were then generated. Each doublet correspond to a single peptidic molecule representing the coding sequence of the original monomer CEC linked in tandem with its inverted coding sequence resulting in an enantiomer molecule at the amino acid level. The sequences of the mature peptides CECdir and CECret and its coding sequences are shown in Figure 1. A DNA fragment encoding the mature domain of Cecropin A was obtained by PCR amplification of pBSCecA DNA, that carries the standard CEC A1 gene (CECdir) used in this study (a kind gift from Dr. Dan Hultmark). The sequence of the 5’ primer used for amplification (5’-GGGTGGCTGAAGAAAATTG-3’) corresponds to codons Gly+1 to Ile+6. The 3’ primer (5’-ACCTCGGGCAGTTGCGGCG-3’) is complementary to codons Ala+35 to Gly+40 (without stop codon). A DNA fragment encoding the mature domain of retro-cecropin A was obtained by PCR amplification of pUC57RET DNA, that carries the retro CEC A1 gene (CECret) (obtained from GenScript), using the 5’ primer (5’-GGTCGAGCCACTGCAGCCG-3’) and 3’ primer (5’-CCCCCACAGCTTTTTAATG-3’). PCR was performed under the following conditions: incubation at 94ºC for 10 min, 30 cycles incubation at 94ºC for 30 sec, 55ºC for 30 sec, 72ºC for 45 sec, and final incubation at 72ºC for 10 min. The PCR product of CECdir and CECret were analyzed by agarose gel electrophoresis (2%) and purified using Rapid Gel extraction systems (Marligen). The resulting fragments were used as templates for a three-primer driven PCR reaction, where the coding sequence of CECdir was linked in frame with the coding sequence of CECret in the case of CECdir-CECret, and the coding sequence of CECret was linked in frame with the coding sequence of CECdir in the case of CECret-CECdir. For the construction of CECdir-CECret PCR amplification was done with a hybrid primer complementary to the last 7 codons of CEC and the first 7 codons of CECret (5’-GACGGCTGCAGTGGCTCGACCACCTCGGGCAGTTGCGGCGAC-3’), as well as the 5’ primer (5’-GAGATATACATATGGGGTGGCTGAAGAAAA-3’) corresponding to codons Gly+1 to Ile+6 of CECdir, and the restriction site for NdeI, and the 3’ primer (5’-GCGGATCCTCATCACCCCCACAGCTTTTTAA-3’) complementary to codons Ile+35 to Gly+40 of CECret, with a stop codon and restriction site for BamHI. The same procedure was used for the construction of CECret-CECdir using a hybrid primer complementary to the last 7 codons of CECret and the first 7 codons of CECdir (5’-GCCAATTTTCTTCAGCCACCCCCCCCACAGCTTTTTAATGCC-3’), as well as the 5’ primer (5’-GAGATATACATATGGGTCGAGCCACTGCAG-3’) corresponding to codons Gly+1 to Ala+6 of CECret, and the restriction site for NdeI, and the 3’ primer (5’-GCGGATCCTCATCAACCTCGGGCAGTTGCGG-3’) complementary to codons Ala+35 to Gly+40 of CECdir, with a stop codon and restriction site for BamHI. PCR amplifications were performed under the following conditions: incubation at 94ºC for 10 min, 30 cycles incubation at 94ºC for 30 sec, 65ºC for 30 sec, 72ºC for 45 sec, and final incubation at 72ºC for 10 min, using different concentrations of the hybrid primer and 1 U of Taq DNA polymerase (Invitrogen, Brazil). The resulting doublet fragments and the corresponding monomers were analyzed by agarose gel electrophoresis (2%) and precipitated by standard methods (Sambrook et al. 1989) and subject to restriction digestion with NdeI and BamHI. The resulting fragments were precipitated again and ligated between the NdeI and BamHI restriction sites within the multiple cloning site of pET27b+. Ligation mixtures were used to transform competent TOP10 cells (Invitrogen). Plasmid DNA was isolated by miniprep kit (Qiagen). Plasmids were sequenced to confirm the identity of the constructs. Expression plasmids were subsequently used to transform competent BL21(DE3) cells (Stratagene). Expression and purification of doublets E. coli BL21(DE3) pET/CECdir-CECret and pET/CECret-CECdir and the corresponding monomers pET/CECdir and pET/CECret transformation single colonies were used to inoculate 10 ml LB Broth supplemented with antibiotics at 37ºC and grown overnight. Next morning 10 ml of each culture (20 µl/ml LB) were added to 500 ml LB Broth without antibiotic selection and incubated for 2 hrs until the A600 reached 0.3. After collection of preinduction samples, IPTG was added to the cultures to a final concentration of 0.5 mM and incubated for up to 4 hrs before sampling. Samples were separated by centrifugation at 7,000 g x 10 min at 4ºC into cell pellet and supernatant fractions. Samples of cell pellets solubilized directly in sample loading buffer containing sodium dodecyl sulphate (SDS) were analyzed on 18% Tris-tricine/Urea-polyacrylamide gels (Schagger and von Jagow, 1987) and stained with R-250 Coomassie Blue. Samples were also subjected to peptide extraction. Cell pellets were treated with lyses buffer (Tris-HCl 50 mM, NaCl 100 mM, pH 8), lysozyme (100 µg/ml) and vortexed, incubated at room temperature for 30 min, subjected to ultra-sonication on ice (3 x 20 sec) and centrifugation at 11000g x 30 min at 4ºC and supernatant fractions were recovered. Samples were purified by hydrophobic interaction chromatography in Sep-Pak C-18® columns pre-equilibrated