
|
Electronic Journal of Biotechnology
Universidad Católica de Valparaíso
ISSN: 0717-3458
Vol. 11, Num. 2, 2008
|
Electronic Journal of Biotechnology, Vol. 11, No. 2, April, 2008, pp.
Generation and analysis of an Eucalyptus globulus cDNA library constructed from seedlings subjected to low temperature conditions
Susana Rasmussen-Poblete1 , Jorge Valdes2, Maria Cecilia Gamboa3, Pablo D.T. Valenzuela4 , Erwin Krauskopf*5
1Facultad Ciencias de la Salud,
Universidad Andres Bello,
Republica 252, Santiago, Chile,
Fundación Ciencia para la Vida,
Zañartu 1482, Ñuñoa, Santiago, Chile
Tel: 56 2 2383178
Fax: 56 2 2372259
E-mail: susana.rasmussen@bionova.cl
2Facultad Ciencias de la Salud,
Universidad Andres Bello,
Republica 252, Santiago, Chile,
Center for Bioinformatics and Genome Biology,
Zañartu 1482, Ñuñoa, Santiago, Chile
Tel: 56 2 2383178
Fax: 56 2 2372259
E-mail: jorge.valdes@gmail.com
3Facultad Ciencias de la Salud,
Universidad Andres Bello,
Republica 252, Santiago, Chile,
Fundación Ciencia para la Vida,
Zañartu 1482, Ñuñoa, Santiago, Chile
Tel: 56 2 2383178
Fax: 56 2 2372259
E-mail: cecy_gamboa@yahoo.es
4Facultad Ciencias de la Salud, Universidad Andres Bello,
Republica 252, Santiago, Chile,
Fundación Ciencia para la Vida,
Zañartu 1482, Ñuñoa, Santiago, Chile,
Millenium Institute For Fundamental and Applied Biology,
Zañartu 1482, Ñuñoa, Santiago, Chile
Tel: 56 2 2398969
Fax: 56 2 2372259
E-mail: pvalenzu@bionova.cl
5Facultad Ciencias de la Salud,
Universidad Andres Bello,
Republica 252, Santiago, Chile,
Fundación Ciencia para la Vida,
Zañartu 1482, Ñuñoa, Santiago, Chile,
Millenium Institute For Fundamental and Applied Biology ,
Zañartu 1482, Ñuñoa, Santiago, Chile,
1482, Ñuñoa, Santiago, Chile
Tel: 56 2 2383178
Fax: 56 2 2398969
E-mail: ekrauskopf@unab.cl
*Corresponding
author
Financial support: This work was partially funded by Universidad Andrés Bello (DI Proyect: 04-05/1) and MIFAB (Proyect: P04-071-F) and by the Microsoft Joint Research Program.
Received August 13, 2007 / Accepted December 6, 2007
Code Number: ej08022
Abstract
Eucalyptus globulus is the most important commercial temperate hardwood
in the world because of its wood properties and due to its characteristics for
biofuel production. However, only a very low number of expressed sequence tags (ESTs)
are publicly available for this tree species. We constructed a cDNA from E.
globulus seedlings subjected to low temperature and sequenced 9,913
randomly selected clones, generating 8,737 curated ESTs. The assembly produced
1,062 contigs and 3,879 singletons forming a Eucalyptus unigene set.
Based on BLASTX analysis, 89.3% of the contigs and 88.5% of the singletons had
significant similarity to known genes in the non-redundant database of GenBank.
The Eucalyptus unigene set generated is a valuable public resource that
provides an initial model for genes and regulatory pathways involved in cell
wall biosynthesis at low temperature.
Keywords: cellulose, cold-temperature, EST
database, forest biotechnology, lignin.
Note: The sequences have been deposited in GenBank. Accession numbers:
ES588357-ES597093
Abbreviations: |
4CL: CoA ligase
AUX/IAA: auxin/indole-3-acetic acid
bZIP: basic leucine zip
C3H: p-coumarate 3-hydroxylase
C4H: 4-hydroxylase
CAD: cinnamyl alcohol dehydrogenase
CCR: cinnamoyl CoA reductase
CCoAOMT: caffeoyl-CoA 3-O-methyltransferase
ESTs: expressed sequence tags
F5H: ferulate 5-hydroxylase
GO: gene ontology
HCT: hydroxycinnamoyltransferase
PAL: phenylalanine ammonia lyase |
Forests cover nearly 30% of the earth surface, nearly
4 billion hectares, serving multiple functions including conservation of
biological diversity, renewing the oxygen supply of the atmosphere, preventing
soil erosion and supplying pulp and wood (FAO, 2005). Forest tree breeding aims
to improve the quality of trees by the selection of individuals with desirable
traits that will later be used to produce trees with improved genotype. Genetic
improvement programs such as controlled cross-pollination breeding have been
used since the 1950s. Nevertheless, phenotype assessment is a complex process
due to the long generation times of woody species (Grattapaglia, 2004). It is
within the context of reducing this time-frame that functional genomics has
become a powerful tool in forestry.
In the last few years functional genomics has been
used extensively for gene discovery in species whose genomes have not been
completely sequenced. A cost-effective and rapid way to obtain new data from an
organism is through partial sequencing of randomly selected cDNA clones
(Braütigam et al. 2005). The resulting collection of expressed sequence tags
(ESTs) reveals a portion of genes in an organism expressed under a particular
condition. Using this approach, several traits have been analyzed in trees,
such as wood formation (Allona et al. 1998; Sterky et al. 1998; Israelsson et al. 2003) or cold tolerance (Nanjo et al. 2004; Sterky et al. 2004). Unfortunately,
these studies have focused on gene expression profiles having a direct effect
on the particular trait studied, without expanding the range of effects that
the set condition might have on other metabolic pathways. In fact, cold stress
in poplar cuttings (Populus tremula x Populus tremuloides cv.
Mush1) has been shown to produce variations in parameters such as sucrose
concentration and lignin content, illustrating the direct effect of cold
conditions on wood quality (Hausman et al. 2000).
