 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 4, October 15, 2008 RESEARCH ARTICLE Cloning and characterization of resistance gene analogs from under-exploited plant species Ramasubramanian Thirumalaiandi1 , Michael Gomez Selvaraj2 , Raghu Rajasekaran3 , Mohankumar Subbarayalu*#4
1Division of Crop Protection,
Central Research Institute for Jute and
Allied Fibres,
Indian Council of Agricultural Research,
Barrackpore, Kolkata, 700 120, India
Tel: 91 33 2535 6121/6122. Ext. 220
Fax: 91 33 25350415
E-mail: tramasubbu@rediffmail.com Received March 4, 2008 / Accepted August 20, 2008 Code Number: ej08045 Abstract Genomic DNA sequences sharing homology with NBS region of resistance gene analogs were isolated and characterized from Pongamia glabra, Adenanthera pavonina, Clitoria ternatea and Solanum trilobatum using PCR based approach with primers designed from conserved regions of NBS domain. The presence of consensus motifs viz., kinase 1a, kinase 2, kinase 3a and hydrophobic domain provided evidence that the cloned sequences may belong to the NBS-LRR gene family. Conservation of tryptophan as the last residue of kinase-2 motif further confirms their position in non-TIR NBS-LRR family of resistance genes. The Resistance Gene Analogs (RGAs) cloned from P. glabra, A. pavonina, C. ternatea and S. trilobatum clustered together with well- characterized non-TIR-NBS-LRR genes leaving the TIR-NBS-LRR genes as a separate cluster in the average distance tree constructed based on BLOSUM62. All the four RGAs had high level of identity with NBS-LRR family of RGAs deposited in the GenBank. The extent of identity between the sequences at NBS region varied from 29% (P. glabra and S. trilobatum) to 78% (A. pavonina and C. ternatea), which indicates the diversity among the RGAs. Keywords: Adenanthera pavonina, Clitoria ternatea, NBS-LRR, Pongamia glabra, resistance gene analogs; solanum trilobatum.
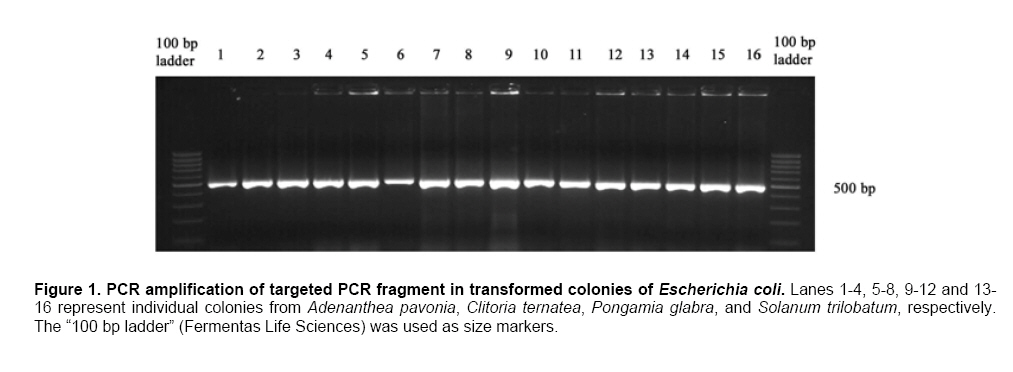
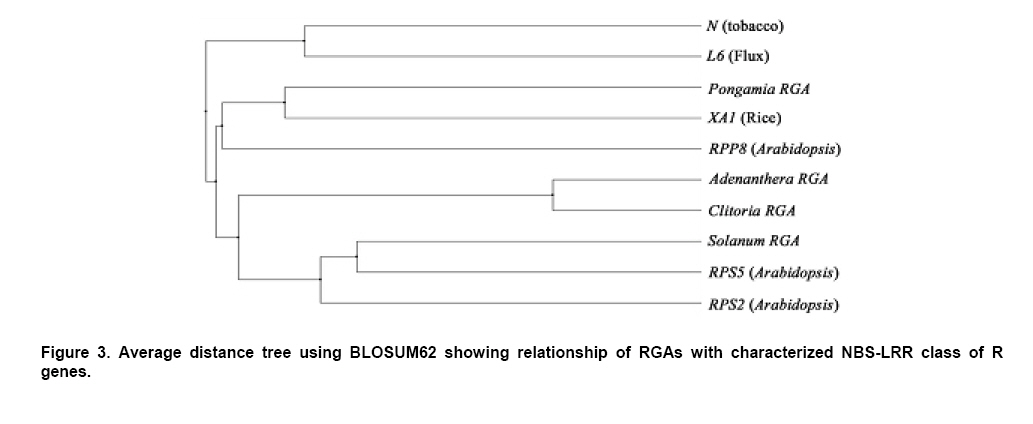
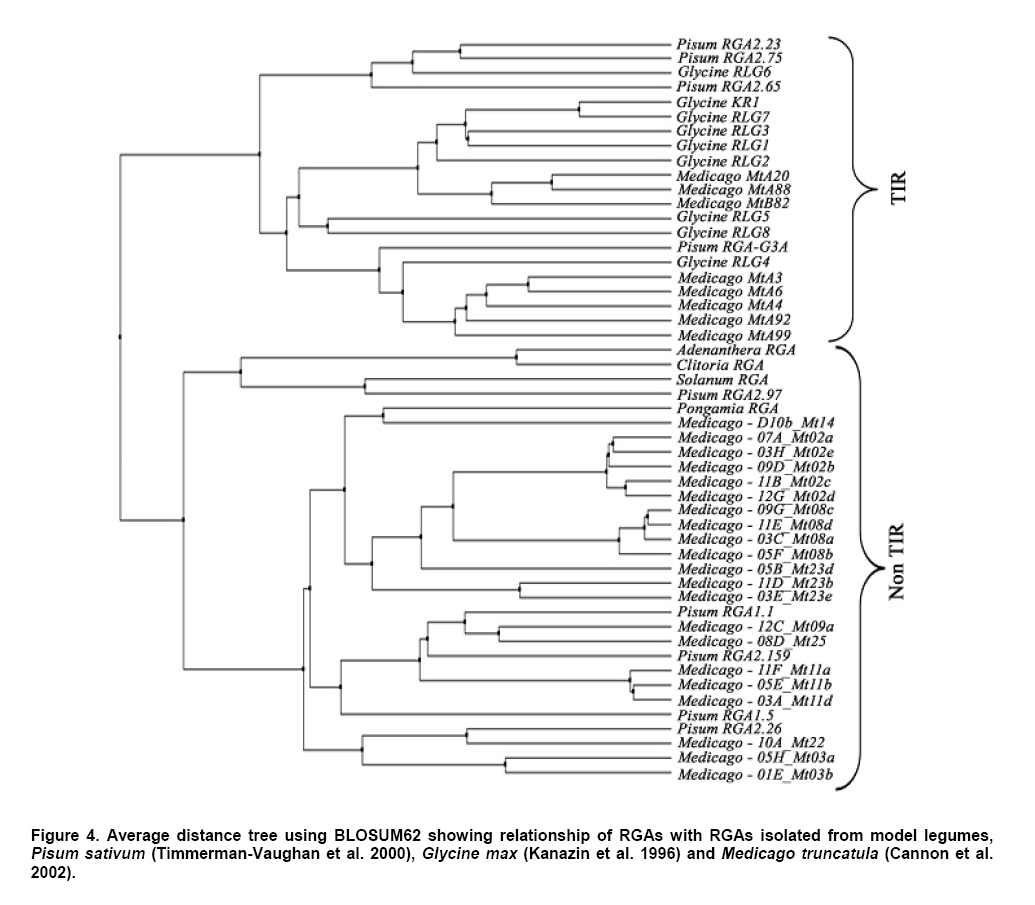
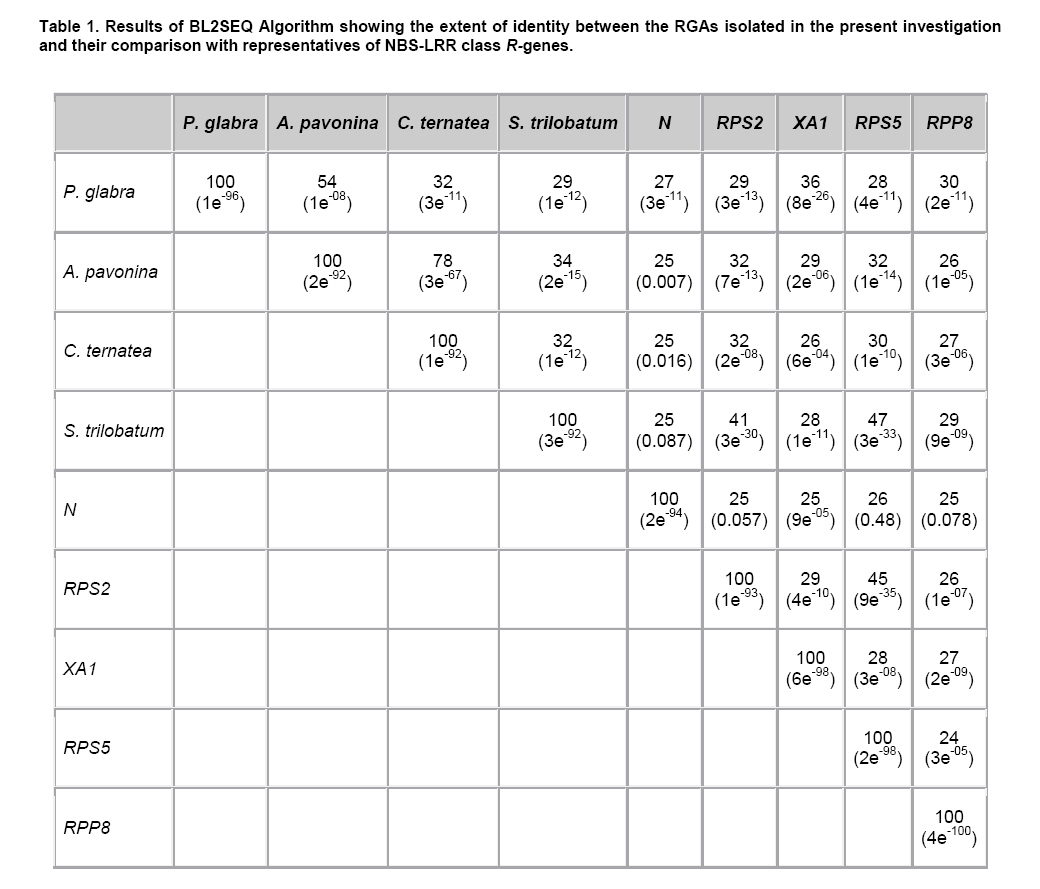
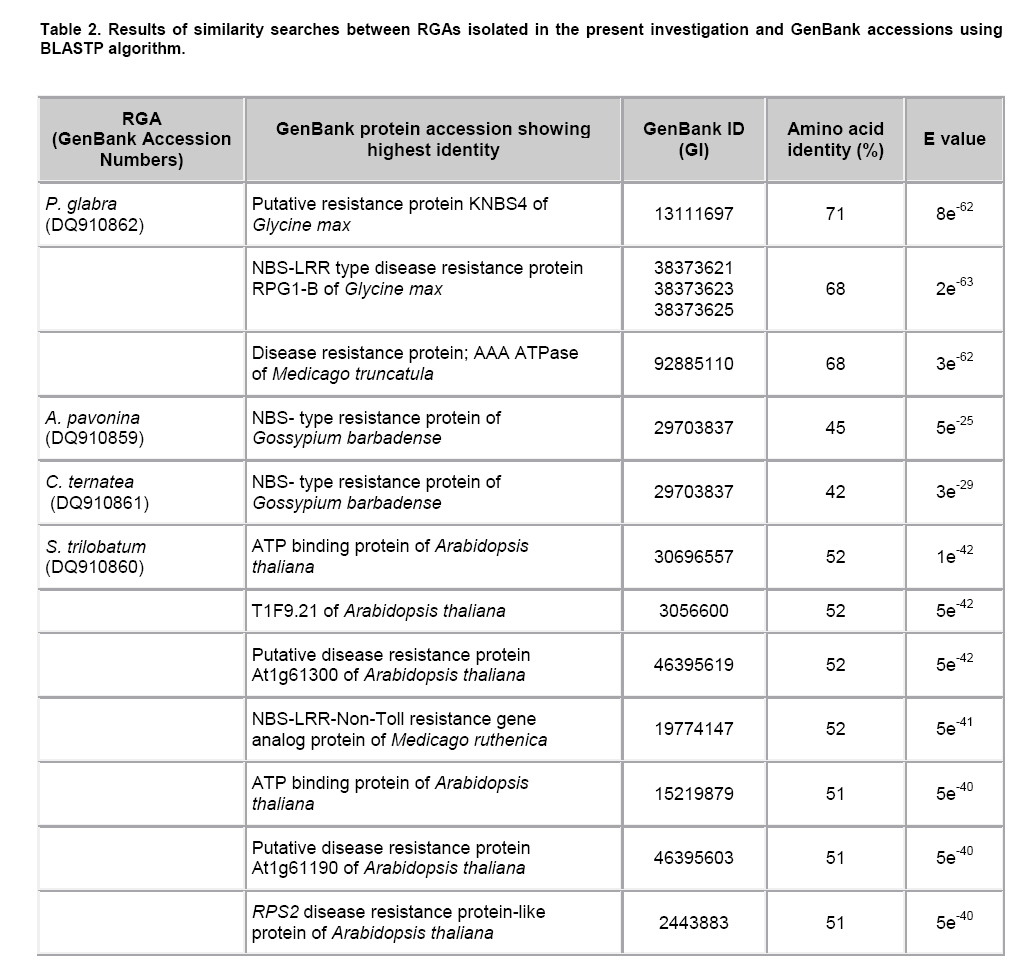
