 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 3, July 15, 2008 RESEARCH ARTICLE PCR-based sensitive detection of the edible fungus Boletus edulis from rDNA ITS sequences Bin Lian*1 , Jin-ping Zang2, Wei-guo Hou3, Sheng Yuan4, Donald L. Smith5
1State Key Laboratory of
Environmental Geochemistry,
Institute of Geochemistry,
Chinese Academy of Sciences,Guiyang, China
Tel: 86 851 5895148
Fax: 86 851 5895148
E-mail: bin2368@vip.163.com Financial support: The National Science Fund for Innovative Research Group (No. 40721002); Attracting Talents of Nanjing Normal University (No. 184070H2B39); Ministry of Science and Technology of China (No. 2006CB403200). Received October 8, 2007 / Accepted January 25, 2008 Code Number: ej08040 Abstract Identification of commercially important fungi, such as the valuable edible fungus Boletus edulis can be difficult considering visual or metabolic approaches. Based on phylogenetic analysis of the rDNA ITS sequence, a pair of specific primers was designed for differentiating B. edulis from other mushrooms by PCR. PCR was performed with total DNA as a template at an annealing temperature between 56-60ºC. Positive amplicons were obtained from B. edulis with all DNA templates from fruit bodies and cultured mycelium, but not from other fungal species at an annealing temperature of 60ºC. The result indicated that B. edulis could be clearly distinguished from other fungi by PCR, and there were no misidentifications under the reaction conditions used. The primers were also successfully employed to identify various tissues of B. edulis. Keywords: Boletus edulis, detection, edible fungi, internal transcribed spacer, PCR, specific primers.
Boletus edulis is a valuable edible mushroom, and also a medicinal fungus that has long been used as a Chinese traditional medicine in the treatment of various gastric digestive diseases. Because of its delicious taste, rich nutrient value, anti-tumour, immunomodulatory and lipid-lowering effects, as well as other possible medical values (Manzi et al. 2001), B. edulis has attracted much attention, resulting in an increasing number of studies on its physiological, biochemical and pharmacological properties all over the world (Tang and Lu, 1999; Manzi et al. 2001; Davoli and Weber, 2002; Wilburn et al. 2004; Wang and Lu, 2005; Tsai et al. 2007). In addition, ectomycorrhizal fungi, including B. edulis, play an important role by not only promoting the growth of the plants, but also mobilizing mineral nutrients from rhizosphere soils (van Hees et al. 2004; Kahle et al. 2005; Fomina et al. 2006; Wilson et al. 2007). Accurate taxonomic identification and phylogenetic classification of B. edulis would be helpful in characterizing the novel genes and useful metabolites that could be provided by this fungus, and could help in delivering useful information for future genetic engineering or the cultivation and commercialization of this important species (Moor et al. 2002; Iotti et al. 2005; Agueda et al. 2006; Mello et al. 2006). On the other hand, the evolutionary history and relationships of B. edulis with other basidiomycetes remains controversial in spite of its economic importance (Hibbett and Donoghue, 2001; Leonardi et al. 2005; Peintner et al. 2007). To solve such problems, correct identification of precious fungi is necessary. Unfortunately, traditional methods for identifying mushrooms are not reliable and are also problematic because most fungi in the mycelial phase are difficult to distinguish from each other using either morphological characteristics or organic/inorganic components. Therefore, an advanced method needs to be developed for the identification of B. edulis and other mushroom. With current advances in biotechnology, molecular genetic markers have been employed for rapid identification of different kinds of mushrooms (Lee et al. 2006; Moreau et al. 2006; Froslev et al. 2007; Urbanelli et al. 2007). The development of DNA-based PCR and taxon specific primers (Mullis and Faloona, 1987) has made the detection and study of fungi increasingly feasible. The internal transcribed spacer (ITS) region has generally been considered a convenient target for the molecular identification of fungi at species level (Sanchez-Ballesteros et al. 2000). Henry et al. (2000) identified the fungus Aspergillus at species level and differentiated it from other true pathogenic and opportunistic molds using ITS 1 and ITS 2, allowing for earlier diagnosis and screening of effective antifungal agents for patients. Oh et al. (2003) developed an effective detection method for wood-decaying fungi by hybridization of immobilized sequence-specific oligonucleotide probes with fluorescent-labeled PCR-amplified fungal rDNA ITS sequences. In this study, a preliminary phylogenetic analysis of ITS sequences was performed, and the results were used to assess the phylogenetic relationship between Boletus and other species. Furthermore, a set of PCR primers specific for B. edulis was designed for establishing a convenient and accurate method of identifying this edible mushroom. Materials and Methods B. edulis fruit bodies (MH1) were collected from Kunming, Yunnan Province, China and were identified according to a previously published guide (Wei, 1979). A small tissue from the fresh section between the pileus and stipe was cut with a sterilized knife and placed into a slant of Glucose-Peptone-Agar medium (Glucose 20 g, peptone 2 g, K2HPO4 1 g, KH2PO4 0.5 g, MgSO4 0.5 g, VB1 0.05 mg, Agar 18 g, and H2O 1000 mL). The tissue was incubated for 8 weeks at 26ºC, until pure mycelium covered the agar surface in the slant tube. Table 1 lists 7 samples of 6 species used in this study. The specimens were placed in the collection of Nanjing Normal University, China. The B. edulis isolate and other fungal species were fostered in 100 mL sterilized liquid Glucose-Peptone-Agar (agar free) medium, 26ºC and 150 rpm. Visualized spherical mycelia of B. edulis can be obtained about 7 weeks later, and those of other species, which are growing much faster, can be obtained within a week.