The amount and type of lignin and cellulose are
important in the timber and pulp industry as they have a direct effect on the
chemical properties of the wood produced by the tree (Jung and Ni, 1998; Fukushima, 2001; Plomion et al. 2001). For the production of biofuels, cellulose
needs to be separated from lignin so it can be made available for enzyme
hydrolysis. Therefore, several research groups have studied different ways by
which to modify lignin and cellulose content on the plant cell wall. As a
result, various studies have shown a co-regulation of these two compounds (Hu
et al. 1999; Li et al. 2003; Rastogi and Dwivedi, 2006). For instance, the
down-regulation of a single lignin biosynthetic gene resulted in a decrease of
lignin production by the plant, while exhibiting an increase in cellulose
production (Hu et al. 1999). Hence, the modification of plant cell wall
composition in trees may provide a way to engineer wood for biofuel production.
E. globulus is considered the most important temperate hardwood plantation species in the world due to its combination of wood properties suitable for the pulp and paper industry (Jones et al. 2002; Grattapaglia, 2004). This tree species has fast growth rates and an ability to adapt to a broad range of geographic locations (ranging from latitude 35ºS to 42ºS), even though its growth rate diminishes due to frost conditions (Jones et al. 2002; Miranda and Pereira, 2002). Most importantly, Eucalyptus has been listed as one of the candidate biomass energy crops by the U.S. Department of Energy (U.S. Department of Energy, 2007). Nevertheless, public genomic information from E. globulus is limited. In fact, an analysis of publicly available E. globulus ESTs at the GenBank EST repository (by July 06, 2007) registered only 3,953 ESTs for E. globulus compared to the mostly represented tree, Pinus taeda (329,469 ESTs). Thus, in this study we provide and describe the first publicly available cDNA library from cold-treated E. globulus seedlings, paying particular attention to genes predicted to be involved in cell wall biosynthesis and the transcription factors suggested to be involved in their regulation).
Materials
and Methods
Plant material
E. globulus seeds were germinated in a soil mixture and grown in
a culture cabinet with a 16 hrs day/8 hs night photoperiod at a temperature of 21ºC. The library was constructed from 3-month old Eucalyptus globulus plants maintained at 4ºC degrees for 30 min. After cold treatment, E. globulus leaves were collected and frozen in
liquid nitrogen until use.
RNA extraction and cDNA library construction
Total RNA was extracted according to the method
described by Chang and collegues (Chang et al. 1993). RNA integrity was
confirmed by gel electrophoresis and 1 mg was quantified using a RNA standard (Invitrogen, Cat 15620-016). Poly (A) mRNA
was isolated from total RNA with the Stratagene Poli (A) Quick mRNA Isolation Kit (Stratagene,
La Jolla, CA, USA). cDNA was prepared and cloned using the vector pExpress
I exploiting the Not I and Eco RV restriction sites. The cDNA library was not
normalized, i.e. no attempt was made to reduce the redundancy of highly
expressed transcripts.
EST sequencing, filtering and assembly
In total, 9,913 bacterial colonies were
randomly picked and single-pass sequence reactions performed. These sequences
were analyzed using Phred base calling software (with Q>20) (Ewing et al.
1998). All traces were subjected to a trimming process for the removal of
ribosomal RNA, poly (A) tails, low-quality sequences, vector and adapter
regions. Sequences with 94% of identity over 40 or more nucleotides were
assembled using the CAP3 software (Huang and Madan, 1999).
Unigene function assignment and
categorization
The unigene set was classified and
analyzed according to gene ontology (GO) terms (Ashburner et al. 2000) across
functional categories. The complete unigene set was compared against the
protein non-redundant database using BLASTX (Altschul et al. 1997) and analyzed
with the InterProScan program (Zdobnov and Apweiler, 2001) to assign a putative
function. GO terms were extracted from the best hits obtained from the BLASTX
comparison against SwissProt-Trembl database (Fleischmann et al. 1999) (E-value
< E-15 and >70% of alignment coverage) and compared to the InterProScan
GO suggestions. All the GO assignments were curated manually (Ashburner et al. 2000).
The unigene dataset was compared to other Eucalyptus cDNA libraries available in Genbank through BlastN program using an e-value
cutoff of E-5.
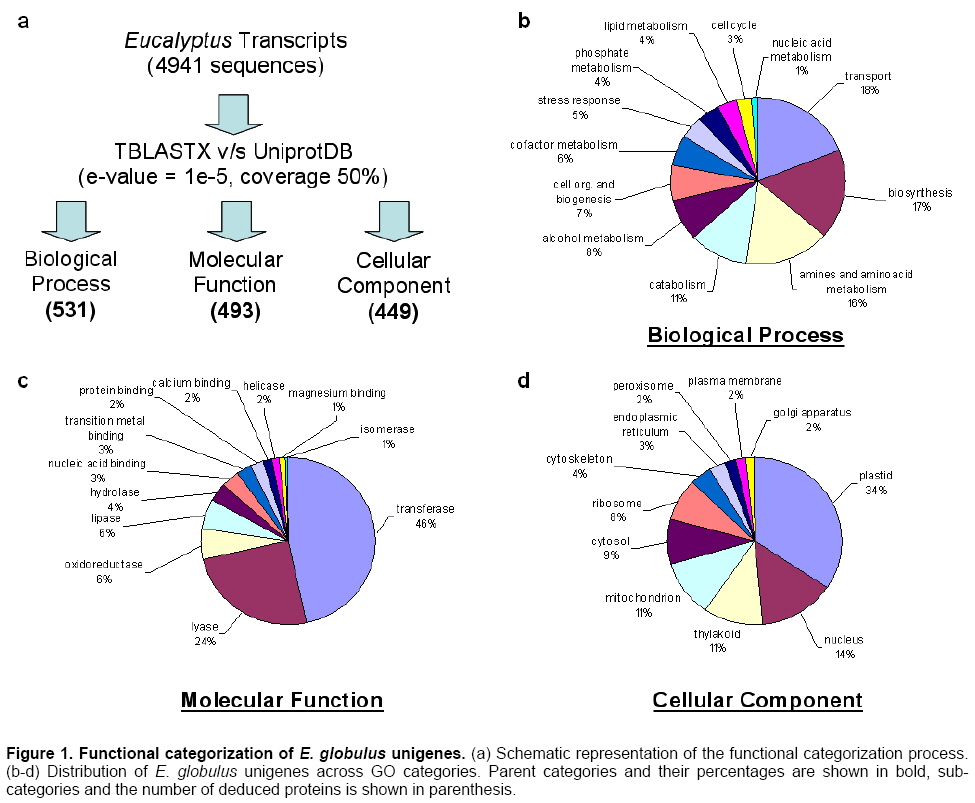
Results and Discussion
Analysis of E. globulus cDNA
library
The analysis of the 9,913 sequence-reads
resulted in the generation of 1,062 contigs and 3,879 singletons (4,941
unigenes) (Figure 1). The fraction of sequences represented by more than
one cDNA was 60.9%, providing an estimate of library redundancy. Based on
BLASTX analysis, 89.3% of the contigs and 88.5% of the singletons had a
significant similarity to known genes in the non-redundant database (Altschul
et al. 1997). As for contigs, their composition ranged from 2 to 118 ESTs. The
deepest contigs were considered highly represented unigenes. Those contigs with
more than 50 ESTs are shown as Table 1 (contigs with 20 or more ESTs are
included as Supplementary data 1).
