 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 12, No. 2, April 15, 2009 Molecular characterization of ‘Candidatus Liberibacter' species/strains causing huanglongbing disease of citrus in Kenya Titus O. Magomere*1 , Silas D. Obukosia2 , Eunice Mutitu3 , Christopher Ngichabe4 , Florence Olubayo5 , Solomon Shibairo6
1Plant Science and Crop Protection Department,
University of Nairobi,
P.O. Box 29053-00625, Nairobi, Kenya,
Tel: 254 02 631314
Fax: 254 02 0632121
E-mail: magomeretito@gmail.com Financial support: Rockefeller Foundation (Forum). Received November 20, 2007 / Accepted October 16,2008 Code Number: ej09012 This study was undertaken to characterize the alpha subgroup of the proteobacteria causing the huanglongbing (HLB) disease of citrus from three different ecological zones of Kenya namely the Lower highlands (LH2, LH3, 1800-1900 m above sea level); Upper midlands (UM3, UM4, 1390-1475m), Lower midlands (LM5, LM4, LM3 of 1290-1340-1390m), by isolation and sequencing DNA encoding the L10 and L12 ribosomal proteins and the intergenic region. A 7I6-basepair DNA fragment was amplified and sequenced and consisted of 536 basepairs of DNA encoding the L10 protein, 44 basepairs of DNA intergenic region and 136 basepairs of DNA that partially encodes the L12 protein. Sequences of rpL10/L12 protein genes from Kenyan strains were 98% and 81% similar to the South African ‘Candidatus Liberibacter africanus strain Nelspruit' and the Asian ‘Candidatus Liberibacter asiaticus' strains, respectively. The intergenic rDNA sequence of Kenyan strain from UM and LM showed 84% similarity with ‘Candidatus L. africanus strain Nelspruit' and 50% similarity with ‘Candidatus L. asiaticus' strain. However, the LH strain had an 11- basepairs deletion, while the LM4 had a 5-basepair deletion in the intergenic region compared to ‘Candidatus L. africanus strain Nelspruit'. The L10 amino acid sequence was 100% homologous among HLB bacteria obtained from the agro-ecological zones in Kenya and the L10 protein sequence was also homologus to ‘Candidatus L. africanus strain Nelspruit'. Nevertheless, the L10 amino acid sequence of ‘Candidatus L. asiaticus' and the ‘Candidatus L. africanus subsp. capensis' differed from the Kenyan strains by 18.36% and 11.82%, respectively. Phylogenetic analysis of both the L10/L12 rDNA sequences and the L10 amino acid sequences clustered the Kenyan strains of the ‘Candidatus Liberibacter' species with members of alpha subdivision of proteobacteria. Keywords: citrus greening disease, DNA sequencing, intergenic regions, phylogeny, ribosomal protein genes.
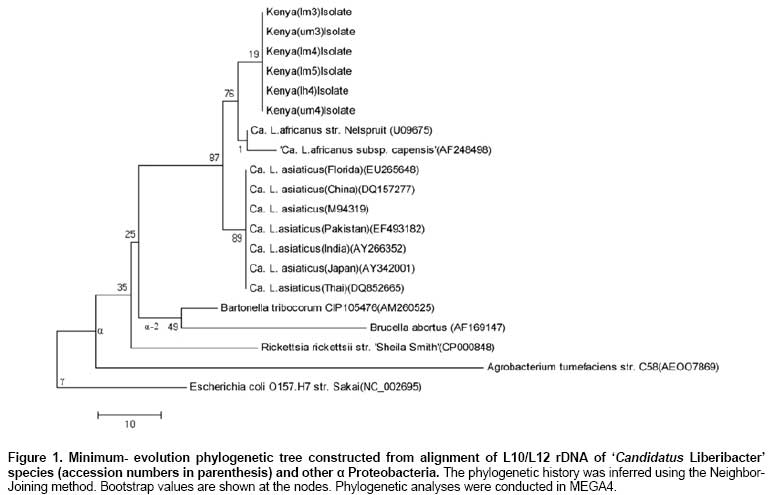
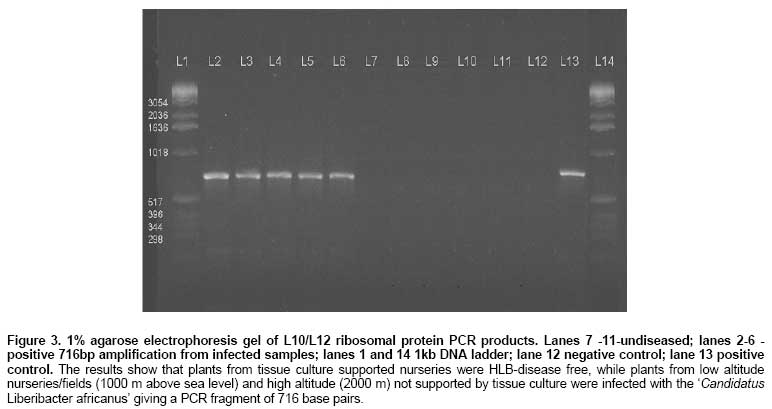
Huanglongbing (HLB) disease of citrus, also called the citrus greening disease was first reported in China in 1919 and in Africa in 1929. It is a severe and widespread disease of citrus in Asia, south-east Asia, south and eastern Africa, the Arabian peninsular (Garnier and Bové, 1993) and the Americas (Coletta-Filho et al. 2005; Teixeira et al. 2005a). In Africa it has been reported in Ethiopia, Kenya, Malagasy, Sudan, Swaziland, Tanzania and Zimbabwe (Bové, 2006). HLB disease is caused by an uncultured obligate, rod shaped, gram-negative phloem restricted bacterium that has recently been characterised (Jagoueix et al. 1994). The ‘Candidatus' generic name Liberibacter as defined by Murray and Schleifer (1994), for uncultured organisms has been given to the bacterium. The Liberibacters from Asia and Africa belong to two different ‘Candidatus' species, namely Liberibacter asiaticus and Liberibacter africanus (Villechanoux et al. 1993; Jagoueix et al. 1994; Planet et al. 