Plant disease resistance genes (R-genes) have been cloned and characterized from both mono- and dicotyledonous plants (Hammond-Kosack and Jones, 1997). More than 50 R-genes have been cloned so far from a variety of plant species (Wenkai et al. 2006). Among them, 37 share the NBS-LRR domain in their protein sequences and form the largest R-gene class (Bent et al. 1994; Lawrence et al. 1995; Kaloshian et al. 1998; Yoshimura et al. 1998). Genes from the NBS-LRR class confer resistance to bacteria, fungi, virus, nematodes and aphids (Timmerman-Vaughan et al. 2000). No function other than disease resistance has yet been assigned to this large class of genes (Bai et al. 2002). The NBS-LRR class resistance genes are proposed to act as receptors in signal transduction pathways that are triggered in response to pathogen attack (Hammond-Kosack and Jones, 1997). The NBS region is thought to be important for ATP binding and overall functionality of the R-gene product (Walker et al. 1982; Saraste et al. 1990). The NBS-LRR class of R genes have been cloned and well characterized from Arabidopsis thaliana, flax, tobacco, tomato and rice (Bent et al. 1994; Mindrinos et al. 1994; Whitham et al. 1994; Grant et al. 1995; Lawrence et al. 1995; Milligan et al. 1998; Yoshimura et al. 1998). The NBS region of characterized R-genes and R-gene analogs (RGAs) contains several highly conserved motifs, in spite of the diversity of pathogens against which they act. The P-loop and the kinase-2 domains have been described as ATP- and GTP- binding sites (Traut, 1994; Meyers et al. 1999). Additional motifs found in the NBS region of plant resistance genes are the kinase-3a and the GLPL (also called “hydrophobic domain”), a putative membrane spanning domain (Baldi et al. 2004). By making use of these conserved domains, NBS-sequences were obtained by PCR-mediated approach and provided candidate resistance genes (or Resistance Gene Analogs) in several crops such as potato (Leister et al. 1996), soybean (Kanazin et al. 1996; Yu et al. 1996; Peñuela et al. 2002; He et al. 2003), maize (Collins et al. 1998), sunflower (Gentzbittel et al. 1998), lettuce (Shen et al. 1998), Brassica (Joyeux et al. 1999), rice (Mago et al. 1999), common bean (Rivkin et al. 1999), citrus (Deng et al. 2000), coffee (Noir et al. 2001), chickpea (Huettel et al. 2002), grapewine (Donald et al. 2002), apple (Lee et al. 2003), wheat (Lacock et al. 2003), chicory (Plocik et al. 2004) and sorghum (Totad et al. 2005). The RGA fragments were also used as molecular markers for tagging the disease resistance loci in