with acidified water (0.05% trifluoroacetic acid in ultrapure water UPW) and washed with acidified water and eluted with 5 and 80% acetonitrile (ACN). Eluted fractions were lyophilized and reconstituted in UPW. Total protein concentration was determined by bicinchoninic acid (BCA) protein assay (Pierce) (Smith et al. 1985). Purification was performed on a RP-HPLC model LaChrom D-7000 and was monitored by UV absorbance at 225 nm. The 80% Sep-Pak eluate was loaded onto a Sephasil C-18 (250 x 4.1 mm) column (LiChroCART). Peptides were eluted with a linear gradient of acetonitrile (5-60%) with acidified water 0.1% trifluoroacetic acid (TFA) over 90 min at a flow rate of 0.6 ml/min. Fractions were collected every two minutes, lyophilized, reconstituted in UPW and frozen at -20ºC until antimicrobial activity assay. Post-HPLC fractions were analyzed on 18% Tris-tricine/Urea-polyacrylamide gels to confirm the identity of the peptide. Antimicrobial activity of the peptides was monitored by liquid growth inhibition assay (Mitta et al. 1999). In microtiter plates 10 µl of the samples with the peptide were incubated with 100 µl of a suspension of bacteria in TBS [(Tris Borate Saline Buffer) for V. ordalii and V. anguillarum] at a starting optical density of A620: 0.001. Bacterial growth was assayed by measurement of optical density at A620 after a 24 hrs incubation at 26ºC. Percentage growth inhibition was determined by subtracting bacterial growth values in the absence of peptides from that displayed by bacteria exposed to different concentrations of the peptide. As positive control an antibiotic was used to fully inhibit bacterial growth in the absence of added salt. As negative control samples of induced E. coli with plasmid without insert was used. Peptides were solubilized in ultra pure water. The standard CEC A1 geneused in this study is GenBank accession no. X16972. Results A retropeptide-coding sequence of mature Cecropin was designed and synthesized (GenScript) to match the expected polarity for each coding triplets CECret. Considering that a retro triplet-nucleotide sequence does not necessary code for a retro-aminoacid sequence, an adjustment was necessary at the nucleotide level (sequence not shown) in order to generate the expected inverted amino acid coding sequence of the gene (Figure 1). CECdir-CECret and CECret-CECdir were designed and constructed by means of a three-primer driven PCR reaction (Figure 2a and b in order to combine the orientation of the charged amino acids in the doublet molecules. In the case of CECdir-CECret the coding sequence of CECdir was linked in frame with the coding sequence of CECret and the doublet peptide displays the positively charged amino acids to the extremes and the hydrophobic tails oriented towards the interior (Figure 2c). In the case of CECret-CECdir the coding sequence of CECret was linked in frame with the coding sequence of CECdir and the doublet peptide displays the positively charged amino acids to the interior and the hydrophobic tails oriented to the extremes (Figure 2c). The tandem doublet CECdir-CECdir was not constructed based on previous information that it lacked antimicrobial activity (Martemyanov et al. 1997). Figure 3 confirms the construction of the two doublets. An amplification product of the desired size of the doublet coding sequences with their restriction sites (264 bp) is seen in lanes 3-4 and 7-8 for each corresponding doublet obtained when limiting the availability of the hybrid primer (0.1 μM ideal primer ratio). Also shown is the formation of PCR intermediates when there is no limitation in the availability of the hybrid primer (Lanes 2 and 6, respectively). Once confirmed the expected size of CECdir-CECret and CECret-CECdir, the doublets and thecorresponding monomers CECdir and CECret sequences as controls were cloned in the T7 promoter driven expression plasmid pET27b+ where correct frame and sequence were confirmed by sequencing. Expression and purification of doublets Peptide expression was attained only with CECdir-CECret doublet as shown in Figure 4.Samples were analyzed on polyacrylamide gels to allow visualization of peptides from crude cell extract. An expression product of approximately the expected size (8540.9 Da) of the doublet CECdir-CECret was obtained for samples induced with IPTG, but was absent in the samples without IPTG induction. The monomers CECdir and CECret as the doublet CECret-CECdir were not expressed or have been degraded. Additional constructs of monomers and doublets cloned in frame with the pelB periplasmic signal peptide permitted the expression of CECdir, CECret and CECdir-CECret, but showed tenfold reduced activity compared to this construct (data not shown). The total amount of protein recovery from the supernatant of the 500 ml cultures of CECdir-CECret post Sep-Pak fractions was 170 µg present in the 80% ACN fraction. Subsequent data showed that the destination of expression product was primarily towards inclusion bodies. RP-HPLC of the 80% Sep-Pak eluate permitted the obtention of a chromatogram with one major peak showing the hydrophobic property of the peptide evident