Table 1. Contigs with ESTs highly represented. Assigned function is indicated in contigs
with more than 50 ESTs. |
Number ESTs |
Contigs length (nt) |
Assigned function |
Relative organism |
Similarity (%) |
Accession number (GI) |
118 |
1604 |
Plastidic aldolase |
Nicotiana paniculata |
94.8 |
4827253 |
86 |
1509 |
Chloroplast latex aldolase-like protein |
Manihot esculenta |
90.9 |
56122688 |
81 |
979 |
Ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit |
Panax ginseng |
93.2 |
77157637 |
80 |
1693 |
Ribulose-1,5-bisphosphate coarboxylase/oxygenase activase precursor |
Malus x domestica |
93.1 |
415852 |
73 |
1590 |
Glyceraldehydes-3-phosphate dehydrogenase A subunit |
Glycine max |
87.8 |
77540210 |
51 |
1721 |
AAA ATPase, central region; homeodomain-like |
Medicago truncatula |
89.1 |
92870561 |
Overall, 541 unigenes were assigned to biological processes, 449 to cellular component and 493 to molecular function categories. This is a low number of assignments compared to other libraries generated in different studies of trees (Pinus: 5474, 5064 and 5886 respectively; Poplar: 6158, 5751 and 6622 respectively; Spruce: 1697, 1467 and 2188 respectively) (Quackenbush et al. 2000). We suggest that this is due to low average similarity between our database and the uniprot sequences database, in addition to the low alignment coverage obtained (we used both parameters to make the assignments). We focused our analyses on the physiological processes (431) being the most represented process related to cellular metabolism, with 48 unigenes related to alcohol metabolism, 95 unigenes associated to amines and aminoacid derivative metabolism, 116 unigenes involved in transport and 164 related to biosynthetic processes.
The most represented molecular functions corresponded
to binding and catalytic activities. The unigenes allocated to binding activity
were associated with ion binding (130) and nucleic acids binding (62). Furthermore,
114 unigenes were associated with enzymes involved in redox reactions related
to lignin biosynthesis and 88 with tranferase activities, including enzymes
involved in lignin and cellulose biosynthesis.
Genes predicted to be involved in wood formation
The EST database was screened for
sequences with significant similarity to genes involved in the biosynthesis of
lignin monomers and cellulose. All of the genes known to participate in the
lignin biosynthetic pathway are represented in our cDNA library. Two of the predicted
gene products, p-coumarate 3-hydroxylase (C3H) and CoA:shikimate/
quinate hydroxycinnamoyltransferase (HCT) had not been described previously
in any Eucalyptus species. However, genes encoding trans-cinnamate
4-hydroxylase (C4H), ferulate 5-hydroxylase (F5H) and 4-coumarate:
CoA ligase (4CL) had been described in other Eucalyptus species but
not in E. globulus (Harakava, 2004). The remainder of the genes found
had been previously described for E. globulus and published in GenBank,
including phenylalanine ammonia lyase (PAL), cinnamoyl CoA reductase (CCR), cinnamyl alcohol dehydrogenase (CAD), caffeic acid
O-methyltransferase (COMT) and caffeoyl-CoA 3-O-methyltransferase (CCoAOMT) (Figure 2) (Supplementary data 2).
The assembly of the C3H and HCT ESTs
showed that two isoforms of their gene-products are represented in our cDNA
library. C3H and HCT
participate in the process of converting p-coumaryl CoA into caffeoyl-CoA,
resulting in the production of coniferyl (G) and sinapyl (S) lignin units. Down-regulation
of C3H in transgenic alfalfa plants and Arabidopsis mutants resulted in
a significant difference in lignin composition due to an alteration in the
number and nature of the monolignol monomers (Franke et al. 2002; Ralph et al.
2006). The characterization of the Arabidopsis reduced epidermal
fluorescence (ref8) mutant defective in C3H suggested that the
genetic modification of this gene may not be appropriate for the reduction of
lignin content in forest species because the mutant plants generated exhibited
vascular collapse, developmental abnormalities and increased susceptibility to
pathogen attack (Boerjan et al. 2003; Cooke et al. 2004).
Three unigenes exhibited similarity to
known cellulose synthase genes. Analysis of their predicted domains by
InterProScan revealed that all of them contained the cellulose synthase domain
that is composed of three aspartic residues and a QXXRW motif, playing a
significant role in the catalytic activity of this enzyme (Krauskopf et al.
2005). However, the zinc finger domains (IPR001841 and IPR011011) present in
cellulose synthase proteins were not found in our sequences since the sequences
were not full-length. The deduced E. globulus proteins were compared
with the ones previously described for E. grandis (Ranik and Myburg,
2006) as no sequences were available for E. globulus (Supplementary
data 3).
