 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 12, No. 2, April 15, 2009 Influence of metal ions and organic carbons on denitrification activity of the halotolerant bacterium, Paracoccus pantotrophus P16 a strain from shrimp pond Punyawatt Pintathong1 , David J. Richardson2 , Stephen Spiro3 ,Wanna Choorit*4,
1 Biotechnology Program,
School of Agricultural Technology,
Walailak University,
Thasala, Nakhon Si Thammarat 80160,
Thailand Financial support: This work was supported by The Royal Golden Jubilee (RGJ) Ph.D. Program (2.B.WL/45/A.1). Received May 22, 2008 / Accepted January 21,2009 Code Number: ej09011 Abstract The effect of metal ions, ferric ion (Fe3+) and molybdenum ion (Mo6+) on the denitrification process of Paracoccus pantotrophus P16 grown under saline conditions was investigated. Results revealed that the dosages of added Fe3+ and Mo6+ significantly accelerated nitrate utilization and nitrite accumulation. Enzymatic studies revealed that the membrane-bound nitrate reductase and the periplasmic nitrite reductase had activities of 998 ± 28 and 373 ± 18 nmol (mg protein)-1 min-1, respectively after growing Paracoccus pantotrophus P16 in medium supplemented with 1.5 µM Fe3+. If provided with 1.5 µM Fe3+and 2.4 µM Mo6+, the membrane-bound nitrate reductase activity increased to 6,223 ± 502 nmol (mg protein)-1 min-1 and the periplasmic nitrite reductase was 344 ± 20 nmol (mg protein)-1 min-1. The results indicated that an addition of Fe3+ and Mo6+ led to an overstimulation of nitrate reductase activity as compared with nitrite reductase activity. When glucose was supplied, the minimal ratio of carbon per nitrate (C/N) was 2.31 mg C/mg NO3--N with denitrification yield of 0.45 g NO3--N/g C. Addition of ethanol instead of glucose, the minimal ratio of C/N was 1.15 mg C/mg NO3--N with denitrification yield of 1.08 g NO3--N/g C. Keywords: metal ions, nitrate reductase, nitrite accumulation, nitrite reductase, Paracoccus pantotrophus.
Waste nitrogen containing compounds, of both organic nitrogen (protein and amino acids) and inorganic nitrogen (ammonia, nitrate and nitrite), can be transformed to nitrogen gas by the action of microorganisms (van Rijn et al. 2006). The first step is ammonification in which all the organic-N is converted to ammonia. This is then oxidized by nitrifying bacteria to nitrate and then converted to nitrogen gas by denitrification (Crab et al. 2007). Inorganic nitrogen, in the form of ammonia, nitrate and nitrite have been considered as compounds that can be toxic to aquatic animals (Cheng and Chen, 2001; Jensen, 2003; Kir et al. 2004; Yildiz and Benli, 2004), and create environmental problems, such as eutrophication of rivers. Some attempts to overcome these problems have been conducted using good agricultural practices (Mishra et al. 2008), and in many areas local authorities have enforceable environmental regulations on the maximum concentrations of nitrogen compounds permitted in effluent discharges (van Rijn et al. 2006). This has lead to an increased emphasis on treating wastewater with high nitrogen levels, for example by means of biological methods. Paracoccus spp. are capable of reducing nitrate to nitrogen gas (Rezić et al. 2006) and are widely found in nitrogen rich environments. As reported by Foglar et al. (2005) Paracoccus was the dominant bacterial species found after wastewater from an industrial yeast process was biologically treated. Paracoccus halodenitrificans was isolated from sediments and used to reduce levels