 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 3, July 15, 2008 RESEARCH ARTICLE Development of transformation vectors for the production of potentially high oleate transgenic oil palm Mat Yunus Abdul Masani*1 , Ghulam Kadir Ahmad Parveez2 , 1Advanced Biotechnology and Breeding
Centre,
Biological Research Division,
Malaysian Palm Oil Board,
P.O. Box 10620,
50720 Kuala Lumpur, Malaysia
Tel: 603 89255846
Fax: 603 89259446
E-mail: masani@mpob.gov.my Financial support: In Vitro Transtormation of Oil Palm Programme, Malaysia Palm Oil Board. Received August 20, 2007 / Accepted January 23, 2008 Code Number: ej08032 Abstract The main target of Malaysian Palm Oil Board (MPOB) genetic engineering programme is to produce high oleate transgenic palms. The availability of effective transformation vector is one of the pre-requisites for genetic manipulation of oil palm through recombinant DNA technology. Here, we describe the construction of a series of transformation vectors that have a maize ubiquitin promoter (UbiPro)-driven bar gene for selection of transformants on herbicide (Basta or Bialaphos), and mesocarp-specific promoter (MSP1) for expression of the transgenes [antisense palmitoyl-ACP-thioesterase (PAT) and sense β-ketoacyl-ACP-synthase II (KASII) and sense δ9-stearoyl-ACP-desaturase (SAD)] potentially responsible for high oleate content in oil palm mesocarp. The transformation vectors constructed in this study are suitable for use in both particle bombardment (biolistic) and Agrobacterium-based transformation protocols. Keywords: oleate, transformation vectors, transgenic oil palm.
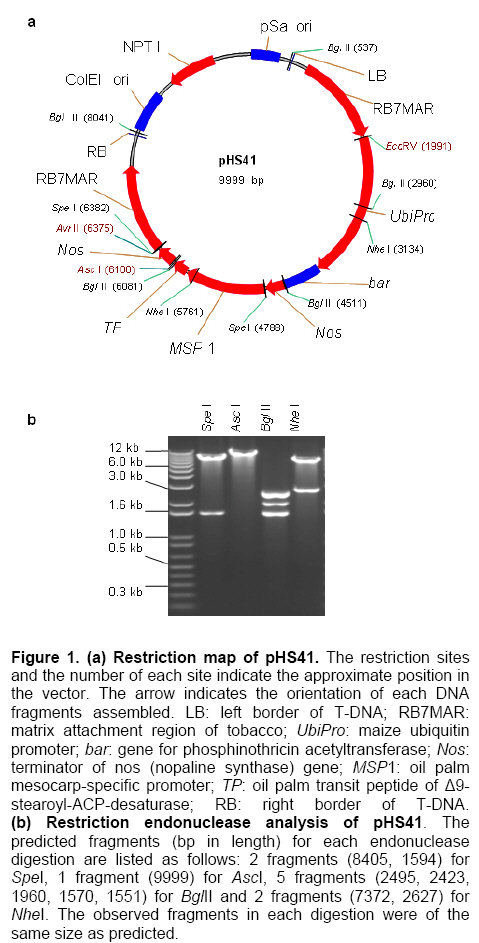
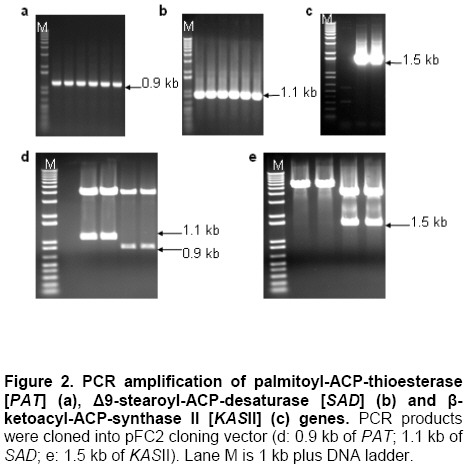
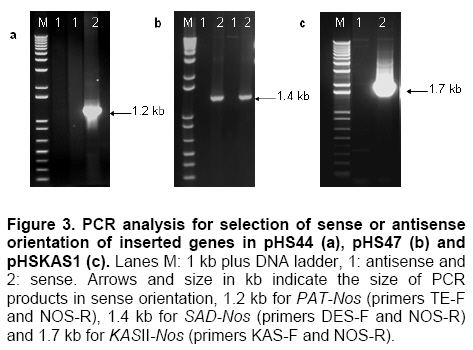
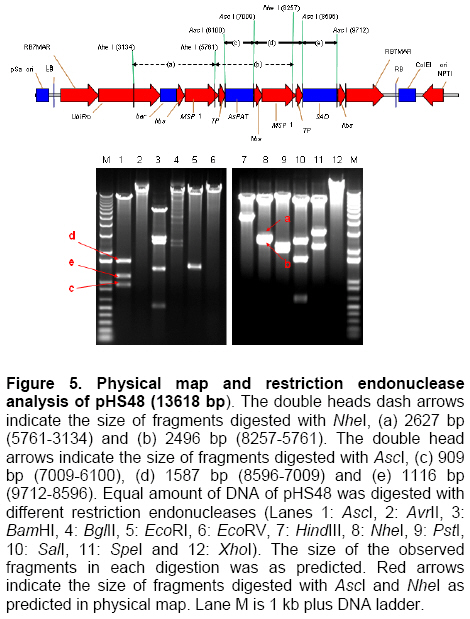
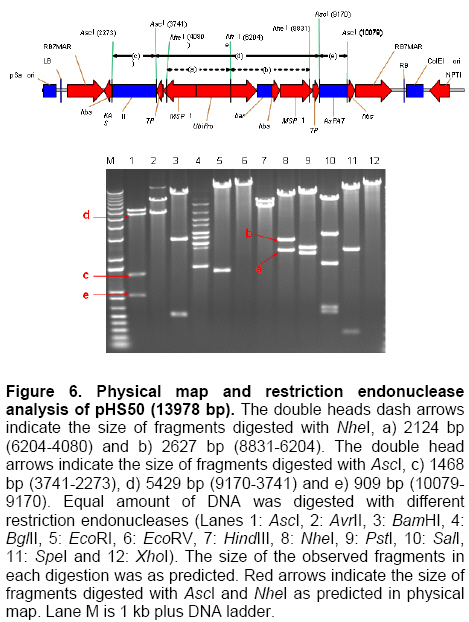
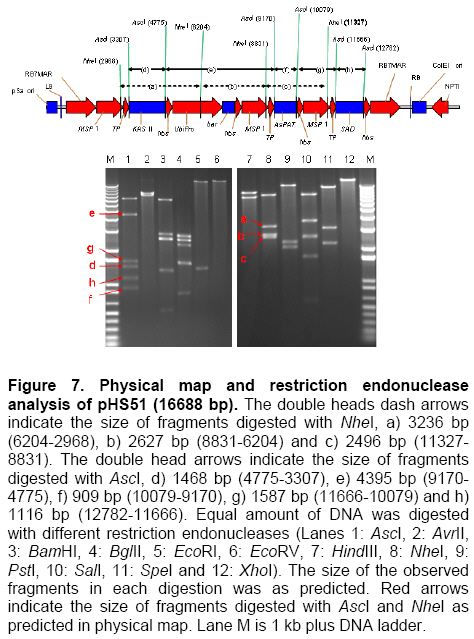
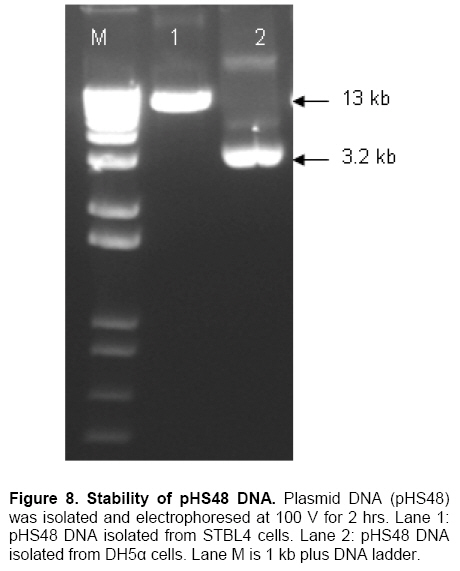
Palm oil is the second largest source of edible oil in the world (Basiron, 2007). The oil palm fruits produce palm oil in the mesocarp and has been admitted as one of the most usable consumer oil compared to other oils derivative plants (Basri et al. 2005). Palm oil has a wide range of applications, about 80% are used for food industries such as cooking oil, shortenings, margarines, ice creams and cookies while the rest are used as feedstock for a number of non-food applications such as diesel fuel substitute, drilling mud, soaps and epoxidised palm oil products, polyols, polyurethanes and polyacrylates (Salmiah, 2000). The relatively high level of palmitate (~44%) provides a degree of stability to the palm oil that makes it suitable for high temperature frying applications (Broun et al. 1999). The approximately 40% oleate and 10% linoleate of palm oil has been shown to lower total serum cholesterol, responsible for the movement of cholesterol within the bloodstream, by lowering the level of the low-density lipoprotein cholesterol (LDL) (Sundram et al. 2003). Furthermore, the high level of the desirable high-density lipoprotein cholesterol (HDL)associated with the removal of cholesterol from the bloodstream, has been proven to protect against heart diseases (Sundram, 1997; Pedersen