Transcription factors involved in wood
formation
Of the 56 transcription factor families
described in Arabidopsis and 63 in rice (Guo et al. 2005; Gao et al. 2006), 11 of them were represented in our library: auxin/indole-3-acetic acid (AUX/IAA)
family, B3 family, basic/helix-loop-helix (bHLH) family, basic leucine zip
(bZIP) family, GRAS family, homeodomain-leucine zipper (HD-Zip) (HB) family, heat
shock family (HSF), MYB family, WRKY family, zinc finger homeobox (ZF-HD)
family and ZIM family. Transcription factors families such as AUX/IAA, MYB and
HD containing domains (zinc finger proteins and homeodomain-leucine zipper)
regulate the expression of genes that participate in xylem development and
secondary wall formation (lignin and cellulose biosynthesis) (Oh et al. 2003; Cánovas
et al. 2004).
Many of the genes encoding the enzymes of
general phenylpropanoid metabolism, such as PAL, C4H, 4CL, COMT and CAD contain
conserved motifs within their promoters that are recognized by plant MYB
transcription factors (Tamagnone et al. 1998). Twelve members of the MYB family
were found in our library. Some of them had a best BLASTX hit with GOLDEN2-like
1 gene, LHY-CCA1-like 5 gene and DIVARICATA gene. The
coverage of the sequences with their best BLASTX hit ranged from 25% and 100%. Two E. gunnii MYB transcription factors sequences were found in GenBank
[GenBank: AJ576023- AJ576024] (Goicoechea et al. 2005). Based on BLASTN analysis,
these sequences were different from the ones obtained in our library. Others
families less represented in our library belonged to the ZF family and bZIP
(with seven members each), WRKY family (five members with coverage of their
best BLASTX hit between 12% and 50%) and one member of the AUX/IAA family, (Supplementary
data 4).
In addition, the data gathered through these analyses
was compared with the few existing Eucalyptus cDNA libraries currently
found in GenBank. The comparison was made against Eucalyptus
gunnii (8,538 ESTs), Eucalyptus
globulus subsp. bicostata (2,685 ESTs), Eucalyptus grandis (1,574
ESTs ) and Eucalyptus globulus ‘blue gum’ (1,266 ESTs). BlastN
comparisons against our E. globulus database revealed a low level of
similarity between our sequenced library and the available datasets. The number
of sequences that have at least one match with E-values better than 1E-5 for
each library were 1,335 ESTs for E. gunnii (15%), 464ESTs
for E. globulus subsp. bicostata (17%), 267 for E. grandis (17%)
and 261 ESTs for E. globulus ‘blue gum’ (17%).
In conclusion, a unigene set of approximately 4900 unigenes was obtained from our E. globulus cDNA library. Analysis of its content has provided valuable data for the future metabolic engineering of plant cell walls by identifying new potential targets that will allow future modification for biofuel production and industrial use. In addition, our results will be useful for comparative genomic studies among hardwoods and softwoods.
Supplementary data 1. Contigs with ESTs highly represented. Assigned function is indicated in contigs with more than 20 ESTs. |
Nº ESTs |
Contigs Length (nt) |
Assigned Function |
Relative Organism |
Similarity (%) |
Accession Number (gi) |
118 |
1604 |
plastidic
aldolase |
Nicotiana
paniculata |
94.8 |
4827253 |
86 |
1509 |
chloroplast
latex aldolase-like protein |
Manihot
esculenta |
90.9 |
56122688 |
81 |
979 |
ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit |
Panax
ginseng |
93.2 |
77157637 |
80 |
1693 |
ribulose-1,5-bisphosphate carboxylase/oxygenase activase precursor |
Malus
x domestica |
93.1 |
415852 |
73 |
1590 |
glyceraldehyde-3-phosphate dehydrogenase A subunit |
Glycine
max |
87.8 |
77540210 |
51 |
1721 |
AAA ATPase, central region; Homeodomain-like |
Medicago
truncatula |
89.1 |
92875540 |
49 |
1320 |
glyceraldehyde-3-phosphate dehydrogenase B subunit |
Glycine
max |
79.5 |
77540212 |
47 |
1051 |
Ferritin-related |
Medicago
truncatula |
86.7 |
92870561 |
45 |
951 |
glycolate oxidase |
Mesembryanthemum
crystallinum |
97.4 |
1773330 |
43 |
1085 |
unnamed protein product type I (CAB-21) (LHCP) |
Nicotiana
tabacum |
96.7 |
19823 |
41 |
940 |
serine-glyoxylate aminotransferase |
Spirodela
polyrrhiza |
84.6 |
74229863 |
39 |
1068 |
thi |
Citrus
sinensis |
85.1 |
2582665 |
36 |
1468 |