of perchlorate and nitrate (Okeke et al. 2002). It is known that many parameters, such as oxygen level, temperature, pH, nitrate concentration, carbon per nitrate (C/N) ratio, NaCl concentration and trace elements influence denitrification (Cervantes et al. 1998; Glass and Silverstein, 1998; Bilanovic et al. 1999; Glass and Silverstein, 1999; Carrera et al. 2003; Estuardo et al. 2008). Denitrification involves several proteins that require metal ions as a cofactor (Ferguson, 1998; Philippot and Højberg, 1999). Mo6+ acts as a component of the molybdenum cofactor of nitrate reductase, while Fe3+ is required for the cytochrome subunits of both nitrate and nitrite reductases. A high dosage of trace metals (Fe, Cu, Zn, Mo and Mn) caused an increase in denitrification rate from 200 to 700 g NOx-N/d. However, only Fe was proved to be a significant influence on rates of denitrification, and the combination of Fe + Mn had greater effect (Labbé et al. 2003). Cyplik et al. (2007) reported that an addition of Fe, Cu and Mo significantly increased the specific nitrate reduction rate of Haloferax denitrificans. The nature of the carbon substrate also influences denitrification rates. For example an accumulation of nitrite was 5 mg NO2-/l when sucrose was used as a carbon source (Gómez et al. 2000), but ethanol and methanol supported a higher denitrification rate as compared to sucrose (Lorrain et al. 2004). Thus, in order to complete denitrification (N2 gas formation) the factors influencing the denitrification process must be investigated. In this study, we isolated a salt tolerant bacterium Paracoccus pantotrophus P16 from a mud sample obtained from a shrimp pond at Nakhon Si Thammarat, in the southern part of Thailand. Glucose and ethanol were selected as carbon sources supporting denitrification in this bacterium. Glucose is normally used as the carbohydrate source for shrimp feeds, and also for controlling water quality (Shiau, 1998; Avnimelech, 1999; Hari et al. 2004; van Rijn et al. 2006; Asaduzzaman et al. 2008). On the other hand, ethanol is superior in terms of high denitrification rates with low biomass production (Gómez et al. 2000; Lorrain et al. 2004). The aims of this study were to determine the influence of added metal ions and carbon sources on the denitrification process catalysed by Paracoccus pantotrophus P16.
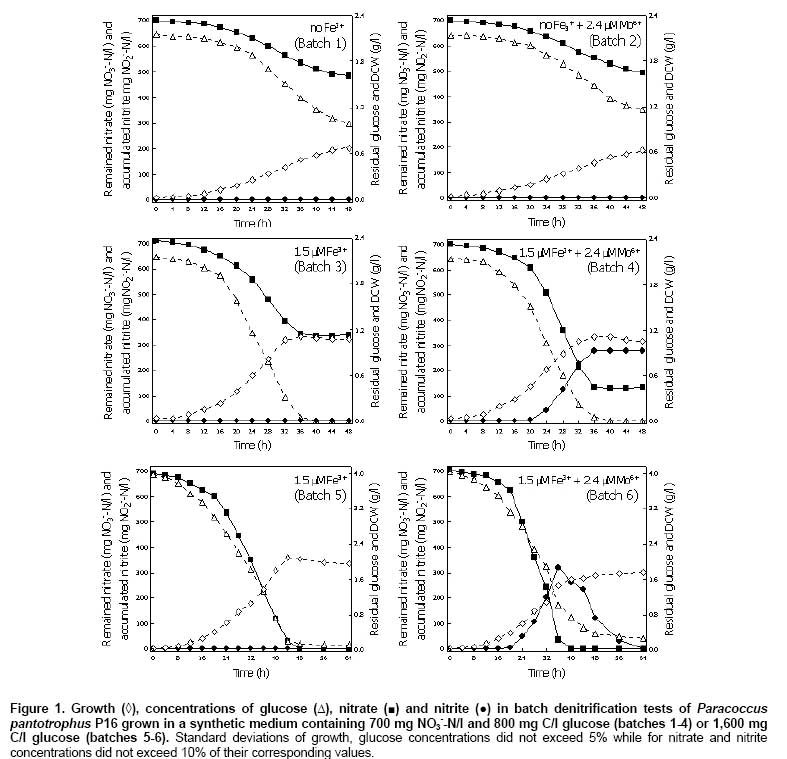
Bacteria, media and starter culture preparation The bacterial strain P16 was isolated from a mud sample obtained from a shrimp pond in Nakhon Si Thammarat, Thailand. It was identified as a member of Paracoccus and closely related to Paracoccus pantotrophus based on its morphological, biochemical and physiological properties, and 16S rDNA sequencing. Under denitrifying conditions, it can grow in medium containing NaCl up to 6%. Growth was observed at 25-47ºC. Sugar substrates, i.e. glucose, fructose, sucrose and maltose were excellent carbon sources for growth, while growth in the presence of methanol was not observed. The bacterium can tolerate and reduce nitrate and nitrite at about 2,100 mg NO3--N/l and 400 mg NO2--N/l to N2 gas, respectively. Moreover, it can carry out aerobic denitrification (unpublished data). The synthetic medium used was modified from Sasaki et al. (1988) and contained, g/l, 0.5 KH2PO4, 0.5 K2HPO4, 0.8 (NH4)2HPO4, 0.2 MgSO4•7H2O, 0.053 CaCl2•2H2O, 0.001 MnSO4•5H2O, 0.01 yeast extract, 30 NaCl and x glucose or ethanol with the pH of the medium adjusted to 7.5. For denitrifying growth, FeCl3•6H20 