et al. 2005). Besides the nutritional benefit, the increasing demands from oleochemical industries have make the effort to increase oleate content significant. Based on these factors and the disadvantages of traditional plant breeding techniques, production of high oleate transgenic oil palm through genetic engineering is needed urgently. Increasing oleate of palm oil was considered using the following approaches: increasing β-ketoacyl-ACP-synthase II activity, reducing ACP-thioesterase activity towards palmitoyl-ACP and increasing Δ9-stearoyl-ACP-desaturase activity. These approaches were based on the biochemical and gene expression studies in oil palm mesocarp (Sambanthamurthi et al. 2000). Oil palm Δ9-stearoyl-ACP-desaturase [SAD] (Siti Nor Akmar et al. 1999), palmitoyl-ACP-thioesterase (FatB type thioesterase having predominant specificity towards palmitoyl-ACP and abbreviated here as PAT) (Othman et al. 2000) and β-ketoacyl-ACP-synthase II [KASII] (Ramli et al. 2004) genes have been successfully isolated and characterized. In order to genetically engineer the oil palm fruits for producing high oleate, the three target genes (PAT, SAD, KASII) need to be specifically silenced or expressed in the mesocarp. Isolation of mesocarp-specific promoter (MSP1) has also been successfully achieved (Siti Nor Akmar et al. 2002). Thus, all the genes of interest (GOI) and tissue-specific promoter required to control the enzymes activity of fatty acid biosynthesis in palm oil mesocarp are in hand. Work is continuing to construct the transformation vectors for subsequent transfer into oil palm to produce high oleate transgenic oil palm. DNA work was carried out according to standard protocols (Sambrook et al. 1989). Restriction endonucleases were purchased from Research Biolabs and digestions were performed according to recommendation by the manufacturer. A QIAquick Gel Extraction Kit (QIAGEN) was used to isolate specific restriction and PCR fragments from agarose gels. DNA ligations and subsequent transformations into competent Escherichia coli strain were carried out according to standard protocols (Sambrook et al. 1989). All PCR reactions were performed in a 25 μl volume containing 100 ng plasmid DNA, 10 μM dNTPs mixture, 1.75 units Pfu DNA polymerase (Promega) or Pwo DNA polymerase (Research Biolabs) or Expand High Fidelity PCR System (Roche). Buffer solution contains 100 mM KCl, 100 mM (NH4)2SO4, 200 mM Tris/HCl (pH 8.75), 20 mM MgSO4, 1% Triton X-100, 1000 μg BSA ml-1 supplemented with 1 μM respective primers. The primers were based on the sequence of the target genes and contained convenient cloning site. Amplification reactions were carried out in a MJ Research Inc. Programmable Thermal Controller (PTC-100TM or PTC-200TM). The PCR programme used is: 94ºC for 3 min to denature the DNA template and continued with 25-35 cycles of 1 min at 94ºC (denaturation), 1 min at an appropriate annealing temperature for the primers used to hybridize to the target sequence, and the extension step for polymerization at 72ºC for 1 min. The extension time was roughly set as 1 min for 1 kb DNA. Thus, a longer DNA fragment would require a longer extension time. After the final