 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 11, No. 4, October 15, 2008 RESEARCH ARTICLE Kinetic study on inducibility of polygalacturonases from Aspergillus flavipes FP-500 Aurora Martínez-Trujillo1 , Juan S. Aranda2 , Guillermo Aguilar-Osorio*3 1Laboratorio de Catálisis
Enzimática,
Tecnológico de Estudios
Superiores de Ecatepec,
Av. Tecnológico s/n esq. Av. Carlos Hank González,
Ecatepec, Estado de México, CP 55210, México
Tel: 01 55 00 23 00. Anexo 2227
E-mail: amartinezt@tese.edu.mx Financial support: This work was financially supported by the National Council for Science and Technology (Mexico), the Technologic Institute for Higher Studies of Ecatepec (México), the Research Office of the National Polytechnique Institute (Mexico), and the National University of Mexico, DGAPA projects IN207603 and IN209008. Received March 25, 2008 / Accepted June 16, 2008 Code Number: ej08049 Abstract The aim of this work was to describe growth dynamics, substrate depletion and polygalacturonases production by Aspergillus flavipes FP-500 in batch cultures by means of unstructured models. The microorganism was cultivated on several mono- di- and poly- saccharides, and then the culture development modeled with Monod and Leudeking-Piret equations. The kinetic parameters related to the models (μmax, γx/s, α and β) were obtained by minimizing the quadratic residuals function with a simplex algorithm. An accurate description of experimental data was attained with the proposed models. Besides, modeling provided significant kinetic information on microbial degradation of complex substrates, such as the correlation between specific growth rate μmax and production yield α, suggesting that A. flavipes FP-500 polygalacturonases are actually constitutive, but also that there is a certain degree of induciblility in these enzymatic activities. Keywords: constitutive enzymes, inducible enzymes, initial pH, kinetic modeling, pectinases. Pectin is a complex polysaccharide found in middle lamella of plant cell wall. It represents the first plant barrier during microbial attack which involves the action of several enzymes (de Vries and Visser, 2001). Fungi of the genus Aspergillus are among the microbial species that can degrade pectin, so producing a number of pectinases (Texeira et al. 2000). The importance of pectin-degrading enzymes lies mainly in their actual and potential uses in a number of industries, like in food processing, textile industry, paper and pulp industry, pectic wastewater treatment and animal feed, among others (Jayani et al. 2005; Niture, 2008). In addition, the understanding of the regulation process of the production of polygalacturonases will contribute to get insights in the molecular dialogue between the host and the pathogen, during microbial invasion of plant cell wall (Esquerré-Tugayé et al. 2000; Lang and Dörnenburg, 2000). Although the specific signal molecule that triggers pectinases synthesis remains unknown, the general accepted idea is that fungal cells produce a low level of constitutive pectinolytic enzymes, which release a few molecules of mono or oligosaccharides from a polymer in plant structure. These small molecules are transported into the microorganism and start a massive expression of the degrading pectinases (Mach and Zeilinger, 2003). This would mean that several pectinolytic activities are induced by a number of substrates (Prade et al. 1999). Pectinases production is regulated in different fungal species, depending on the carbon source (Crotti et al. 