putative galactinol synthase |
Pisum
sativum |
83.2 |
541885 |
35 |
916 |
myo-inositol 1-phosphate synthase (MI-1-P) synthase (IPS) |
Sesamum
indicum |
94.0 |
9858816 |
33 |
1346 |
catalase |
Soldanella
alpina |
97.9 |
2462661 |
32 |
1368 |
oxygen evolving enhancer protein 1 precursor |
Bruguiera
gymnorrhiza |
88.8 |
9229957 |
32 |
1261 |
glycine hydroxymethyltransferase methylase |
Solanum
tuberosum |
95.6 |
438247 |
31 |
1286 |
galactinol synthase, isoform GoIS-1 |
Ajuga
reptans |
83.6 |
5608497 |
31 |
961 |
BURP domain-containing protein |
Bruguiera
gymnorrhiza |
59.0 |
14422444 |
28 |
1118 |
ferredoxin--NADP(+) reductase |
Nicotiana
tabacum |
90.3 |
2225993 |
27 |
1761 |
S-adenosylmethionine decarboxylase |
lpomoea
nil |
82.9 |
1498080 |
27 |
2319 |
PAPS-reductase-like-protein |
Catharanthus
roseus |
85.7 |
12831474 |
26 |
1256 |
Phosphoribulokinase, chloroplast precursor |
Mesembryanthemum
crystallinum |
92.7 |
125578 |
25 |
1192 |
AlaT1 |
Vitis
vinifera |
95.4 |
71842524 |
24 |
975 |
polyubiquitin |
Pinus
sylvestris |
96.1 |
1332579 |
23 |
843 |
S-adenosylmethionine decarboxylase |
Prunus
persica |
58.0 |
47232488 |
23 |
1119 |
putative RNA binding protein |
Arabidopsis
thaliana |
84.7 |
3850621 |
23 |
1320 |
translation elongation factor 1A-9 |
Gossypium
hirsutum |
95.4 |
74486744 |
23 |
1214 |
AT4g04640 |
Arabidopsis
thaliana |
85.4 |
7267222 |
22 |
1074 |
OJ000223_09.15 |
Oryza
sativa (japonica cultivar-group) |
91.1 |
38346061 |
21 |
1038 |
aminomethyltransferase system T protein |
Arabidopsis
thaliana |
95.3 |
15221119 |
21 |
1066 |
NADPH-protochlorophyllide oxidoreductase |
Cucumis
sativus |
90.5 |
2244614 |
21 |
1288 |
carbonic anhydrase |
Vigna
radiata |
79.5 |
8954289 |
21 |
956 |
NAD-dependent epimerase/dehydratase family protein-like protein |
Solanum
tuberosum |
93.7 |
82400136 |
20 |
1979 |
P-Protein-like protein precursor |
Arabidopsis
thaliana |
84.5 |
7270248 |
20 |
934 |
S-adenosylmethionine decarboxylase |
Prunus
persica |
86.7 |
47232488 |
20 |
1155 |
sedoheptulose-1,7-bisphosphatase precursor |
Oryza sativa
(indica cultivar-group)] cultivar-group) |
91.8 |
27804772 |
20 |
1238 |
Malate dehydrogenase, glyoxysomal precursor |
Citrullus
lanatus |
83.3 |
126894 |
20 |
1857 |
Transketolase, C-terminal-like |
Medicago
truncatula |
88.4 |
92892666 |
Supplementary
data 2. Analysis of E. globulus unigenes corresponding to enzymes
involved in wood formation. |
Gene Name |
Best Blastx Hit |
Score |
E-value |
Coverage |
Domains |
EgPAL1 |
PAL
Daucus carota |
387 |
8e-113 |
29.09% |
IPR001106
IPR008948
PF00221 |
EgPAL2 |
PAL
Camellia sinensis |
306 |
8e-82 |
29.13% |
IPR001106
IPR008948
PF00221 |
EgPAL3 |
PAL
Citrus limon |
278 |
2e-73 |
25.76% |
IPR001106
IPR008948
PF00221 |
EgC4H |
C4H
Populus kitakamiensis |
482 |
3e-142 |
53.07% |
IPR001128
IPR002401
PF00067 |
Eg4CL1 |
4CL
Populus balsamifera
subsp. trichocarpa x Populus deltoides |
246 |
6e-64 |
33.16% |
IPR000873
PF00501 |
Eg4CL2 |
4CL
Eucalyptus
camaldulensis |
260 |
4e-68
|
36.95% |
IPR000873
PF00501 |
EgHCT1 |
HCT
Nicotiana tabacum |
331 |
4e-89 |
49.66% |
IPR003480
IPR02458 |
EgHCT2 |
HCT
Oryza sativa (japonica
cultivar-group) |
186 |
1e-45 |
45.13% |
IPR003480
IPR02458 |
EgC3H1 |
C3H-1
Ocimum basilicum |
249 |
3e-72 |
29.49% |
IPR001128
IPR002401
PF00067 |
EgC3H2 |
C3H
Ocimum basilicum |
375 |
1e-102 |
44.41% |
IPR001128
IPR002401
PF00067 |
EgCOMT |
COMT
Eucalyptus gunnii |
451 |
1e-125 |
63.66% |
IPR001077
PF00891
IPR011991
IPR012967
PF08100 |
EgCCoAOMT1 |
CCoAOMT
Plantago major |
96.7 |
6e-19 |
47.27% |
IPR002935
PF01596 |
EgCCoAOMT2 |
CCoAOMT
Ammi majus |
121 |
2e-26 |
42.32% |
IPR002935
PF01596 |
EgCCR1 |
CCR
Eucalyptus gunnii |
443 |
1e-123 |
63.00% |
IPR001509
PF01370 |
EgCCR2 |
CCR
Arabidopsis thaliana |
256 |
5e-67 |
61.68% |
IPR001509
PF01370 |
EgF5H1 |
F5H
Camptotheca acuminata |
539 |
1e-151 |
69.26% |
IPR001128
IPR002401
PF00067 |
EgF5H2 |
F5H
Camptotheca acuminata |
126 |
8e-28 |
14.00% |
IPR001128
IPR002401
PF00067 |
EgCAD1 |
CAD
Eucalyptus globulus |
530 |
1e-149 |
88.76% |
IPR002085
IPR002328
IPR011032
IPR013149
IPR013154
PF00107
PF08240 |
EgCAD2 |
CAD
Arabidopsis thaliana |
220 |
4e-56 |
57.91% |
IPR001509 |
Supplementary
data 3. Analysis of E. globulus cellulose synthase unigenes. |
Gene Name |
Best Hit with E.