in the late retention time, where CECdir-CECret was eluted at 47-48% ACN. This peak was analyzed on polyacrylamide gels to allow visualization of purified peptides, confirming the presence of the doublet (Figure 5) showing the same migration of induced cells (Figure 4). Mass spectrometry further confirmed the presence of the recombinant doublet. Post HPLC purified fractions of CECdir-CECret were subject to antimicrobial activity assay performed in triplicates and compared with the monomers CECdir and CECret obtained by chemical synthesis (a kind gift from Dr. Fanny Guzman). The antimicrobial activity of the doublet CECdir-CECret was found in the 80% ACN elution fractions with a retention time of 69.18 min, and showed a two fold enhanced activity compared with its parental counterpart against Gram negative bacteria V. anguillarum and V. ordalii (Table 1).
This study shows a novel design of genetic constructions involving doublets of the AMP Cecropin A from Drosophila melanogaster coding sequences. The strategy involved the inversion of the amino acid coding sequence compared to its original form, with the purpose of enhancing their potential interaction with the membrane structure of bacterial pathogen based upon the synergistic activity displayed by dimeric and/or oligomeric forms of a given AMP. The constructs were restricted to only two doublet forms of CECdir and its retro form CECret resulting in novel configurations CECdir-CECret and CECret-CECdir. We expected that the novel configuration could perhaps mimic eventual dimer formation of cecropin on the bacterial surface that could enhance its interaction or penetrating properties with the microbial membrane, affecting antimicrobial properties. This is the first report of the generation of a doublet AMP without cross-linked disulfides bridges joining the monomers (Hristova et al. 2001; Dempsey et al. 2003) directly obtained by biosynthesis from cells and not by chemical synthesis (Merrifield et al. 1995). These constructs are also different from previous classic doublet construction of cecropin in tandem repeats (CECdir-CECdir), obtained from a cell free transcription-translation system, that was not able to show antimicrobial activity (Martemyanov et al. 1997). The design and construction of the new doublets was not an easy task since ligation of two identical cecropin PCR amplification products could only generate a classic doublet in tandem (CECdir-CECdir). The inversion of the CEC sequence does not give the direct inversion of the amino acid sequence due to genetic code degeneracy. Therefore we had to design and construct a new gene, representing the inverted amino acid sequence which we named CECret. The construction of the “retrogene” provided significant information to sustain the idea of new constructions, since to our surprise, the retropeptide, which has the same amino acids than the standard peptide, was not at all functional, a situation that has a number of interpretations based upon reported analogue cases (Merrifield et al. 1995). The enhanced antimicrobial activity displayed by the doublet cecropin (CECdir-CECret) is an intrinsic property of the new molecule, since CECret did not show any activity by itself and the one displayed by the monomer parental counterpart CECdir, is significantly lower. Enhanced activity by doublets of cecropin could maybe support some process of oligomerization or aggregation of this peptide in the bacterial membrane in a similar way that has been demonstrated with magainin (Dempsey et al. 2003; Zemel et al. 2005). This is one of the few cases in which an active peptide is obtained from bacteria without the use of protein fusions. In addition, this doublet form of cecropin showed enhanced expression and stability compared with the monomeric form. Only obtained associated with a signal sequence such as pelB (Piers et al. 1993; Skosyrev et al. 2003). Thus, this result opens a feasible alternative to commercial-scale synthesis of pure forms of selected antimicrobial peptides using recombinant DNA techniques. The fact that we obtained efficient expression of the doublet CECdir-CECret, but not with the inverted form ofCECret-CECdir could perhaps be due to a required freedom of the hydrophobic tails that could react with the membranes of target microorganisms which in the inverted form are oriented towards the interior and thus not available. In conclusion, these results show the viability in producing recombinant antimicrobial peptides in doublet form, without fusion proteins and without signal peptides. This design could show the way to generate novel doublet AMP with enhanced stability and activity. In this frame, new constructs involving the use of inducible self-cleavage activity of a protein splicing element (termed intein) are currently ongoing in our laboratory. Acknowledgments We thank Paulina Schmitt and Dr. Luis Mercado for their invaluable help and expertise in purification and antimicrobial assays. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08020f4.jpg] [ej08020f5.jpg] [ej08020f2.jpg] [ej08020f1.jpg] [ej08020f3.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}