Arabidopsis thaliana (Aarts et al. 1998), wheat (Chen et al. 1998), rice (Ilag et al. 2000), melon (Mas et al. 2001), cowpea (Gowda et al. 2002), Lycopersicon (Zhang et al. 2002), common bean (Lopez et al. 2003), cocoa (Lanaud et al. 2004) and cotton (Hinchliffe et al. 2005). Pongamia glabra (synonym: Millettia pinnata; Family: Fabaceae), Adenanthera pavonina (Family: Fabaceae) and Clitoria ternatea (Family: Fabaceae) are among the few under-exploited non-edible legumes. Pongamia oil has immense potential as a synergist by suppressing the activity of detoxification enzymes in insects (Ramasubramanian and Regupathy, 2004). The seeds of A. pavonina contain thermo stable chitinases (Santos et al. 2004), which are one among the important pathogenesis related proteins. C. ternatea is resistant to a number of pests and pathogens owing to the presence of a defense protein designated as “finotin”. Finotin was found to be highly toxic to insects and important fungal pathogens of plants (Kelemu et al. 2004). C. ternatea is one of the important forage legumes having ornamental value, and the potential of which is being explored now through breeding programmes (Kalamani and Michael Gomez, 2001; Kalamani and Michael Gomez, 2002; Kalamani and Michael Gomez, 2003; Michael Gomez and Kalamani, 2003). Solanum trilobatum (Family: Solanaceae) has been in use for various ailments in different parts of India (Govindan et al. 1999, Madhavan and Balu, 1999) and its active principle sobatum was reported to possess anti-tumor activity (Mohanan and Devi, 1996; Mohanan and Devi, 1997). The leaf extract of S. trilobatum was reported to possess oviposition deterrent and skin repellent activities against the mosquito Anopheles stephensi (Rajkumar and Jebanesan, 2005). The insecticidal and antimicrobial activities of these underexploited plant species insisted to look for resistance gene analogs. In this paper we described the cloning, sequencing and diversity of NBS-LRR class of RGAs from P. glabra, A. pavonina, C. ternatea, and S. trilobatum from which no RGAs have yet been cloned. Materials and Methods DNA isolation and PCR amplification Genomic DNA from A. pavonina, C. ternatea, P. glabra and S. trilobatum was extracted by adopting CTAB procedure (Doyle and Doyle, 1990) with necessary modifications. Quality and quantity of the DNA was measured