and Na2MoO4•2H2O were also supplemented as sources of Fe3+ and Mo6+, respectively. Glucose (2 g/l) was used as a carbon source for starter culture preparation. The starter cultures were prepared by cultivating Paracoccus pantotrophus P16 in 500 ml Erlenmeyer flasks containing 200 ml synthetic medium, and incubated in an orbital shaker at 150 rpm at 35ºC until growth reached late log phase. Cells were then harvested by centrifugation at 12,000 rpm for 15 min. To prepare the inoculum, the cell pellets were washed twice and readjusted to a cell concentration (optical density at 660 (OD660)) of 1.0 with fresh medium. A 5% (100 ml) of starter culture was inoculated into the medium. Effect of Fe3+, Mo6+ and carbon sources on the denitrification process Preliminary studies showed that among metal ions to be tested, Fe3+, Mn2+, Cu2+, Zn2+, and Mo6+, only Fe3+ and Mo6+ affected on growth and denitrification rates of the bacterium P16; Mn2+ enhanced growth when added with Fe3+, and nitrate removal increased corresponding to growth. These data agreed well with the previous results (Labbé et al. 2003; Zhou et al. 2007). Taking these results and those reported by Labbé et al. (2003) and Zhou et al. (2007) together led us to focus attention on the effect of Fe3+ and Mo6+ on denitrification. To investigate the influence of Fe3+ and Mo6+on the denitrification process, each concentration of metal ion was added into the synthetic medium supplemented with 5 g/l KNO3 (700 mg NO3--N/l) and 2 g/l glucose (800 mg C/l) (Figure 1). This C/N ratio was 1.15. All batch cultures were grown in reactors with a 12 cm internal diameter and a height of 27 cm. The reactors were maintained at 35 ± 1ºC using hot water circulation. Samples were withdrawn from sampling holes (5 mm in diameter) located 8 cm from the bottom. The reactors were incubated under static conditions and were gently mixed to produce a homogenous mixture before sampling. Each batch culture was made anaerobic by flushing it with N2 gas prior to inoculation of the bacterial cells and after each sampling. At predetermined time intervals, samples were taken and used to investigate growth, carbon consumption, nitrate and nitrite concentrations. The experiments were done in triplicate. In order to study the minimal levels of glucose and ethanol required by Paracoccus pantotrophus P16 to convert nitrate at a concentration of 700 mg NO3--N/l to N2 gas, the carbon per nitrate ratios (C/N ratio, mg C/mg NO3--N) at 1.15, 1.73 and 2.31 were examined. The starter culture was inoculated into the synthetic medium containing predetermined C/N ratios as desired. At the specified concentrations metal ions were added (Figure 1). The reactors were operated as previously described. Subcellular localization and enzyme activity assays In order to study enzymatic activities of subcellular fractions and in whole cells, the bacteria were cultivated in reactors under the above described conditions for 24 hrs. The synthetic medium was supplemented with 5 g/l KNO3 and 2 g/l glucose and different concentrations of metal ions (Table 1). The cells were harvested by centrifugation at 12,000 rpm at 4ºC and washed twice with 10 mM phosphate buffer, pH 7.5. Subcellular fractions were obtained by preparing type II spheroplasts as described by Bell et al. (1990). Nitrate and nitrite reductase activities were measured in whole cells and subcellular fractions by following the substrate-dependent oxidation of dithionite-reduced benzyl viologen (BV+) (Bell et al. 1990).