cycle, the mixtures were further incubated for 5-10 min at 72ºC. The backbone used for all constructs is pMB21, which was modified from the binary Ti vector pGreenII0000 (Hellens et al. 2000a) and contains multiple cloning sites flanked by RB7MAR. The BamHI UbiPro-bar-Nos fragment from pMB16 (Masani et al. 2001) was inserted into BamHI site of pMB21 to create pMB35-46. The SpeI MSP1-Nos fragment was cloned into SpeI site of pMB35-46 to produce pHS41. A unique AscI site in pHS41 was used as cloning site for the insertion of GOI. The GOI, PAT, SAD and KASII were amplified from plasmids, pHA3-1 (Othman et al. 2000), SN19 (Siti Nor Akmar et al. 1999), 2C2 (Ramli et al. 2004) using primer pairs, TE-F (5’-GGCCGGCGCGCCACCCTTGCTGA-3’) and TE-R (5’-GCGCGGCGCGCCTGCCACATCATGC-3’), DES-F (5’-GGCCGGCGCGCCACCGCCTCC-3’) and DES-R (5’-GCGC GGCGCGCCATATTGGTCATGCTC-3’, KAS-F (5’-AAATTTAAGGCGCGCCTGGCGGGCG CCGC-3’) and KAS-R (5’-GCGCGGCGCGCC GCAAACAGGATAACAG-3’), respectively. The PCR products were digested with restriction enzyme AscI and purified by agarose gel electrophoresis. The DNA fragments were later ligated to cloning vector pFC2 (Lonsdale et al. 1995) cleaved by AscI. The resulting plasmids were sequenced to confirm the integrity of the PCR products. The pHS41 is a basic expression vector for the cloning of GOI to be driven by oil palm MSP1 promoter. The salient features and restriction enzyme analysis of pHS41 are given in Figure 1. In order to construct the pHS41 basic expression vector which can overcome the problems in plants transformation applications (Newel, 2000), such as the difficulties of constructing vectors harboring multiple genes and generation of a unique cloning site for insertion of our GOI, the following intermediate plasmids, pMB21 (5636 bp), pMB16 (5727 bp) (Masani et al. 2001) and pMP2 (4554 bp) (Arif et al. 2001) were used. Firstly, to allow for selection of transformed oil palm tissues with the herbicide BastaTM, the bar gene (516 bp), under control of the ubiquitinpromoter (UbiPro; 2010 bp) and terminated by the Nos sequence (Nos; 277 bp), was inserted into pMB21 backbone plasmid. This was carried out by introducing the Ubi-bar-Nos chimeric gene (2766 bp) fragment from pMB16 into the BamHI site in pMB21 and producing a new plasmid designated as pMB35-46 (8405 bp). To confirm the correctness of the insertion, the presence of the bar gene was confirmed by PCR using primers, BAR-F (5’-GCCATGCCGGCGGTCTGC-3’) and BAR-R (5’-TCAGATCTCGGTGACGGGC-3’). The UbiPro-bar-Nos chimeric gene in this plasmid provides selection for herbicide BastaTM (Lohar et al. 2001; Miki and McHugh, 2004). In this study, maize ubiquitin promoter (UbiPro) which has been characterized as suitable promoter for monocot plants was used to drive the bar gene (Ahlandsberg et al. 2001; Rasco-Gaunt et al. 2003). Oil palm transformation studies also revealed that ubiquitin promoter is an efficient and stable promoter in oil palm transformed tissues (Chowdhury et al. 1997; Parveez et al. 1997). In addition, production of transgenics oil palm carrying the bar gene driven by ubiquitin promoter, which are resistant to herbicide BastaTM have been successful obtained (Parveez et