1998; Wubben et al. 2000; Olsson et al. 2003) and the pH of the medium (de Vries and Visser, 2001; Peñalva and Arst, 2002). The mathematical modeling of microbial growth and process performance has led to improved design and operation of mycelial fermentations and has improved the ability of scientists to translate laboratory observations into commercial practice. Unstructured modeling represents a particular and useful application of growth mathematical analysis that could provide some important hints on constitutiveness and inducibility of metabolites production, such as some enzymatic activities, on a kinetic, quantitative basis. In this context, the present work aimed at describing the dynamics of fungal growth, substrate depletion and polygalacturonases production in batch cultures under different conditions of carbon source and initial pH of the culture, using the Monod and Leudeking-Piret unstructured models (Thilakavathi et al. 2007). After obtaining the kinetic parameters, their numerical values were correlated in order to point out the constitutive or inducible nature of the produced polygalacturonases. Materials and Methods Aspergillus flavipes FP-500, was isolated in Mexico from rotten tomatoes. The strain was maintained at 4ºC on PDA agar plates. The strain was identified by conventional methods considering its morphological characteristics growing on different media and by microscopic examination. A comparison with type strains was done resulting in the identification of the strain as member of flavipes species. A. flavipes FP-500 was grown on basal medium, containing (g L-1): K2HPO4, 2; KH2PO4, 2; and (NH4)2SO4, 5. The medium was sterilized by autoclaving at 121ºC for 20 min. The initial pH of the medium was adjusted with 2 M NaOH or H2SO4. To this basal salt solution, a suitable carbon source was added to attain a final concentration of 10 g L-1. Spores were collected from 3-day-old agar slants with Saline-Tween solution (NaCl, 0.9% and Tween 80, 0.01%). A final concentration of 1 x 106 spores/mL was obtained by adjusting with sterile water. Monosaccharide based media (see below) were supplemented with 0.1% (w/v) of yeast extract. Production of enzymes under different carbon sources and initial pH In order to evaluate the effect of carbon source and initial pH on growth and enzyme production, A. flavipes FP-500 was grown on basal medium supplemented with different carbon sources: pectin and polygalacturonic acid, as complex polysaccharides; galacturonic acid, arabinose, rhamnose and xylose, as the main monosaccharide constituents of pectin; glycerol as a simple substrate not related to pectin structure; lactose and glucose, this latter considered a universal catabolic repressor. Also, three initial pH values of the culture media were established (3.5, 4.2 and 5.0), in order to evaluate microbial growth and polygalacturonases production under these different conditions. Cultures were carried out in 100 mL of medium, and were incubated at 37ºC in a reciprocant shaker at 200 rpm. Samples were withdrawn every 24 hrs until 72 hrs of culture. Microbial biomass. Cell growth was measured by dry weight and expressed as g L-1. Polygalacturonase activity. This was measured by determination of the reducing sugars produced from 1% (w/v) pectin solution after incubation at 45ºC for 20 min at pH 5.0. One unit of polygalacturonase activity was defined as the amount of enzyme that catalyzes the formation of 1 μmol of galaturonic acid under assay conditions (Trejo-Aguilar et al. 1996). Substrate depletion. Monosaccharide substrates consumption was measured through quantification of reducing sugars using 3,5-dinitrosalicilic acid, with the corresponding sugar as the reference standard (Miller, 1959). Polymers consumption was estimated after acid hydrolysis of polysaccharides containing samples with a sulfuric acid and following color development of the phenol reagent (Dubois et al. 1956). Culture behavior in time is described with the following equations:
Where
μmax maximal specific growth rate (h-1) S substrate concentration (gs L-1) ks Monod saturation constant (gs L-1) x biomass concentration (gb L-1) Yx/s biomass on substrate yield coefficient (gb gs-1) α growth-associated coefficient for pectinases production (U gb-1) β growth-independent coefficient for pectinases production (U gb-1 h-1) Monod constants μmax and ks are not simultaneously identifiable in batch process (Nihtila and Virkkunen, 1977; Chouakri et al. 1994), so the maximal specific growth rate is the only useful obtained parameter. Kinetic parameters in the Leudeking-Piret model, α and β, indicate the relation between growth and pectinases production in a fungus culture. The parameter α expresses the enzyme production associated with microorganism growth, in a way that production is considered growth-associated whenever α ≠ 0. Since pectinases production is needed for the assimilation of complex substrates in order the miocroorganism to grow, parameter α is the more meaninful factor in our modeling approach. Besides, the estimation of β resulted in a nil value for any Aspegillus flavipes culture. The former considerantions lead to specify the vector parameter P as: P = [μmaxγx/s α] and the initial conditions to model equations (x0, S0 and p0) are the experimental concentrations at process time t = 0. The growth yield (Yx/S) and the identifiable parameters for Monod (μmax) and Luedeking-Piret (α) models were obtained as reported in a previous work (Aranda-Barradas et al. 2000), with the following residuals function F as the minimizing criterium:
Where superscripts exp and mod indicate experimental data and model results, respectively. The differences between experimental and theoretical values were calculated for the n sample points (n = 12). The estimated kinetic parameters Pest are those that satisfy the condition F (Pest) = min Meaning that the estimated parameters are chosen in order to produce the minimum residual errors between the model theoretical values and the experimental data. The search of the minimum was carried out with a Nelder-Mead simplex method, given an arbitrary initial vector P0 to start up the algorithm. The model can be coupled to estimated parameters Pest in order to create synthetic data sets, by adding white noise to numerical results on x, S and p from the model. Each synthetic data set produces a new vector of equivalent estimated parameters. Therefore, calculating a number of equivalent parameters allows the estimation of the mean and the standard deviation for every kinetic parameter (Table 1). Variance estimations in the model Variance associated to parameters. Confidence intervals of the best estimated parameters are given by
Where σ standard deviation for each kinetic parameter (i = μmax, γx/s or α), obtained from synthetic data m experimental determinations made in N samples N samples obtained from each culture q number of estimated parameters t0.975 Student’s factor for a confidence level αt = 0.975 Estimated variance to x, S and p. Variance of experimental data obtained after estimation of biomass (x), substrate (S) and exopectinases (p) was estimated by