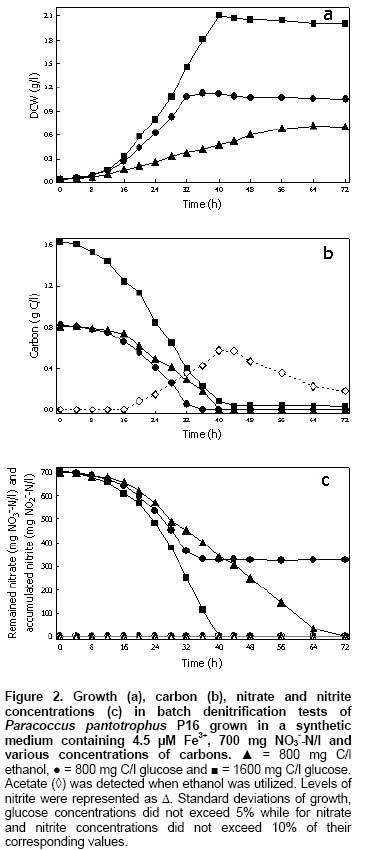
Nitrate and nitrite reduction by intact cells Nitrate and nitrite reductase activities of bacterial cells grown in the presence of nitrate (700 mg NO3--N/l) and two carbon substrates, glucose and ethanol, (800 mg C/l) were measured as described by Kariminiaae-Hamedaani et al. (2004). During the growth, the cells were collected at predetermined time intervals, by centrifugation and then washed 3 times with 0.1 M potassium phosphate buffer, pH 7.2. The concentration of cell suspensions were adjusted to 1.0 (OD660) with 0.1 M potassium phosphate buffer. Subsequently, 5 ml of cell suspensions were transferred into reaction tubes and 0.1 ml of concentrated solution of nitrate or nitrite was added into each tube to provide a final concentration of 10 mg NO3--N/l or 10 mg NO2--N/l, respectively. The reaction was carried out at 35ºC. In the clear supernatant, the remaining concentration of nitrate and nitrite was measured. The enzymatic reduction of nitrate and nitrite was defined in terms of milligrams of nitrate and nitrite remained per liter, respectively. Bacterial growth was measured as turbidity by use of a spectrophotometer (Schimadzu, UV-1601, Kyoto, Japan) at a wavelength of 660 nm and correlated with the dry cell weight (DCW) measured by drying the cells at 105ºC to constant weight for 12 hrs. Protein concentrations were determined using the bicinchonic acid assay with bovine serum albumin as the standard. Nitrate and nitrite were analyzed using FIAstarTM 5000, version 1.3 (FOSS Analytical AB, Höganäs, Sweden) by automated cadmium reduction and the sulfanilamide colorimetric methods, respectively. Glucose, acetate and ethanol were determined by HPLC with Metacarb H Plus (Varian) and a refractive index detector (Waters). Membrane proteins (10 µg total protein) separated by sodium dodecyl sulphate - polyacrylamide gel electrophoresis (SDS-PAGE) were blotted onto PVDF membrane (Amersham Biosciences). The blots were probed with a polyclonal primary antibody to Paracoccus NarG that was raised in sheep against purified NarGH protein. Antibody bound to the ~140 kDa NarG was detected using an anti-sheep alkaline phosphatase-conjugated secondary antibody. Antibodies were used at a concentration of ~1 μg /ml. Results Effect of Fe3+, Mo6+ and carbon sources on the denitrification process The time profiles of nitrate removal and nitrite accumulation during anaerobic growth of Paracoccus pantotrophus P16 are shown in Figure 1. In the absence of Fe3+ nitrate removal was low, such that the remaining nitrate, from the 700 mg NO3--N/l added, in batches 1 (no Fe3+) and 2 (no Fe3+ + 2.4 µM Mo6+) was 483.5 and 493.7 mg NO3--N/l, respectively. The comparison with batch 3 (1.5 µM Fe3+) showed that in the presence of Fe3+ the remaining nitrate concentration was much lower at 337.4 mg NO3--N/l. Furthermore, an addition of Fe3+ together with Mo6+ (batch 4) resulted in a rapid decrease in nitrate concentration and the remaining nitrate was only 132.1 mg NO3--N/l and under this condition nitrite accumulated to 277.7 mg NO2--N/l. Figure 1 (batch 4) also demonstrates that after 36 hrs to the end of cultivation, the nitrate and nitrite concentrations remained at a constant level. At this point all the glucose substrate had been removed. However, the accumulation of nitrite might indicate that nitrate is a more favoured electron acceptor for this bacterium under this condition as compared with nitrite. A comparison of the calculated specific growth rates (µ) of batches 1 - 4 showed that increasing Fe3+, resulted in increased µ, from 0.07 h-1 in the absence of Fe3+ (batch 1) to 0.124 h-1 in the presence of Fe3+ (batch 3). The presence of Fe3+ also led to the complete consumption of glucose and biomass levels were doubled from 0.67 g dry cell/l to 1.10 g dry cell/l. The addition of Mo6+ did not increase