in NanoDrop® ND-1000 spectrophotometer. PCR amplifications were carried out in a 100 μl reaction containing 100 ng genomic DNA, 0.2 μM of each primer (designed from consensus sequences of NBS region 5’-GGTGGGGTTGGGAAGACAACG-3’, 5-CAACGCTAGTGGCAATCC-3’) (Leister et al. 1996), 0.25 mM each of the four dNTPs (Fermentas Life Sciences, Canada), 2 mM MgCl2, 1 x Taq buffer (GeNei, India) and 2.5 U Taq DNA polymerase (GeNei, India). PCR was performed in MyCyclerTM thermal cycler (BioRad, USA) with initial denaturation at 94ºC for 3 min, then cycling for 19 times using a touch-down strategy (an initial cycle of 94ºC for 30 sec, 63ºC for 30 sec, 72ºC for 1 min, then lowering the annealing temperature for each cycle by 0.5ºC during the following 18 cycles), followed by 19 cycles at 94ºC for 15 sec, 55ºC for 30 sec and 72ºC for 1 min then final extension at 72ºC for 10 min. Amplified PCR products were electrophoresed in 1.5% w/v agarose gels (Sigma-Aldrich, India) stained with ethidium bromide. PCR fragments were excised under UV transilluminator (Fotodyne, USA) and purified using GenEluteTM Gel Extraction Kit (Sigma Aldrich, USA). The purified PCR fragments of P. glabra, A. pavonina, C. ternatea and S. trilobatum were cloned into plasmid vector pTZ57R/T using InsT/AcloneTM PCR Product Cloning Kit (Fermentas Life Sciences, Canada) and transformed into competent Escherichia coli strain DH5α by following manufacturer’s instructions. The transformed colonies were screened and confirmed for the presence of insert by colony PCR. Plasmid DNA was isolated by adopting the protocol described by Birnboim and Doly (1979) with necessary modifications. The presence of the insert was confirmed by double digestion of plasmid DNA with EcoRI and BamHI restriction enzymes (Fermentas Life Sciences, Canada). Sequencing of the purified plasmid was done in an automated ABI 3100 Genetic Analyser based on Sangers’s sequencing method at GeNei, Bangalore, India. Diversity analysis of cloned DNA fragments using Bioinformatic tools The nucleotide sequences of the cloned fragments were translated in to amino acid sequences using ExPASy (Expert Protein Analysis System) Translate Tool, a proteomics server of Bioinformatics. The deduced amino acid sequences were subjected to motif analyses using the online version of CLUSTALW multiple alignment program of