grandisi |
Best Blastx Hit |
Score |
E-value |
Coverage |
Domains |
EgCesA1 |
EgrCesA5 |
CesA
Populus tremula x
Populus tremuloides |
400 |
1e-110 |
24.11% |
IPR005150
PF03552 |
EgCesA2 |
No EgrCesA related |
CesA
Medicago truncatula |
238 |
2e-62 |
28.34% |
IPR005150
PF03552 |
EgCesA3 |
EgrCesA4 |
CesA4
Eucalyptus grandis |
309 |
9e-83 |
17.4% |
IPR005150
PF03552 |
Supplementary data 4. Analysis of E. globulus unigenes
corresponding to transcription factors. |
Best Blast Hit |
Related Organism |
Score |
E-value |
Coverage |
Domains |
putative MYB transcription factor |
Oryza sativa (japonica
cultivar-group)] cultivar-group)] |
130 |
1e-28 |
30.8% |
IPR001005
IPR006447
IPR009057
IPR012287 |
MYB transcription factor
LHY-CCA1-like5 |
Arabidopsis thaliana |
241 |
5e-62 |
90.1% |
IPR001005
IPR006447
IPR009057
IPR012287 |
GPRI1 (GOLDEN2-LIKE 1);
transcription factor |
Arabidopsis thaliana |
239 |
2e-61 |
77.9% |
IPR000183
IPR001005
IPR006447
IPR009057 |
MYB-like transcription factor
DIVARICATA |
Antirrhinum majus |
328 |
2e-88 |
73.9% |
IPR001005
IPR006447
IPR009057
IPR012287 |
MYB-like transcription factor 6 |
Gossypium raimondii |
288 |
2e-76 |
103.1% |
IPR001005
IPR009057
IPR012287 |
MYB11 |
Malus x domestica |
230 |
4e-59 |
50.3% |
IPR001005
IPR009057
IPR012287 |
MYBR2 |
Malus x domestica |
75.9 |
2e-12 |
41.6% |
No related
InterPro |
MYB-like DNA-binding protein |
Catharanthus roseus |
95.5 |
2e-18 |
24.4% |
No related
InterPro |
transcription factor MYB1 |
Malus xiaojinensis |
88.6 |
2e-16 |
47.0% |
No related
InterPro |
MYB, DNA-binding |
Medicago truncatula |
78.6 |
2e-13 |
28.3% |
No related
InterPro |
MYB transcription factor
LHY-CCA1-like5 |
Arabidopsis thaliana |
147 |
5e-34 |
35.5% |
IPR001005
IPR006447
IPR009057
IPR012287 |
MYB-related protein |
Arabidopsis thaliana |
218 |
2e-55 |
48.8% |
IPR001005
IPR009057
IPR012287 |
IAA18; transcription factor |
Arabidopsis thaliana |
55.5 |
2e-06 |
33.3% |
No related
InterPro |
Transcriptional factor B3 |
Medicago truncatula |
102 |
2e-20 |
32.7% |
IPR003340 |
Transcriptional factor B3 |
Medicago truncatula |
188 |
2e-46 |
22.7% |
No related
InterPro |
bZIP transcription factor protein |
Capsicum annuum |
75.1 |
3e-12 |
45.5% |
IPR004827
IPR008917
IPR011616 |
Putative ripening-related bZIP
protein |
Vitis vinifera |
171 |
3e-41 |
38.7% |
No related
InterPro |
bZIP transcription factor ZIP-2 |
Nicotiana tabacum |
80.5 |
7e-14 |
32.9% |
IPR004827
IPR008917 |
Putative ripening-related bZIP
protein |
Vitis vinifera |
123 |
7e-27 |
37.4% |
No related
InterPro |
ATBZIP60 |
Arabidopsis thaliana |
82 |
2e-14 |
61.7% |
IPR004827
IPR008917
IPR011616 |
Putative ripening-related bZIP
protein |
Vitis vinifera |
176 |
9e-43 |
45.2% |
No related
InterPro |
Putative ripening-related bZIP
protein |
Vitis vinifera |
84.3 |
6e-15 |
16.3% |
No related
InterPro |
GRAS transcription factor |
Medicago truncatula |
67.8 |
5e-10 |
10.2% |
No related
InterPro |
GRAS transcription factor |
Medicago truncatula |
252 |
9e-66 |
33.4% |
IPR005202 |
ATHB-7 |
Arabidopsis thaliana |
142 |
2e-32 |
33.3% |
IPR000047
IPR001356
IPR003106
IPR009057
IPR012287 |
Heat shock transcription factor |
Phaseolus acutifolius |
272 |
1e-71 |
60.7% |
IPR000232
IPR002341
IPR011991 |
WRKY9; transcription factor |
Arabidopsis thaliana |
127 |
1e-27 |
75.4% |
IPR003657 |
Putative WRKY-type DNA binding
protein |
Glycine max |
380 |
1e-104 |
50.5% |
IPR003657
IPR000583 |
Putative WRKY4 transcription
factor |
Vitis aestivalis |
74.3 |
4e-12 |
34.4% |
No related
InterPro |
DNA-binding WRKY |
Medicago truncatula |
168 |
2e-40 |
38.3% |
IPR003657 |
Putative WRKY4 transcription
factor |
Vitis aestivalis |
240 |
5e-62 |
66.6% |
IPR003657 |
Putative zinc finger transcription
factor |
Oryza sativa (japonica
cultivar-group)cultivar-group |
242 |
1e-62 |
35.1% |
IPR000571 |
Putative zinc finger transcription
factor |
Oryza sativa (japonica
cultivar-group)cultivar-group |
243 |
4e-63 |
35.1% |
IPR000571 |
Zinc finger protein |
Malus x domestica |
134 |
3e-30 |
26.1% |
IPR007087 |
Zinc finger protein, putative |
Arabidopsis thaliana |
137 |
1e-30 |
52.4% |
IPR000315 |
putative zinc finger protein |
Oryza sativa (japonica
cultivar-group)cultivar-group |
133 |
9e-30 |
73.5% |
IPR007087 |
Putative zinc finger transcription
factor |
Oryza sativa (japonica
cultivar-group)cultivar-group |
225 |
2e-57 |
31.3% |
IPR000571 |
Zinc finger protein-like |
Arabidopsis thaliana |
123 |
6e-27 |
5.4% |
IPR003349 |
Zinc finger protein, putative |
Plasmodium falciparum 3D7 |
60.1 |
8e-08 |
28.8% |
IPR002653 |
Putative zinc finger transcription
factor |
Oryza sativa (japonica
cultivar-group)cultivar-group |
239 |
9e-62 |
34.8% |
IPR000571 |
Zinc finger protein, putative |
Arabidopsis thaliana |
109 |
2e-22 |
21.6% |
IPR000315
IPR002906 |
Zinc finger protein, putative |
Arabidopsis thaliana |
152 |
1e-35 |
36.3% |
IPR000315 |
Putative zinc finger transcription
factor |
Oryza sativa (japonica
cultivar-group)cultivar-group |
218 |
2e-55 |
28.4% |
IPR000571 |
Acknowledgments
We thank Dr. Danilo González who provided the computer cluster to generate the unigene set and Dr. David Holmes for critical reading of the manuscript.
References
- ALLONA, I.; QUINN, M.; SHOOP, E.;
SWOPE, K.; ST. CYR, S.; CARLIS, J.; RIEDL, J.; RETZEL, E.; CAMPBELL, M.;
SEDEROFF, R. and WHETTEN, R. Analysis of xylem formation in pine by cDNA sequencing. Proceedings
of the National Academy of Sciences of the United States of America, August
1998, vol. 95, no. 16, p. 9693-9698.