the growth rates (batch 2) even though it did have a big effect on nitrate removal when in combination with Fe3+ (batch 4). In the experiment described so far 800 mg C/l glucose and 700 mg NO3--N/l nitrate were used as sole sources of carbon and electron acceptor, respectively. From the batch 3 it can be calculated that 800 mg C/l glucose can effectively convert about 365 mg NO3--N/l to N2 gas in the presence of added Fe3+. Figure 1 (batch 5) shows that 700 mg NO3--N/l was completely reduced to N2 gas within 48 hrs when the initial glucose concentration was increased to 1,600 mg C/l glucose and Fe3+ were present. In this case the levels of nitrite were always very low (0-1.5 mg NO2--N/l) since more electrons were available for reduction of both nitrate and nitrite. Growth of the bacterium under complete denitrification conditions was increased to 2.08 g dry cell/l. So an increase in growth of 100 mg dry cell/l caused by addition of Fe3+ and glucose required the utilization of about 33.4 mg NO3--N/l. Addition of Mo6+ and Fe3+ had effects on growth and nitrate utilization. It was observed that within 40 hrs, nitrate exhausted. Again, a high rate of nitrate removal was achieved in the presence of both Fe3+ and Mo6+ with an initial glucose concentration of 1600 mg/l but under these conditions nitrite accumulated transiently (Figure 1) (batch 6). At 36 hrs, the levels of nitrate and accumulated nitrite in batch 6 were 35.1 mg NO3--N/l and 320.2 mg NO2--N/l, respectively. After nitrate exhausted, then nitrite was utilized. The maximum biomass of batch 6 was 1.75 g dry cell/l. Thus growth of 100 mg dry cell/l caused by addition of Fe3+, Mo6+ and glucose required the utilization of about 40.2 mg NO3--N/l. In the presence of Fe3+ and Mo6+, growth of the bacterium was retarded and nitrite was accumulated. These experiments also showed that under saline conditions, Paracoccus pantotrophus P16 consumed 700 mg NO3--N/l (without net accumulation of nitrite) when 1,600 mg C/l of glucose was added (Figure 1) (batches 5 and 6). Time courses of growth, carbon concentration, nitrate concentration and accumulation of nitrite are shown in Figure 2. In the reactors containing 800 mg C/l glucose and 1,600 mg C/l glucose growth of the bacterium were 1.12 and 2.11 g dry cell/l, respectively and in the presence of ethanol growth was 0.72 g dry cell/l (Figure 2a). The results showed that both substrates were completely consumed within 36-44 hrs. However, as ethanol was consumed the bacterium produced acetate (Figure 2b). The C/N ratio need to remove 700 mg NO3--N/l varied according to the carbon source added. When ethanol was supplied at 800 mg C/l, the initial ratio of C/N was 1.15 mg C/mg NO3--N. In growth experiments with glucose the C/N ratio was either 2.31 mg C/mg NO3--N or 1.15 mg C/mg NO3--N (Figure 2). In the presence of glucose when C/N ratio was 1.15 only about half of the nitrate was used but it was all consumed at this initial C/N ratio when ethanol was used (Figure 2c). Interestingly, the nature of compound used as the carbon source (glucose and ethanol) and the C/N ratios did not cause nitrite accumulation by the bacterium. It should be noted that when ethanol was used instead of glucose in starter culture preparation step the results were same as those in Figure 2 (data not shown). Table 2 shows that if provided with ethanol, maximum dry cell weight, denitrification rate and denitrification yield were 0.72 ± 0.02 g/l, 9.68 ± 0.11 mg NO3--N/l/h and 1.08 ± 0.02 g NO3--N/g C, respectively. When using glucose, denitrification rate increased to 17.56 ± 0.18 mg NO3--N/l/h, while maximum dry cell weight and denitrification yield were 2.11 ± 0.02 g/l and 0.45 ± 0.01 g NO3--N/g C, respectively. The comparison of specific denitrification rate in term of mg NO3--N/h/g DCW revealed that ethanol was superior to those of glucose.