al. 2000). In order to control expression of transgenes in oil palm mesocarp, the MSP1 promoter (1140 bp) has been used to drive expression of the GOI. During the period of oil synthesis (15-20 weeks after anthesis), MSP1 promoter is postulated to control the expression pattern of transgenes similar to the fatty acid biosynthetic genes in the mesocarp. This promoter will ensure that most of the effects on lipid metabolism is confined to storage lipids without affecting lipid metabolism in leaves or other tissues which can otherwise lead to deleterious effects in the transgenic plants (Siti Nor Akmar et al. 2002). The plasmid pMP2 was digested with SpeI, and resulting a 1.59 kb DNA fragment carrying the MSP1-TP-AscI-Nos fragment which is then ligated into the SpeI site of plasmid pMB35-46 to produce pHS41 (9999 bp). In oil palm as in other plants, fatty acid synthesis occurs in plastids, and that the newly synthesized fatty acids are exported from plastid to cytoplasm (Sambanthamurthi et al. 2000). Here the fatty acids are utilized in the assembly of triglycerides, which occurs in the endoreticular membranes. In order to target each of the gene of interest into plastid, a 213 bp DNA fragment of transit peptide (TP) encoding an oil palm plastid targeting sequence of Δ9-stearoyl-ACP-desaturase (SAD) gene have been fused at 3’end of MSP1 sequences before AscI cloning site. This was followed by the insertion of Nos sequences, containing the 3-untranslated sequence and polyadenylation signal of the nopaline synthase gene from Agrobacterium tumefaciens, to ensure transcription termination of introduced genes. An AvrII site at 3’-end of Nos sequences was introduced by PCR amplification to allow for ligation compatibility with SpeI site. This cloning site was chosen to insert our GOI without disturbing the internal sites of the sequences. pHS41 have the sequences, MSP1-TP-AscI-Nos and UbiPro-bar-Nos, in the same orientation. This eliminates the possibility of transcriptional read-through into a downstream gene where the opposite orientation could substantially reduce expression of both genes, in transient expression as well stably in transgenic plants (Kusaba, 2004; Tang and Galili, 2004) The matrix attachment region of tobacco (RB7MAR; 1194 bp) was also inserted to flank UbiPro-bar-Nos and MSP1-TP-AscI-Nos at Left-Border (LB) (32 bp) and Right Border (RB) (36 bp) as shown in Figure 1. Matrix attachment regions (MAR) are DNA sequences that bind to the cell’s proteinaceous nuclear matrix to form loop domains (Allen et al. 2000). Transgenes flanked with MAR are thought to be able to form their own chromatic domain and thus be insulated from the influences of factors in the chromatin adjacent to its site of insertion. Because a large majority plant chromatin including oil palm is an inactive conformation at any given time, insulating transgenes with MAR may reduce the incidence of gene silencing as well as enhance transgene expression. It has been shown that when MAR are used to flank both sides of a transgene, the expression level in stably transformed cell lines may become proportional to transgene copy number, indicating that gene activity is independent of