- ALTSCHUL, S.; MADDEN, T.; SCHAFFER, A.; ZHANG, J.;
ZHANG, Z.; MILLER, W. and LIPMAN, D. Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids Research,
September 1997, vol. 25, no. 17, p. 3389-3402.
- ASHBURNER, M.; BALL, C.; BLAKE, J.;
BOTSTEIN, D.; BUTLER, H.; CHERRY, J.; DAVIS, A.; DOLINSKI, K.; DWIGHT, S.;
EPPIG, J.; HARRIS, M.; HILL, D.; ISSEL-TARVER, L.; KASARSKIS, A.; LEWIS, S.;
MATESE, J.; RICHARDSON, J.; RINGWALD, M.; RUBIN, G. and SHERLOCK, G. Gene Ontology: tool for the unification of
biology. Nature Genetics, May 2000, vol. 25, p. 25-29. [CrossRef]
- BOERJAN, W.; RALPH, J. and BAUCHER, M. Lignin
biosynthesis. Annual Review of Plant Biology, June 2003, vol. 54,
p. 519-546. [CrossRef]
- BRAÜTIGAM, M.; LINDLÖF, A.; ZAKHRABETKOVA, S.;
GHARTI-CHHETRI, G.; OLSSON, B. and OLSSON, O. Generation and analysis of 9792
EST sequences from cold acclimated oat, Avena sativa. BMC Plant
Biology, September 2005, vol. 5, p. 18. [CrossRef]
- CÁNOVAS, F.; DUMAS-GAUDOT, E.; RECORBET,
E.; JORRIN, J.; MOCK, H.-P. and ROSSIGNOL, M. Plant proteome analysis. Proteomics 2004, vol. 4, p.285-298. [CrossRef]
- CHANG, S.; PURYEAR, J. and CAIRNEY, J. A simple and
efficient method for isolating RNA from pines trees. Plant Molecular Biology
Reporter, June 1993, vol. 11, p. 113-116. [CrossRef]
- COOKE, J.; MORSE, A. and DAVIS, J. Forestry. In: KLEE
and P. CRISTOU eds. Handbook of Plant Biotechnology. United Kingdom. John
Wiley and Sons, 2004, vol. 2, p. 881-904.
- EWING, B.; HILLIER, L.; WENDL, M. and GREEN, P.
Basecalling of automated sequencer traces using phred I. Accuracy Assessment. Genome
Research, March 1998, vol. 8, no. 3, p. 175-185. [CrossRef]
- FAO. Progress towards sustainable forest managment.
In: FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS eds. Global
forest resources assessment. Rome. FAO forestry paper, 2005, p. 147.
- FLEISCHMANN, W.; MOELLER, S.; GATEAU, A. and APWEILER,
R. A novel method for
automatic functional annotation of proteins. Bioinformatics,
March 1999, vol. 15, no. 3, p. 228-233. [CrossRef]
- FRANKE, R.; HUMPHREYS, J.M.; HEMM, M.R.; DENAULT, J.W.;
RUEGGER, M.O.; CUSUMANO, J.C. and CHAPPLE, C. Changes in secondary metabolism
and deposition of an unusual lignin in the ref8 mutant of Arabidopsis. The Plant Journal, April 2002, vol. 30, no. 1, p. 33-45. [CrossRef]
- FUKUSHIMA, K. Regulation of syringyl to
guaiacyl ratio in lignin biosynthesis. Journal of Plant Research,
February 2001, vol. 114, no. 4, p. 499-508. [CrossRef]
- GAO, G.; ZHONG, Y.; GUO, A.; ZHU, Q.; TANG, W.; ZHENG,
W.; GU, X.; WEI, L. and LUO, J. DRTF: a database of rice transcription factors. Bioinformatics, March 2006, vol. 22, no. 10, p. 1286-1287. [CrossRef]
- GOICOECHEA, M.; LACOMBE, E.; LEGAY, S.; MIHALJEVIC, S.;
RECH, P.; JAUNEAU, A.; LAPIERRE, C.; POLLET, B.; VERHAEGEN, D.; CHAUBET-GIGOT,
N. and GRIMA-PETTENATI, J. EgMYB2, a new transcriptional activator from Eucalyptus xylem, regulates secondary cell wall formation and lignin biosynthesis. The
Plant Journal, August 2005, vol. 43, no. 4, p. 553-567. [CrossRef]
- GRATTAPAGLIA, D. Integrating genomics into Eucalyptus breeding. Genetics and Molecular Research, September 2004, vol. 3, no.
3, p. 369-379.
- GUO, A.; HE, K.; LIU, D.; BAI, S.; GU, X.; WEI, L.
and LUO, J. DATF: a database of Arabidopsis transcription factors. Bioinformatics,
February 2005, vol. 21, no. 10, p. 2568-2569.[CrossRef]
- HARAKAVA, R. Genes encoding enzymes of the lignin
biosynthesis pathways in Eucalyptus. Genetics and Molecular Biology,
2004, vol. 28, no. 3, p. 601-607. [CrossRef]
- HAUSMAN, J.F.;
EVERS, D.; THIELLEMENT, H. and JOUVE, L. Compared
responses of poplar cuttings and in vitro raised shoots to short-term chilling
treatments. Plant Cell Reports, October 2000, vol. 19, no. 10, p.
954-960. [CrossRef]
- HU, W.J.; HARDING, S.A.; LUNG, J.; POPKO, J.L.; RALPH,
J.; STOKKE, D.D.; TSAI, C.J. and CHIANG, V.L. Repression of lignin biosynthesis
promotes cellulose accumulation and growth in transgenic trees. Nature
Biotechnology, August 1999, vol. 17, p. 808-812. [CrossRef]
- HUANG, X. and MADAN, A. CAP3 A DNA sequence assembly
program. Genome Research, September 1999, vol. 9, no. 9, p. 868-877.