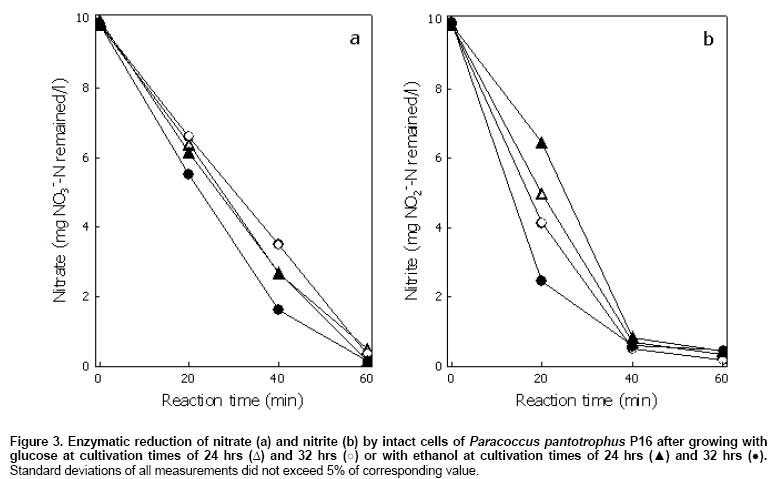
Subcellular localization and activity of nitrate and nitrite reductases Testing of the 3 subcellular fractions, cytoplasmic, periplasmic and membrane fractions, for nitrate and nitrite reductase activities showed, in all cases, that nitrate reductase was present in the membrane fraction while nitrite reductase was found in the periplasmic fraction. As shown in Table 1, the membrane-bound nitrate reductase and periplasmic nitrite reductase activities were 998 ± 28 and 373 ± 18 nmol (mg protein)-1 min-1, respectively after growing Paracoccus pantotrophus P16 in the medium supplemented with 1.5 µM Fe3+. Of special interest was the finding that further addition of 2.4 µM Mo6+ further increased the nitrate reductase activity to 6,223 ± 502 nmol (mg protein)-1 min-1. However, the periplasmic nitrite reductase activity of 344 ± 20 nmol (mg protein)-1 min-1 was slightly less than that in the absence of 2.4 µM Mo6+. It seems likely that the greater imbalance of the 2 enzymes caused by the addition of Mo6+ led to the accumulation of nitrite. Table 1 also shows nitrate and nitrite reductase activities of whole cells. Nitrate reductase and nitrite reductase activities were 44 ± 4 and 18 ± 3 nmol (mg DCW)-1 min-1, respectively when medium was supplemented with 2.4 µM Mo6+ into the medium. Nitrate reductase activity was 240 ± 30 while nitrite reductase activity was 162 ± 13 nmol (mg DCW)-1 min-1 in the culture containing 1.5 µM Fe3+. This clearly indicated the effect of Fe3+ toward the enzymes activities. On the contrary, Fe3+ and Mo6+ supported nitrate reductase activity while they slightly decreased nitrite reductase activity. The profiles of enzyme activities of whole cells and of subcellular fractions were similar. However, the results of enzyme activities in subcellular fractions were largely different with the whole cell. This due to benzyl viologen must crossed cell membranes of the bacterium prior to react with the enzymes. Nitrate and nitrite reduction by intact cells Enzymatic reduction of nitrate and nitrite in phosphate buffer containing 10 mg NO3--N/l nitrate or 10 mg NO2--N/l nitrite by intact cells of Paracoccus pantotrophus P16 after growth on glucose or ethanol was investigated (Figure 3). After growing cells for 24 and 32 hrs, the profiles of nitrate and nitrite reduction by intact cells were similar irrespective of the carbon substrate. Furthermore, it can be observed that under the tested conditions, the whole cells can hold the activities of nitrate and nitrite reducing enzymes over 60 min. Also both enzymes had the activities in the presence of 3% NaCl. Discussion Martienssen and Schöps (1999) observed that the accumulation of nitrite during denitrification processes has been shown to occur by (1) inhibition of nitrite reductase by nitrate due to competitive utilization between nitrate and nitrite to serve as the terminal electron acceptors, (2) inhibition of nitric oxide reductase by nitrate and the accumulated nitric oxide causing further inhibition of nitrite reductase by (3) the extreme sensitivity of nitrite reductase to inhibition by low oxygen concentrations, (4) an imbalance of nitrate and nitrite reductase activities, and (5) a delayed expression of nitrite reductase mRNA reflected in a delay to the synthesis of nitrite reductase when compared with nitrate reductase. In this experiment, denitrification activity was influenced by the concentrations of Fe3+ and Mo6+. When Fe3+ was the only ion added, nitrate and nitrite were synergistically reduced to N2 gas. Addition of Fe3+ and Mo6+ led to nitrite accumulation although a greater rate of nitrate removal was observed (Figure 1). Enzymatic assays showed that increasing levels of Fe3+ produced increased levels of both membrane-bound nitrate and periplasmic nitrite reductase activities. 