European Bioinformatics Institute (Thompson et al. 1997). The amino acid sequences of the RGAs were compared with protein sequences deposited in the GenBank using BLASTP algorithm (Altschul et al. 1997). Pair wise comparison of RGA sequences with known R-gene sequences at NBS region was made using BL2SEQ algorithm (Tatusova and Madden, 1999). In addition to these, amino acid sequences of RGAs isolated from the model legumes Glycine max (Kanazin et al. 1996), Pisum sativum (Timmerman-Vaughan et al. 2000) and Medicago truncatula (Cannon et al. 2002) were retrieved from the GenBank and were trimmed to start and end at the P-loop and GLPL motifs to facilitate accurate alignments with the sequences cloned in the present study. The Jalview Java alignment editor was used to construct average distance tree based on BLOSUM62 (Clamp et al. 2004). Results and Discussion PCR amplification of targeted RGA fragment The discovery of conserved motifs among the NBS-LRR class resistance genes opens the avenue for the use of PCR based strategy in isolating and cloning R gene family members and analogs using degenerate or specific primers. This approach thus provides an alternative to the classical methods of transposan tagging and map-based cloning strategies used so far (Seah et al. 1998). In earlier studies, co-amplification of non specific fragments apart from the expected amplicon (500-600 bp) was observed in crops like soybean (Yu et al. 1996), rice (Mago et al. 1999) and sorghum (Totad et al. 2005). Increasing the stringency of PCR condition by adopting higher annealing temperature, “hot start” and “touch-down” protocols were not successful in obtaining the more specific fragment in Cucumis melo (Brotman et al. 2002). However, in the present study we were successful in amplifying only the targeted RGA fragment (as was expected from the distance between the sequence motifs in N gene of tobacco and RPS2 of Arabidopsis thaliana) from the genomic DNA of A. pavonina, C. ternatea, P. glabra and S. trilobatum probably due to high stringency PCR conditions combined with touch-down strategy (Figure 1). Sequence analysis for conserved motifs The amino acid sequences of RGAs cloned in the present investigation were compared with other known R genes viz., N gene of tobacco, RPS2, RPS5 and RPP8 of A. thaliana, XA1 of rice and L6 of flax in NBS