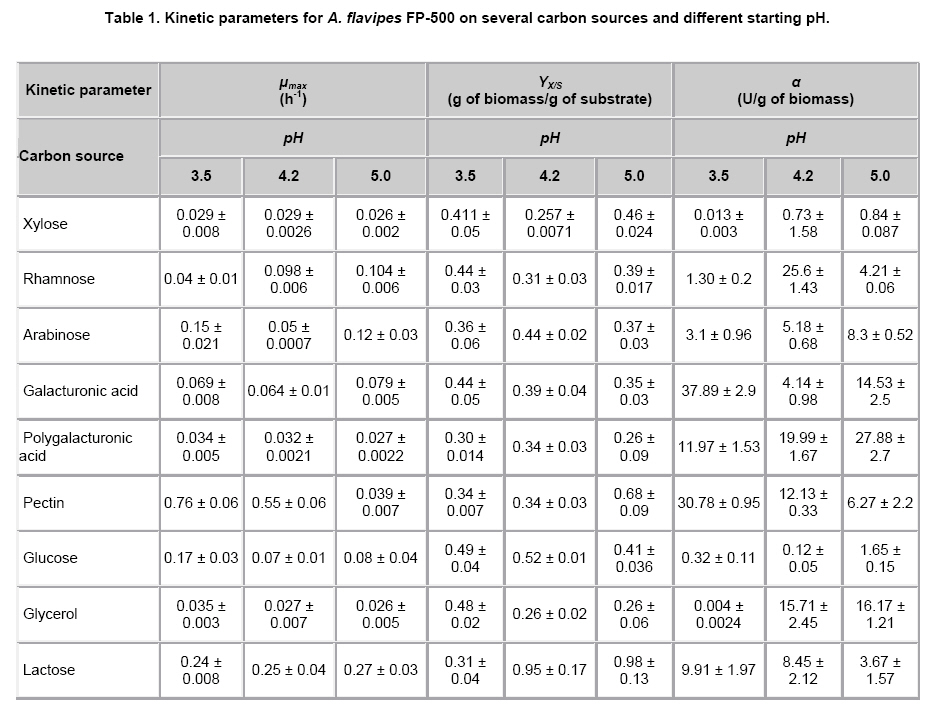
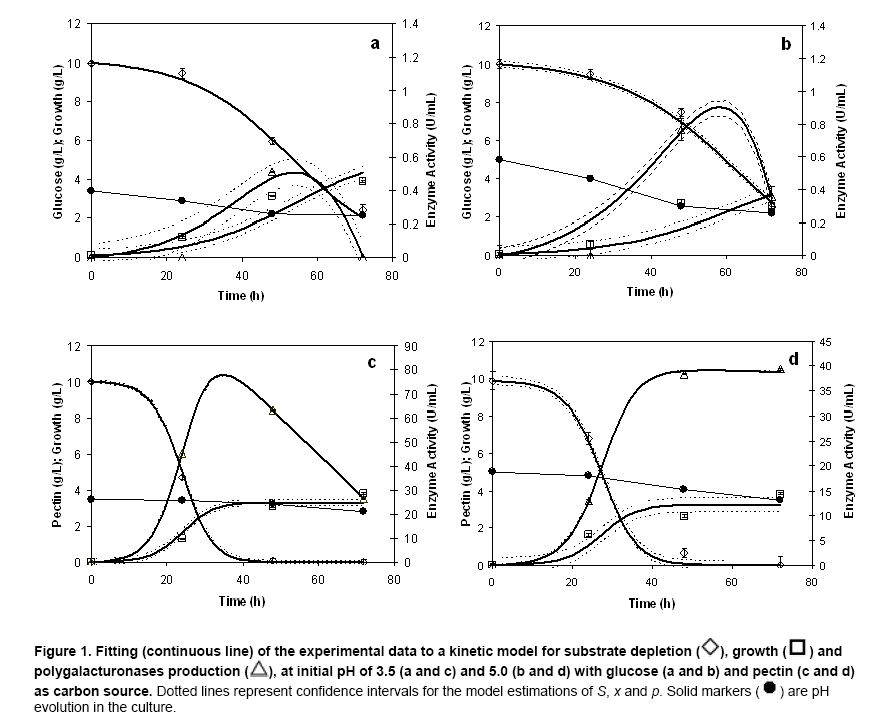
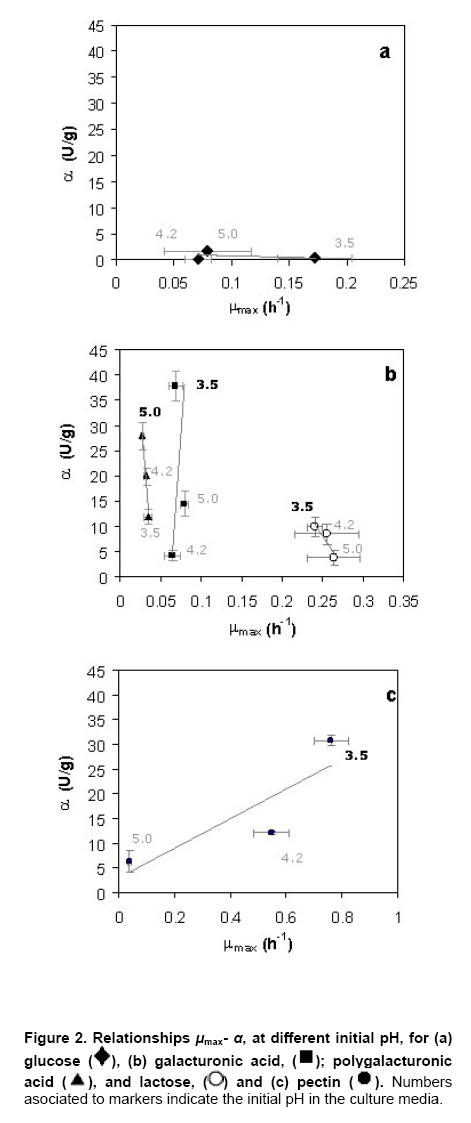
where u x, S or p Results Modeling of growth and pectinases production Kinetic parameters (μmax, γx/s, α) estimated by Nelder-Mead simplex algorithm (Table 1) showed to be accurate enough to build a reliable model that describes the kinetics of Aspergillus flavipes FP-500 on several carbon sources, as it can be seen from the model fitting in Figure 1. A good fit to experimental data was reached with this mathematical treatment. Kinetic parameters as a function of pH and substrate Kinetic parameters for the mathematical model were determined in each experimental condition described before. Monod-type growth on substrates such as xylose, glycerol and polygalacturonic acid resulted in maximal specific growth rates around 0.03 h-1, while on galacturonic acid μmax ≈ 0.07 h-1 and on lactose μmax was about 0.25 h‑1. With all the forementioned substrates the maximal specific growth rate was approximately constant regardless the initial pH in the culture. Thus, for these substrates, estimated μmax values seem to indicate dependence of growth on the carbon source, but not on the initial pH of the culture. However, estimated μmax for other group of substrates (glucose, rhamnose, pectin) was strongly affected by the initial pH. On the one hand, experiments with rhamnose as a carbon source showed increasing μmax values as the initial pH was rised from 3.5 to 5.0. Growth on glucose and pectin presented a significative drop of the specific growth rate μmax when the initial pH in the culture was increased. Concerning polygalacturonases production represented by the Leudeking-Piret model, estimated β values were equal to zero for all the substrates and initial pH tested. In contrast, α parameter estimations showed a dependence on both the substrates and the initial pH. The lowest α value was obtained for glycerol (0.004 U/g, pH = 3.5), meanwhile high values were obtained for rhamnose (25.6 U/g, pH = 4.2), galacturonic acid (37.89 U/g, pH = 3.5), polygalacturonic acid (27.88 U/g, pH = 5.0) and pectin (30.78 U/g, pH = 3.5). Values obtained for pectin and galacturonic acid were exceptionally high when compared to other substrates in media with initial pH of 3.5. Similarily, for glycerol, the product yield α is relatively high (15.71 U/g for pH = 4.2 and 16.17 U/g for pH = 5.0) although this substrate is not a structural component of pectin. Polygalacturonases constitutiveness and inducibility from kinetic Parameters After comparing kinetic parameters for different carbon sources and initial pH values, some interesting remarks can be established concerning the relationship between estimated μmax and α (Figure 2). Three major trends were observed: i) For glucose, plotting μmax vs. α it could be observed that the estimated μmax values strongly depended on the pH of the medium (Figure 2a). While α value at pH 3.5 and 4.2 was similar at 5.0 a higher value was obtained. However, this latter value was relatively low as compared with other substrates. ii) For substrates as lactose, galacturonic acid or polygalacturonic acid, estimated μmax was the same on the three initial pH tested. However, different α values were obtained, depending on the initial pH used for each medium. A graphical representation of μmax – α parameters shows a linear, nearly vertical, relationship (Figure 