its position in the chromosome (Mankin et al. 2003; Halweg et al. 2005). It was reported that flanking a GUS reporter gene with the RB7MAR is resulted in higher number and less variable transgene expression in plant cells (Allen et al. 2000). pHS41 was engineered to allow for transformation into oil palm cells via Agrobacterium-mediated or particle bombardment (biolistic) methods. The backbone of pHS41 vector is pGreenII0000 (3304 bp) (Hellens et al. 2000a; Hellens et al. 2000b) obtained from John Innes Centre, UK. pGreenII 0000 is a mini binary vector that has the advantages of being relatively small (3.3 kb) which facilitates cloning, replicates to high copy numbers in E. coli, and stable in Agrobacterium tumefaciens. pGreenII0000 carries the NPTI resistance gene that confer resistance to antibiotic kanamycin both in E. coli and A. tumefaciens. Its characteristics are suitable for carrying target genes which are larger than 15 kb. This is because it was proposed that the efficiency of in vitro recombination procedures is inversely proportional to the size of the plasmids (Sambrook et al. 1989). Based on the our strategy to produce higher amount of oleate in oil palm mesocarp, KASII and SAD genes will need to be up-regulated while PAT gene will need to be down-regulated. Up-regulating SAD may not be necessary as the enzyme was proven to be very active in mesocarp resulting in most of the stearoyl-ACP being desaturated to oleoyl-ACP (Sambanthamurthi et al. 2000). However, in this study we are testing two sets of constructs, one with SAD and another one without SAD, to ensure that if in the actual situation in transgenic oil palm the SAD is required to effectively pushing the stearoyl-ACP to oleoyl-ACP for accumulating oleic acid, than, we do not have to repeat the whole vector construction and transformation again. However, in the event the SAD was not actually required, than we have transformants from another vector, without SAD, ready for future evaluation and exploitation. The use of these two approaches are important as oil palm transformation is a slow process and to repeat the experiment will be very unproductive and time consuming. PCR amplification was used to amplify the coding regions of PAT gene [917 bp] (Figure 2b), SAD gene [1108 bp] (Figure 2b) and KASII gene [1460 bp] (Figure 2c) with an addition of AscI site at 5’-end and 3’-end of each coding regions. The forward primers (TE-F, DES-F and KAS-F) were designed to remove the start codon of each coding regions to ensure that the translation initiates from the start codon of the transit peptide. The PCR fragments were digested with AscI and cloned into same site of pFC2 (Lonsdale et al. 1995) to create plasmid pTE, pDES (Figure 2d) and pKAS1 (Figure 2e), respectively. A 917 bp AscI fragment from pTE plasmid was then cloned in an antisense orientation next to the transit peptide region of pHS41 plasmid to create pHS44 (10908 bp). A 1.1 kb AscI fragment of pDES was inserted in sense orientation downstream from the transit peptide sequence in pHS41 to generate pHS47 (11115 bp). The KASII coding sequence, isolated as a 1.5 kb AscI fragment from pKAS1, was inserted in sense orientation between