region using CLUSTALW multiple alignment program (Figure 2). The various motifs of NBS, i.e., kinase-1a, kinase-2 and kinase-3a were conserved in P. glabra, A. pavonina, C. ternatea, and S. trilobatum RGAs. The hydrophobic region represented by the GLPL domain was also observed in all the four RGA sequences. High sequence homology of the RGAs with NBS motifs of N, RPS2, RPS5, RPP8, L6 and XA1 clearly establishes that the RGAs cloned in the present investigation might belong to NBS-LRR class of R-genes. The NBS-LRR genes are usually grouped into two different subfamilies (Meyers et al. 1999; Pan et al. 2000): subfamily I contains the TIR element (Toll-Interleukin-1 Receptor-like domain) and has been found only in dicots, while subfamily II lacks the TIR domain and has been found both in dicots and monocots. The partial sequence of the NBS portion is usually sufficient to assign a given gene to either subfamily I or II. The last residue of the kinase-2 domain can be used to predict with 95% accuracy whether an RGA would belongs to the TIR-NBS or the non-TIR-NBS family; conservation of tryptophan (W) at this location is tightly linked to non-TIR R-genes (RPS2, RPS5 and RPP8 of A. thaliana), whereas conservation of aspartic acid (D) or its uncharged derivative aspartate (N) is characteristic of TIR class of R-genes (N and L6) (Meyers et al. 1999; Pan et al. 2000; Jeong et al. 2001; Peñuela et al. 2002). The conservation of tryptophan (W) in the P. glabra, A. pavonina and S. trilobatum RGAs with the exception of C. ternatea indicates that these belong to the non-TIR subfamily of NBS-LRR genes. A. pavonina clone is likely to be a pseudogene because it contains a stop codon, which might have evolved during the course of evolution. In TIR-NBS-LRR group, a characteristic consensus motif FXXXXF and a highly conserved glycine is present between kinase1a and kinase-2 domains, whereas the non-TIR-NBS-LRR group contains the consensus sequence FXXXXW (Pan et al. 2000). The presence of consensus motif FXXXXW between kinase 1a and kinase-2 domains of P. glabra and S. trilobatum RGA sequences further ensures their position in non-TIR-NBS-LRR subfamily. Phylogenetic analysis of RGA sequences with already characterized R-genes was done by constructing average distance tree using BLOSUM62 in the Jalview Java alignment editor. The RGAs of