Total genomic DNA was extracted as described by Graham et al. (1994). About 100 mg fresh mushroom tissue or fungal mycelia (Table 1) were ground in 0.8 mL lysis buffer (200 mM Tris-HCl, pH 8.0, 100 mM NaCl, 25 mM EDTA, and 0.5% Sodium dodecyl sulfate) and transferred into a 1.5 mL microfuge tube. After incubation at 65ºC for 1 hr, 0.7 mL 5% CTAB buffer (5% CTAB [w/v], 100 mM Tris-HCl pH 8.0, 1.4 M NaCl) was added, and the mixture was placed in a water bath at 65ºC for 10 min. The solution was extracted with an equal volume of a mixture of phenol:chloroform:isoamyl alcohol (25:24:1, v/v), and then centrifuged at 12,000 rpm for 10 min, followed by extraction with chloroform: isoamyl alcohol (24:1, v/v) until the upper phase was clear. The upper phase was then transferred to a new tube and DNA was precipitated with an equal volume of isopropanol. The precipitate was washed with 70% ethanol and pure ethanol, dried, and re-suspended in 50 μL sterile water. Genomic DNA was visualized in a Gel Documentation System LG 2020 (Hangzhou Langqi, Inco., China). PCR amplification and sequencing of the rDNA ITS region Amplification and sequencing of B. edulis was performed using a pair of universal primers: ITS5 (5’~GGAAGTAAAAGTCGTAACAAGG~3’) and ITS4 (5’~TCCTCCGCTTA TTGATATGC~3’) (White et al. 1990) for the region containing ITS1 and ITS2 and the 5.8S rDNA, and they were also employed as a positive control in subsequent diagnostic PCR. The amplification was carried out in a PTC-100 thermocycler in a 50 μL reaction mixture containing 5 μL 10 x PCR buffer, 4 μL 25 mM MgCl2, 1 μL dNTP Mix (10 mM each), 1 μL 20 μM each primer, 4 μL template DNA, and 0.25 μL Taq DNA polymerase (5U/μL). The reaction mixtures were denatured at 94ºC for 5 min and subjected to 30 cycles of 1 min at 94ºC, 1 min at 56-60ºC, 1 min at 72ºC, and a final extension step of 10 min at 72ºC. Amplified products were purified using a DNA purification kit (Shanghai Watson Bioengineering, Inc., China) according to the manufacturer’s instructions, and then sequenced directly with BigDyeTM on an ABI-PRISM 310 automated DNA sequencer (PE Applied Biosystems Inco., USA). Sequence data from ribosomal ITS genes of Homobasidiomyetes were used to perform a preliminary phylogenetic analysis. 40 ITS sequences were obtained from Genbank-database (htpp://www.ncbi.nlm.nih.gov/) (Table 2). These species designations were based on morphological species concepts or referred to previous reports (Hibbett et al. 1997; Hibbett and Binder, 2002). Phylogenetic analysis of the aligned sequences was performed by Neighbor-Joining (NJ) analysis, using the MEGA (2.0) Kimura2-Parameter Distance model, in which gaps were treated as missing data (Jeanmougin et al. 1998). Bootstrapping was performed with NJ analysis using 1000 replicates.