1.5 µM Fe3+ produced a 2.6 and 1.7 fold increase, respectively. However, as mentioned previously the increases in membrane-bound nitrate reductase activity were greatly increased by the further addition of Fe3+ and Mo6+. Addition of Fe3+ and Mo6+, caused the membrane-bound nitrate reductase activity to increase up to 16-21 fold while the increase in periplasmic nitrite reductase activity was only about 1.5-3 fold (Table 1). Therefore an overstimulation of membrane bound nitrate reductase activity as compared to periplasmic nitrite reductase activity caused an accumulation of nitrite. It is already known that the membrane-bound nitrate reductase (Nar) enzyme required by bacterial denitrifiers contains a Mo-molybdopterin complex (González et al. 2006). Tavares et al. (2006) also reported that Fe and Mo are required for membrane-bound nitrate reductase enzymes. An increase in the concentration of Fe2+ or Mo6+ to 70 mg/dm3 or 5 mg/dm3, respectively resulted in an increase in the nitrate removal (Cyplik et al. 2007). The periplasmic nitrite reductases of Paracoccus spp. are cytochrome cd1 type, which contained hemes as a major component (Moura and Moura, 2001; Einsle and Kroneck, 2004). Although, trace elements, i.e. Cu, Zn and Mn were found in yeast extract; comparison the results of batch 1 with the others were clearly indicated the effect of adding metal ions (Figure 1). Figure 1 (batches 5 and 6) shows that adding glucose to 4 g/l led to complete nitrate reduction. Even the C/N ratio was 2.31 mg C/mg NO3--N; transient accumulation of high nitrite concentration was observed when Fe3+ and Mo6+ were present. In addition, the accumulated nitrite in the presence of Fe3+ and Mo6+ still remained when culture medium was limited (Figure 1) (batch 4). This accumulated nitrite could be highly toxic to human and aquatic animals when it was contaminated to environments (Chow and Hong, 2002; Jensen, 2003). Importantly, many researches reported that the presence of nitrite accumulated during the denitrification process can promote an accumulation of nitric and nitrous oxides, reflecting incomplete denitrification processes (Schulthess et al. 1995; Béline et al. 1999; Alinsafi et al. 2008). The emission of nitrous oxide (N2O) also causes undesirable environmental problems, such as the green house effect and ozone depletion in the stratosphere (Park et al. 2000). Therefore, metal ion concentration is the critical parameter that affects the ability of denitrifying bacteria to remove nitrate and nitrite. It is emphasized that the amount of Fe3+ and Mo6+ to be added in the treatment system should be carefully controlled. In order to control denitrification rate, without an accumulation of nitrite, a suitable concentration of trace metals should be supplied. Under experiments to be tested, the membrane-bound nitrate reductase had 2.7 fold higher activity than periplasmic nitrite reductase activity was the optimized conditions to control denitrification rate. The results showed that nitrate reductase was found in the membrane fraction and was therefore regarded as a true respiratory enzyme to drive ATP synthesis