- ISRAELSSON, M.; ERIKSSON, M.; HERTZBERG, M.; ASPEBORG, H.; NILSSON, H. and MORITZ, T. Changes in gene expression in the
wood-forming tissue of transgenic hybrid aspen with increased secondary growth. Plant Molecular Biology, July 2003, vol. 52, no. 4, p. 893-903. [CrossRef]
- JONES, R.C.; STEANE, D.A.; POTTS, B.M. and
VAILLANCOURT, R.E. Microsatellite and morphological analysis of Eucalyptus
globulus populations. Canadian Journal of Forest Research, January
2002, vol. 32, no. 1, p. 59-66. [CrossRef]
- JUNG, H.-J. and NI, W. Lignification of plant cell
walls. Impact of genetic manipulation. Proceedings of the National Academy
of Sciences of the United States of America, October 1998, vol. 95, no. 22,
p. 12742-12743. [CrossRef]
- KRAUSKOPF, E.; HARRIS, P. and PUTTERILL, J. The
cellulose synthase gene PrCESA10 is involved in cellulose biosynthesis in
developing tracheids of the gymnosperm Pinus radiata. Gene, May
2005, vol. 350, no. 2, p. 107-116. [CrossRef]
- LI, L.; ZHOU, Y.H.; CHENG, X.F.; SUN, J.Y.; MARITA, J.M.;
RALPH, J. and CHIANG V.L. Combinatorial modification of multiple lignin traits
in trees through multigene cotransformation. Proceedings of the National Academy of Sciences of
the United States of America, April 2003, vol. 100,
no. 8, p. 4939-4944. [CrossRef]
- MIRANDA, I. and PEREIRA, H. Variation of pulpwood
quality with provenances and site in Eucalyptus globulus. Annals of
Forest Science, 2002, vol. 59, p. 283-291. [CrossRef]
- NANJO, T.; FUTAMURA, N.;
NISHIGUCHI, M.; IGASAKI, T.; SHINOZAKI, K. and SHINOHARA, K. Characterization of full-length enriched
expressed sequence tags of stress-treated poplar leaves. Plant and Cell
Physiology, 2004, vol. 45, no. 12, p. 1738-1748. [CrossRef]
- OH, S.; PARK, S. and HAN, K.-H. Transcriptional regulation of secondary
growth in Arabidopsis thaliana. Journal of Experimental Botany, December
2003, vol. 54, no. 393, p. 2709-2722. [CrossRef]
- PLOMION, C.; LEPROVOST, G. and STOKES, A.
Wood formation in trees. Plant Physiology, December 2001, vol. 127, p.
1513-1523. [CrossRef]
- QUACKENBUSH, J.; LIANG, F.; HOLT, I.;
PERTEA, G. and UPTON, J. The TIGR Gene Indices: reconstruction and
representation of expressed gene sequences. Nucleic Acids Research,
2000, vol. 28, no. 1, p. 141-145.
- RALPH, J.; AKIYAMA, T.; KIM, H.; LU, F.;
SCHATZ, P.F.; MARITA, J.M.; RALPH, S.A.; SRINIVASA-REDDY, M.S.; CHEN, F. and
DIXON, R.A. Effects of coumarate 3-hydroxylase down-regulation lignin
structure. Journal of Biological Chemistry, March 2006, vol. 281, no.
13, p. 8843-8853. [CrossRef]
- RANIK, M. and MYBURG, A. Six new cellulose synthase genes
from Eucalyptus are associated with primary and secondary cell wall
biosynthesis. Tree Physiology, May 2006, vol. 26, no. 5, p. 545-556.
- RASTOGI, S. and DWIVEDI, UN. Down-regulation of
lignin biosynthesis in transgenic Leucaena leucocephala harboring
O-methyltransferase gene. Biotechnology Progress, 2006, vol. 22, p.
609-616. [CrossRef]
- STERKY, F.; REGAN, S.; KARLSSON, J.; HERTZBERG,M.; ROHDE, A.; HOLMBERG, A.; AMINI, B.; BHALERAO, R.; LARSSON, M.;
VILLAROEL, R.; VAN MONTAGU,M.; SANDBERG, G.; OLSSON, O.; TEERI, T.; BOERJAN, W.; GUSTAFSSON, P., UHLÉN, M.; SUNDBERG, B. and LUNDEBERG, J. Gene discovery in the wood forming
tissues of poplar: analysis of 5,692 expressed sequence tags. Proceedings of
the National Academy of Sciences of the United States of America, October
1998, vol.95, no. 22, p. 13330-13335.
- STERKY, F.; BHALERAO, R.; UNNEBERG, P.; SEGERMAN, B.; NILSSON, P.; BRUNNER, A.; CHARBONNEL-CAMPAA, L.; LINDVALL, J.; TANDRE, K.;STRAUSS, S.; SUNDBERG, B.; GUSTAFSSON, P.; UHLÉN, M.; BHALERAO, R.; NILSSON, O.; SANDBERG, G.; KARLSSON, J.; LUNDEBERG, J. and JANSSON, S. A populus EST resource for plant
functional genomics. Proceedings of the National Academy of Sciences of the
United States of America, September 2004, vol. 101, no. 38, p. 13951-13956. [CrossRef]
- TAMAGNONE, L.; MERIDA, A.; PARR, A.;
MACKAY, S.; CULIANEZ-MACIA, F.; ROBERTS, K. and MARTIN, C. The AmMYB308 and
AmMYB330 transcription factors from Antirrhinum regulate phenylpropanoid and
lignin biosynthesis in transgenic tobacco. Plant Cell, February 1998,
vol. 10, no. 2, p. 135-154.
- U.S. DEPARTMENT OF ENERGY. OIT Agriculture
Plants/crop-based renewable resources 2020. [cited date July 2007], 2007. Available
from Internet: http://www.energy.gov.
- ZDOBNOV, E.M. and APWEILER, R. InterProScan-an
integration platform for the signature-recognition methods in InterPro. Bioinformatics,
2001, vol. 17, no. 9, p. 847-848.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication.
© 2008 by Pontificia Universidad Católica
de Valparaíso -- Chile
The following images related to this document are available:
Photo images
[ej08022f1.jpg]
[ej08022f2.jpg]
|

{kind=link}
{kind=link}