Based on the results of the phylogenetic analyses, we investigated ITS sequences of the Boleteae family and of other species (Table 3). A pair of primers, BE1 and BE2, was designed for PCR detection of authentic species of B. edulis. The closely related rDNA ITS sequences were retrieved from the DNA database using the BLAST program (htpp://www.ncbi.nlm.nih.gov/blast/) to verify the specificity of the primer pairs. The primer pairs were tested for sensitivity with total genomic DNA from 7 samples of fungi (Table 1) in PCR and a DNA fragment (approximately 300 bp) from the rDNA ITS gene was amplified. Diagnostic PCR was performed under the conditions mentioned above.
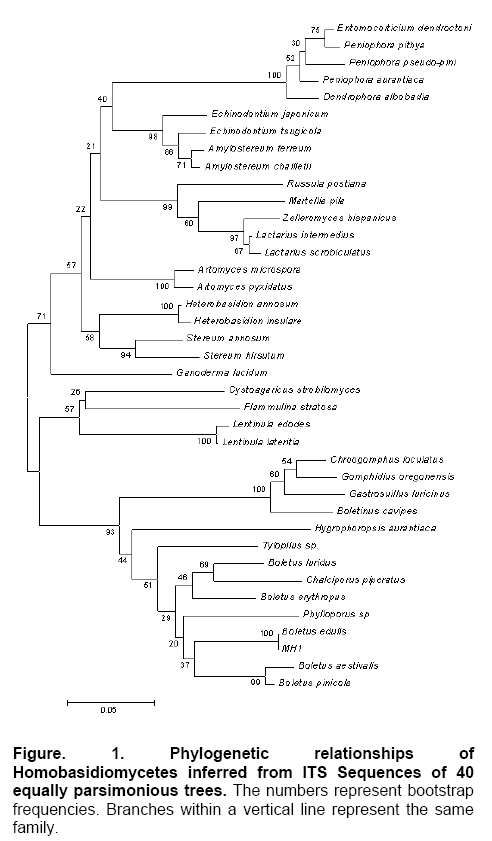
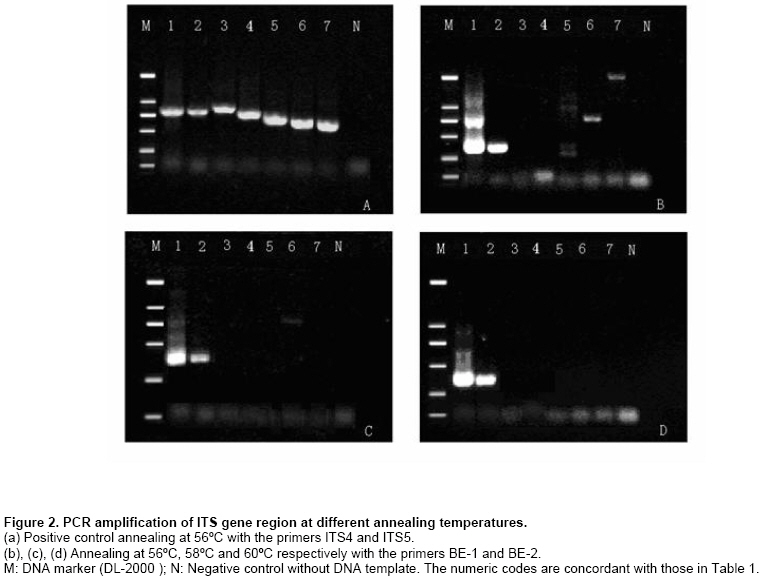
Results Sequencing and phylogenetic analysis A 698 bp fragment of the r-DNA ITS region was amplified and sequenced, the results were submitted to gene bank databases (Accession number: DQ352861). The phylogenetic tree was generated from 40 aligned sequences (Table 2) of Homobasidiomycetes (Figure 1). The genus of Boletus lineage could be identified from the tree, in spite of a low frequency value and other species such as Phylloporus sp. and Chalciporus piperatus appearing within the Boletus lineage (indicated in the bottom of the tree). We comprehend the situation for that the Boletus, Phylloporus and Chalciporus genera all belong to the family Boletaceae and maybe share a very close phylogenetic distance. However, it is also shown that there are great genetic variations within the Boletus genus according to phylogenetic distances based on ITS sequences, and it was feasible to design a pair of specific primers within B. edulis ITS sequence to identify the fungal species. Species specific PCR primers design The primers were designed on the basis of phylogenetic analysis, and then six fungal species were chosen to test them. The selected ITS amplicon for specific primers ranged from bp 4 ~ 26, and from bp 334 ~ 354, respectively. The sequences of the specific primer pairs were BE1 (5’~CATTATCGAGTTAGACCGGGAAG~3’) and BE2 (5’~CCATGCCCTCGAGATCAGA TC~3’). The primer-blast results showed that partial sequences of the ITS of the isolates B. edulis (from China and Switzerland), and B. aereus and B. aestivalis (from Italy) shared 100% identity with the primer BE1; only the sequence of B. edulis from Yunnan, China shared 100% identity with the primer BE2 and an isolate of B. aestivalis from Italy shared 95% (20/21) identity with BE2. However, the false results may be eliminated by a higher annealing temperature during diagnostic PCR. All the samples gave amplicons with sizes between 600 bp and 700 bp with the primer pairs of ITS5 and ITS4 (Figure 2a). With the specific primers BE1 and BE2 and a chosen annealing temperature of 56ºC, a DNA fragment with a size of about 300 bp was clearly amplified from total DNA from the B. edulis isolate as well as from tissues of B. edulis fruiting body, but a weak signal was obtained in the case of G. sinense, A. oryzae and A. nidulans (Figure 2b). When the annealing temperature was increased to 58ºC, we were able to eliminate the negative signals of G. sinsense and A. nidulans other than A. oryzae (Figure 2c). Once the annealing temperature was set to 60ºC, the amplified DNA band only appeared for B. edulis, and no PCR product was obtained for other fungi (Figure 2d). Discussion With increasing demand for edible and medicinal fungal materials, the shortage of Chinese edible and medicinal wild fungi, such as B. edulis, G. lucidum, Tricboloma mutsutake and Cordyceps sinensis, becomes more severe. Consequently, adulterants or