1995; Hung et al. 2004). Recently, a third species ‘Candidatus Liberibacter americanus' was identified in Brazil (Coletta-Filho et al. 2005; Teixeira et al. 2005a; Teixeira et al. 2005b; Teixeira et al. 2005c). HLB disease is transmitted among citrus trees by two psyllid vectors, Trioza erytrae (Del Guercio) in Africa and Diaphorina citri (Kuwayawa) in Asia (French et al. 2001; Boykin et al. 2007; Grafton-Cardwell et al. 2007). In Asia the proteobacteria causing HLB disease seem to be heat-tolerant as symptoms occur even at temperatures well above 30ºC. In Africa, the disease is prevalent only in cold areas where temperatures do not exceed 30ºC. The proteobacteria is thought to be heat-sensitive (Bové et al. 1974). For many years the distinction between the heat-tolerant and the heat-sensitive has been the only indication that two different strains of bacterium occurred, until Garnier et al. (1987), developed MA against “Fuzhou” strain from China and “Nelspruit” strain from South Africa. MAs against proteobacteria are highly specific and only react with the homologous MA serotypes of bacteria; however, the MAs are too specific for detection of all or most strains of the proteobacteria (Villechanoux et al. 1992). Villechanoux et al. (1992) first reported the use of DNA probes to detect Asia HLB causing bacterium strains. Hocquellet et al. (1997) developed radioactive probes and later non-radioactive probes for distinguishing between the Africa and Asiatic bacterium strains. Planet et al. (1995) cloned and sequenced the rplKAL-rpoBc operon of the African proteobacteria, which was used for characterization of the African strain of HLB causing bacterium. Teixeira et al. (2008) sequenced the β operon of the American liberibacter, and compared it with that of the African and Asian liberibacters. Other molecular studies on the proteobacteria have involved PCR detection based on the 16S ribosomal operon followed by digestion of the PCR products to distinguish between the African and Asiatic strain. Later an improvement on this protocol involved PCR-based detection of the ribosomal protein genes of the β operon without concomitant enzymatic digestion (Hocquellet et al. 1999a; Hocquellet et al. 1999b; Fang et al. 2004; Lan et al. 2004; Okuda et al. 2005; Li et al. 2007; Li et al. 2008; Tatineni et al. 2008). Bastianel et al. (2005) described an omp-based PCR-RFLP analysis method for detecting and differentiating ‘Candidatus Liberibacter asiaticus' isolates. Jagoueix et al. (1997) cloned and sequenced the intergenic spacer region 16S/23S rDNA of two strains of ‘Candidatus L. asiaticus' (Fuzhou from China), Poona from India and one strain Nelspruit from South Africa belonging to ‘Candidatus L. africanus'. These three strains from the two species belong to different serotypes and had 79.46% similarity in DNA of the intergenic spacers. The Asian strains were further characterized at the 16S and 16S/23S rDNA showing 85.5% to 100% similarity (Subandiyah et al. 2000; Fusayasu et al. 2006; Urasaki et al. 2007; Urasaki et al. 2008). It is difficult to get accurate assessments of yield losses from HLB disease in a perennial crop such as Citrus. However, infected branches produce little or no marketable fruit. Experiments in South Africa and reunion to rejuvenate HLB infected Citrus, using tetracycline injection, have reported increases in yields of up to 140%. In Kenya, yield losses of 100% have been reported and it is speculated that HLB disease causing proteobacteria in Kenya emanated from introduction of infected scions imported from South Africa since most citrus propagules were obtained from South Africa and to a limited