for anaerobic nitrate respiration. In contrast the periplasmic nitrate reductase can be referred to as a dissimilatory enzyme because it can be expressed under both aerobic and anaerobic conditions but it cannot generate proton motive force during electron transfer in the quinol-Nap span (Steenhoudt et al. 2001; Gates et al. 2003). Western blot analysis was performed and showed that the intensity of the ~140 kDa band of NarG, a catalytic subunit of membrane-bound nitrate reductase, was similar under conditions containing only Fe3+ or Fe3+ and Mo6+ (data not shown). The results imply that added Fe3+ and Mo6+ may not have a direct effect on the expression of membrane-bound nitrate reductase, but they may have a direct influence on enzyme activity. Several types of carbon source have been reported for supporting heterotrophic denitrification, but capability to use these substrates can be different in different microorganisms (Kesserü et al. 2002; Kariminiaae-Hamedaani et al. 2004; Cyplik et al. 2007). In this study, the application of glucose and ethanol in the process enabled complete removal of 700 mg NO3--N/l without accumulation of nitrite. The C/N ratio for complete denitrification when glucose was supplied was 2.31 mg C/mg NO3--N. Whereas for ethanol it was 1.15 mg C/mg NO3--N with an efficient denitrification yield of 1.08 ± 0.02 g NO3--N/g C. Also, as reported by Her and Huang (1995) and Gómez et al. (2000), the optimal C/N ratio suggested for complete of denitrification was 1.08 and 2.00 for ethanol and glucose, respectively. On addition of glucose, the biomass increased as also observed by Cyplik et al. (2007). Although glucose was reported to be a poor substrate for denitrification (Etchebehere et al. 2001, Cyplik et al. 2007), but a greater denitrification rate of 17.56 ± 0.18 mg NO3--N/l/h was obtained by Paracoccus pantotrophus P16 grown in the presence of glucose. The experiment with ethanol showed a greater effect in term of specific denitrification rate (13.46 ± 0.48 mg NO3--N/h/g DCW). Therefore, an application of glucose and ethanol containing waste as sole of carbon source for denitrification in nitrate/nitrite-containing wastes by Paracoccus pantotrophus P16 is an alternative for a cost effective process. The whole cells to be collected from cultures broth could reduce nitrate and nitrite. The activities of nitrate and nitrite reduction can sustain over 60 min; therefore it is possible to apply the cells for decreasing nitrate and nitrite contents in water, foodstuffs, vegetable and fruit juices (Walkowiak-Tomczak, 2002; Okafor and Ogbonna, 2003). Concluding Remarks This work has reported the effects of metal ions, Fe3+ and Mo6+, carbon sources, C/N ratios on the growth, the rate of nitrate removal, nitrite accumulation and nitrate and nitrite reductase activities of a halotolerant Paracoccus pantotrophus P16 growing under saline environments and under denitrifying conditions. The presence of both Fe3+ and Mo6+ caused an accumulation of nitrite; this was consistent with the observed nitrate and nitrite reductase activities. The bacterium had an ability to remove 700 mg NO3--N/l within 40 hrs when 1,600 mg C/l glucose and 4.5 µM Fe3+ were added. If given 800 mg C/l ethanol, 700 mg NO3--N/l was removed within 72 hrs. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2009 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej09011f1.jpg] [ej09011f3.jpg] [ej09011f2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}