the transit peptide and Nos sequence of pHS41 to give pHSKAS1 (11467 bp). The antisense or sense orientation of PAT, SAD and KASII genes in pHS44, pHS47 and pHSKAS1, were confirmed by PCR using forward primers, TE-F, DES-F, KAS-F and reverse primer, NOS-R, respectively (Masani et al. 2001). As shown in Figure 3, in sense orientation, PCR products of approximately 1.2 kb for PAT-Nos, 1.4 kb for SAD-Nos, 1.7 kb for KASII-Nos were amplified, whereas in antisense orientation, no PCR products were amplified. Digestions with SpeI, AscI, BglII and NheI (Figure 4) confirmed the orientation and size of DNA fragments in pHS44, pHS47 and pHSKAS1 single-gene transformation vectors. In addition, considering the issues related to presence of antibiotics selection gene or the insertion of undesired DNA fragment from the transformation vector backbone, the SpeI DNA fragments of pHS44, pHS47 and pHSKAS1 can be used to digest and transform oil palm with minimal DNA fragments for clean-gene technology approach (Xiangdong et al. 2000). This approach will eliminate the use of circular vector containing antibiotic resistant and other genes in the backbone vector. It also seems that transformation using DNA fragments will be the easiest system to introduce desired multiple-gene without further manipulation to combine the genes in a single vector or crossing the transgenic plants carrying different gene. A number of approaches have been used to introduce multiple-genes into plant genomes and followed by coordinated transgene expression (Abbott et al. 2002). These approaches include sexual crossing between plants carrying different transgenes and co-transformation with multiple vectors. Each approach has its specific limitations. For example, genetic crossing between transgenic plants is very time-consuming and requires the use of different selectable marker genes whereas the efficiency of co-transformation with multiple vectors decreases progressively with increasing vector number. Thus, the copy numbers and the relative arrangement among transgenes cannot be controlled. In addition, multiple vectors co-transferred by biolistics are often integrated into a few chromosome loci at high copy number, which is not favourable for expression of transgenes (Chen et al. 1998). Considering the above limitations, transformation of multiple-genes in a single vector is a conventional and reliable approach for increasing oleate in oil palm as it will introduce four genes (bar, PAT, SAD and KASII). This was accomplished by constructing the following pHS48, pHS50 and pHS51 multiple-genes transformation vector as illustrated in Figure 5, Figure 6 and Figure 7, respectively. pHS48 (13618 bp) contains antisense PAT and sense SAD genes was constructed by cloning the SpeI fragment from pHS47 containing a MSP1-TP-SAD-Nos (2.6 kb) into the AvrII site of pHS44. The MSP1-TP-KASII-Nos expression region, isolated as a 3.0 kb SpeI fragment from pHSKAS1, was inserted into the EcoRV site of pHS44 by blunt-end ligation to generate pHS50 (13978 bp) containing antisense PAT and sense KASII genes. As the main objective is to assemble the antisense PAT and sense SAD genes with KASII gene, the rendered