P. glabra, A. pavonina, C. ternatea and S. trilobatum were clustered together with well- characterized non-TIR-NBS-LRR genes RPS2, RPS5 and RPP8 of A. thaliana, while the TIR-NBS-LRR genes N and L were grouped as a separate cluster (Figure 3). Another average distance tree was constructed for the RGAs cloned in this study and those isolated from the model legumes Glycine, Pisum and Medicago. The RGAs were grouped in to two major clades each represents the TIR and Non-TIR NBS-LRR subfamilies of R genes (Figure 4). The RGAs cloned from P. glabra and S. trilobatum showed high similarity with Medicago-D10b_Mt14 RGA and Pisum RGA 2.97, respectively. The Pisum RGA 2.97 has already been reported to share high similarity with the well-known non-TIR-NBS-LRR gene RPS2 (Kanazin et al. 1996). The RGAs isolated from A. pavonina and C. clitoria were clustered together in a single subclade and exhibit their sequence similarity. Comparison of RGAs with known R-genes using BL2SEQ algorithm The RGAs cloned in the present study were compared with representative NBS-LRR class R-genes viz., N gene of tobacco, RPS2, RPS5 and RPP8 of A. thaliana and XA1 of rice in NBS region using BL2SEQ algorithm (Tatusova and Madden, 1999). The R-gene analogs showed comparable identity with Non-TIR-NBS-LRR genes and TIR-NBS-LRR gene. However, all the four RGAs had lowest E values with Non-TIR-NBS-LRR genes, RPS2, RPS5, RPP8 and XA1 as against the TIR-NBS-LRR gene, N (Table 1). This confirms the strong match with Non-TIR-NBS-LRR gene family. The extent of amino acid identity between the representative NBS-LRR class R-genes was also analysed for comparison using the BL2SEQ algorithm. The percentage of identity at NBS region between N and RPS2 (25%), N and XA1 (25%), RPS2 and XA1 (29%) (Table 1) indicates that the homology between them was mainly restricted to the different motifs of NBS, thereby indicating sequence diversity among the NBS-LRR class resistance genes. The extent of identity between the RGAs themselves varied between 29% and 78%, which reveals the diversity among the RGAs isolated in the present study. BLAST search for identical protein sequences It was well established that the protein level search and comparison showed more homology with NBS-LRR region of many RGAs than at the nucleotide level due to degeneracy of the genetic code. Thus, pair