substitutes for those precious fungi begin to appear in the market of some areas, being sold fraudulently for high profit. Identification of those high-quality fungal species is not only necessary but has great economic significance as it will allow product distributors to verify the material they are selling. Molecular technology can greatly enhance detection sensitivity, as well as simplify and expedite the identification of fungi. Several methods, including random amplified polymorphic DNA (RAPD), arbitrary primed PCR (AR-PCR), restriction fragment length polymorphism (RFLP), PCR-RFLP and DNA sequencing, have recently been used for the authentication of biological materials (Lotufo et al. 1994; Lakra et al. 2007; Taylor and Ford, 2007). Although they have been proven to be efficient in taxonomic identification and in distinguishing genuine crude drugs from their substitutes or adulterants in previous reports, the application of these methods is limited by the high cost of the fine quality template DNA that is required in these experiments. The reproducibility of RAPD analysis is heavily affected by the quality and concentration of the template DNA, the ratio of template to primer, and slight fluctuations of reacting components or cycling parameters. In regards to the PCR-RFLP method, the length of PCR products also confines its utilization, as the number of restriction enzyme sites is limited in DNA segments between two primers. Although sequence analysis of PCR products is quite precise and stable, the relatively high expense of DNA sequencing and the sensitivity of contamination in the PCR reaction using universal primers obstructs its wide acceptance in quality control of medicinal materials (Wang et al. 2000). This study presents an efficient method for identifying economically important fungi on species level. Based on the target DNA sequence analysis of those high-value species of medicinal mushrooms, we designed a pair of specific primers that exactly match a specific DNA sequence of B. edulis, and incompletely match with the sequence of other species. Therefore, a high-stringency PCR reaction with primers specific for B. edulis gave a positive signal only for genuine species of B. edulis, but not for other fungi. Furthermore, the process of identification by PCR is very simple and convenient to use. We choose B. edulis for the design of detection PCR primers because it is abundant in China. To probe into the evolutionary history of the genus Boletus and its relationship with other basidiomycetes, and to prepare for the specific primer design, we established a phylogenetic tree for Boletus and some strains related at the order and family levels in the Homobasidiomycetes. Based on this phylogenetic tree, the ITS sequences from Boleteae family and other species were aligned, and the results were used to design a pair of specific primers (BE-1 and BE-2) only for Boletus edulis. In conclusion, an accurate and practical phylogenetic analysis establishes a theoretical foundation for defining a classified status of new edible or medicinal fungi. In addition, their evolutionary relationships could provide an important clue for further exploration of the active compounds. Furthermore, specific detection PCR is a time- and cost-effective method for the authentication of edible and medicinal mushrooms. This method will aid in the quality control of identification of other economical materials and species, especially in the inspection of Chinese B. edulis export in big batch. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08040f1.jpg] [ej08040f2.jpg] |
| |||||||||

{kind=link}
{kind=link}