2b). iii) With pectin as the carbon source, there is a positive proportional relationship between μmax and α (Figure 2c). These experimental results could be interpreted in terms of constitutiveness or inducibility of pectinases as a function either of initial pH in the culture media or of the carbon source. Discussion It can be clearly seen on Figure 1 that although final biomass for growth on glucose and pectin is similar, glucose is not depleted in the process time, contrary to pectin which is exhausted after 48 hrs. These findings suggest that A. flavipes FP-500 is better adapted to complex polysaccharides than to monosaccharides, since kinetics of growth is faster on the former substrates. It should be observed that μmax varied in different carbon sources, with a clear trend to be higher for lactose and pectin than for monosacharide susbtrates. It is interesting to notice that this strain was able to grow on lactose as a sole carbon source and this is not usual. Other filamentous fungi showed a low growth rate on this substrate (Pakula et al. 2005), even when this carbon source has been reported as the inducer of cellulases in Hypocrea jecorina (anamorph of Trichoderma reesei), (Seiboth et al. 2004). The growth on lactose is related to the presence of β-galactosidase activity, a key enzyme for lactose utilization by this fungus. Considering that lactose does not occur in the natural environment of fungi, it has been proposed that the function of this β-galactosidase is the hydrolysis of terminal non-reducing β-D-galactose residues in plant cell wall components including hemicelluloses or pectins (Seiboth et al. 2007). Furthermore, in Aspergillus there is not a clear picture about lactose utilization; while A. nidulans is able to grow on lactose and galactose, A. niger does not (Seiboth et al. 2007). As it can be seen from our results, A. flavipes grows on lactose and is able to produce α- and β-galactosidase. So, it is probable that A. flavipes FP-500 is using a similar strategy to A. nidulans that allow the growth on lactose. Nevertheless, it would be necessary to develop additional experiments for demonstrating this hypothesis. The highest μmax values were reached with data from a pectin culture, so pectin allows a good microbial biomass production in this strain. Besides, this behavior could explain the elevated polygalacturonases production of the fungus when it grew on pectin as carbon source. A. flavipes FP-500 is a good pectinases producer, whose pectinases production is better than other Aspergillus strains, such as those reported for A. niger F-1119, which produced barely 10.8 U/ml and 2.1 g/L of biomass on citrus pectin (Shubakov and Elkina, 2002). The α parameter involved in Leudeking-Piret model expresses the enzyme production associated with the growth of the microorganism. Moreover, analysis of μmax - α plots can give some insight about constitutiveness, if α remains constant eventhough changes in μmax are presented; or inducibility, if α increases regardless the behavior of μmax. For growth on glucose (Figure 2a), no matter what initial pH was adjusted in the culture, low values for α were obtained. The numerical values were not higher than 1.65 U/g what is considered relatively low, and might be reflecting a basal enzyme activity. Taking into account that glucose is not a constituent of pectin structure, the behavior observed in this μmax – α relationship suggests that there is a constitutive part of polygalacturonases produced by A. flavipes FP-500, which