wise comparison at amino acid level is more reliable than nucleotide-nucleotide comparison for establishing the identity of any RGA (Totad et al. 2005). Hence, the nucleotide sequences were translated in to polypeptides using ExPASy translate tool and identity search was made using BLASTP algorithm. All the four R-gene analogs showed high similarity with RGAs deposited in the GenBank (Table 2). P. glabra RGA had 71% identity (E value 8e-42) with putative resistance protein KNBS4 of Glycine max (GI: 13111697) followed by 68% identity (E value 2e-63) with NBS-LRR type disease resistance protein RPG1-B of G. max (GI: 38373621; GI: 38373623; GI: 38373625) and disease resistance protein, AAA ATPase of Medicago truncatula (GI: 92885110). The lowest E values confirm that the matches are more reliable and not due to chance.The RGAs identified in A. pavonina and C. ternatea showed 45% (E value 5e-25) and 42% (E value 3e-29) identity with NBS- type resistance protein of Gossypium barbadense (GI: 29703837). S. trilobatum RGA exhibited strong matches with the amino acid sequences of ATP binding protein of A. thaliana (GI: 30696557; Identity- 52%; E value 1e-42), T1F9.21 of A. thaliana (GI: 3056600; Identity- 52%; E value 5e-42), putative disease resistance protein At1g61300 of A. thaliana (GI: 46395619; Identity- 52%; E value 5e-42) followed by NBS-LRR-Non-Toll resistance gene analog protein of Medicago ruthenica (GI: 19774147; Identity- 52%; E value-5e-41). The E value was taken in to consideration for the appropriateness of the match. The expected (E) value refers to the number of matches expected by chance alone; lower the E value indicates that the match is more reliable and not occurred purely due to chance (Di Gaspero and Cipriani, 2002; Totad et al. 2005). Studies on R-genes and RGAs are still explorative in nature. Information on more R-gene sequences is necessary to delineate more structural domains, which is the basis for the search of RGAs in any crop plant (Totad et al. 2005). The diversity among the novel RGAs characterized in this study may lead to mining of R- genes in other non edible legumes and wild Solanum species. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08045t2.jpg] [ej08045f3.jpg] [ej08045f1.jpg] [ej08045f4.jpg] [ej08045f2.jpg] [ej08045t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}