blunt MSP1-TP-KASII-Nos fragment from pHSKAS1 was later been cloned into the EcoRV site of pHS48 to produce pHS51 (16688 bp). One of the problems associated with the effort to clone multiple-gene in a single vector is maintaining the repeated sequences such as promoter, transit peptide and Nos terminator sequences. The repeated sequences are unstable in most commonly used strains of E. coli (Schmidt and Bloom, 1999). In this study, for the construction of pHS48, pHS50 and pHS51, the problem was avoided by transforming the ligation vectors into E. coli STBL4 (Life Technologies) rather than conventional E. coli DH5α. E. coli STBL4 has been shown to maintain plasmid containing over 100 copies of 32 bp repeats sequences (Schmidt and Bloom, 1999). As shown in Figure 8, pHS48 transformed into E. coli STBL4 was able to retain the expected size of single circular-form at 13 kb compared to pHS48 transformed into E. coli DH5α. In E. coli DH5α, the pHS48 plasmid was reduced to an approximately 3.2 kb and appear as multimeric circular-forms due to deletions and probably rearrangement. The multiple-gene transformation vectors, pHS48, pHS50 and pHS51, were further analysed using restriction enzymes digestion as shown in Figure 5, Figure 6 and Figure 7, respectively. The vectors were digested to the expected number of fragments and sizes as in the right orientation. Digestion with AscI showed that all inserted genes, 0.9 kb PAT, 1.1 kb SAD and 1.4 kb KASII, were fragmented to the expected sizes in the vectors. The repeating sequences of MSP1 promoter, two in pHS48 and pHS50 and three in pHS51, were demonstrated to be stably maintained without deletion or rearrangement when digested with NheI (Figure 5, Figure 6 and Figure 7). Six transformation vectors were successfully constructed and bombarded into oil palm embryogenic cultures. The transformation vectors contain different genes and different combinations are expected to increase oleate in oil palm mesocarp. Additionally, an efficient multiple-gene assembly vector strategy that overcomes the drawbacks of existing cloning methods and allows the performing of many rounds of gene recombination was developed in this study. The designed strategy for construction of multiple-gene vectors is flexible and versatile, and the placement and orientation of inserted genes can be simply designed and cloned in a reliable step-by-step process. Therefore, the strategy provides a powerful tool for manipulating multiple-gene for the genetic improvement of oil palm and other plants. The authors thank the Director-General of MPOB for permission to publish this paper. We would also like to acknowledge Datin Prof. Madya Dr. Siti Nor Akmar, Dr. Mohd Arif Manaf, Dr. Umi Salamah Ramli and Dr. Abrizah Othman for the mesocarp-specific promoter and genes. Thanks are also due to Fatimah Tahir, Norlinda Yazid, Siti Marlia Silong, Mohd Ali Abu Hanafiah and other Transformation Group members for their technical assistances.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08032f2.jpg] [ej08032f1.jpg] [ej08032f8.jpg] [ej08032f7.jpg] [ej08032f6.jpg] [ej08032f3.jpg] [ej08032f4.jpg] [ej08032f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}