is expressed under any pH condition. With other substrates unrelated to pectin structure (glycerol and lactose) α ranged from 0.004 to ~15 U/g. However, α raised up to 25 U/g or more for cultures developed on carbon sources related to pectin. Among them, the highest was attained on galacturonic acid, and it was comparable in magnitude to those observed for polygalacturonic acid and pectin. This seems to indicate a certain substrate-inducibility degree in the enzymatic production. When a graphical analysis between μmax and α was performed for pectin-related substrates, different behaviors were observed. On galacturonic acid, the fungus had the same maximal specific growth rate (~0.07 h-1) but different α values for every initial pH tested (Figure 2b). This suggests that on galacturonic acid there is an induction in this strain of polygalacturonase activity depending on initial pH, as it has been reported for other Aspergillus species (de Vries et al. 2002). On polygalacturonic acid (Figure 2b) it was clear that initial pH of the medium also conditioned the polygalacturonases activity. It seems that for almost all the substrates, the initial pH of 3.5 increases polygalacturonases activity. Thus, initial pH conditions regulate pectinases activity. Some similar conclusions have been reached in induction and repression research on cellulases by A. niger (Hanif et al. 2004). However, although our results on kinetic modeling certainly give some insight on polygalacturonases production, constitutiveness and inducibility, there is still room for other confirmatory experiments. Concluding Remarks Aspergillus flavipes FP-500 is a pectinases producer strain. Our results have shown that polygalacturonases production is strongly influenced by the available carbon source, and that enzyme activity is regulated by the initial pH in the culture medium. The main contribution lies in the kinetic characterization of polygalacturonases production by the strain, that allowed to stress to some extent the constitutiveness and inducibility of those enzymes: i) Growth on monomeric substrates unrelated to pectine (glucose, gycerol) resulted in a low polygalacturonases production, considered a basal constitutive enzyme activity, that can be modified by means of a change in the initial pH of the culture medium. ii) Culture media containing monomeric substrates related to pectin (xylose, rhamnose, arabinose) produced in general a low polygalacturonases yield (α), eventhough some important increases in enzymatic activity are attained by establishing the appropiate initial pH (3.5 for galacturonic acid and 4.2 for rhamnose) in the culture medium. iii) Complex substrates such as pectin or polygalacturonic acid induced important polygalacturonase production that was significantly increased at low pH (3.5) for pectin, or at a slightly higher pH (5.0) for polygalacturonic acid. Even though kinetic parameters are not definitive evidence of enzyme induction, an indirect inference from μmax - α relationship would indicate the conditions for an increase of enzymatic activity, mainly due to substrate induction. Acknowledgments The authors acknowledge Dr. Edgar Salgado for the critical reading of the manuscript. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej08049f1.jpg] [ej08049t1.jpg] [ej08049f2.jpg] |
| |||||||||

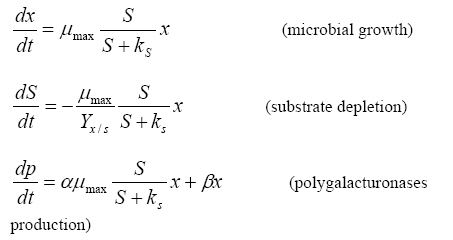

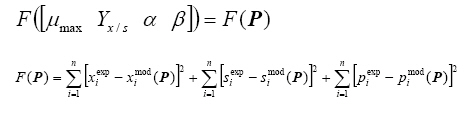
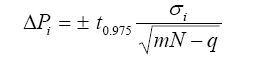
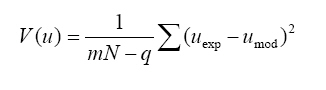
{kind=link}
{kind=link}
{kind=link}