extent from Israel. HLB disease was first diagnosed in Kenya using electron microscopy in 1982 and the disease led to the subsequent destruction of all sources of propagules from National Nurseries located in the highland areas. Although HLB disease has been in Kenya for the last 20 years, there is no information of what species/strain of the proteobacteria present. ‘Candidatus L. africanus' has been thought to be the cause of the disease based on the assumption that HLB disease in Kenya originated from South Africa. In a recent study (unpublished) we adopted the PCR diagnostic procedures developed by Hocquellet et al. (1999) to HLB samples from Kenya collected at different altitudes. In this study we report on DNA sequencing of the L10/L12 ribosomal protein genes and their intergenic region in HLB disease causing proteobacteria samples collected from infected citrus at different altitudes from three agro-ecological zones and define phylogenetic relationships among the proteobacteria. Materials and Methods DNA extraction and partial purification Leaf midribs (0.1 to 0.3 g) were chopped to a fine mince with a razor blade in a disposable petri-dish in extraction buffer containing 1 mL of TE buffer (10 mM Tris pH 8.0, 400 mM EDTA plus 1% SDS and 0.25 mg of proteinase K) (Jagoueix et al. 1996). The homogenate was transferred to an eppendorf tube and incubated for 2 hrs at 65ºC. The suspension was centrifuged for 15 min at 12,000 x g and the supernatant mixed with 1 mL of Wizard Min Prep DNA purification resin (Promega). The resin was transferred to a mini-column and washed twice with 2 mL of 80% isopropanol. Thereafter, 50 µL of hot water (80ºC) was added, incubated for 1 min and centrifuged at 16,000 x g for 30 sec in an eppendorf tube. This step was repeated to have a total yield of 100 µL of extract. The extract was stored at -20ºC. Amplification of the L10/L12 ribosomal protein rDNA Two primers were used to amplify the intergenic region of ribosomal protein L10/L12 rDNA and the L10 rDNA of the bacterium. Forward primer 5' (CATCGGGAGATGAAAGTTGAATA) and reverse primer 5' (TTCCCCTGCCGCAGACGCAACA). Amplification was done by PCR (Hybrid OMN-E Thermal Cycler) with the reaction conditions by Jagoueix et al. (1996). The PCR reaction was performed in 50 µL of reaction mixture containing 1 μm of each of the primers, Gibco buffer, 200 µM of each of the four dNTP, 2 mM MgCl2, 0.05% W1 detergent (Gibco BRL), 100 µg/mL BSA and 2.5 U of Taq polymerase (Gibco BRL). The reaction mix was amplified for 35 cycles with the thermal cycler at: 92ºC for 20 sec (denaturation of template into single strand); 62ºC for 20 sec (annealing of primers to each original strand, high temperatures are used due to the specificity of the primers); and 72ºC for 45 sec (elongation and new strand synthesis). PCR products were analyzed by electrophoresis of 20 µL of each sample (Jagoueix et al. 1996). Agarose gel electrophoresis and DNA elution from the gel PCR products were electrophoresed on 1% agarose gels and Tris-borate buffer (1 X TBE) and stained in ethidium bromide at 0.5 µg/mL. 20 µL of the PCR products were mixed with 1 µL of gel loading dye and loaded into gel. The gel was run at a voltage of 4 V/cm. After electrophoresis and photography, the DNA was extracted from the gel by use of a Gel Extraction Kit (Life Technologies). A gel slice containing the DNA was cut with a sharp razor under UV Light. The gel slice was weighed and 400 mg of the 1% gel was placed into a 1.5 mL propylene tube. Thereafter, 1.2 mL of gel solubilization buffer (concentrated sodium percholate, sodium acetate, and TBE solubilizer) was added. This was incubated at 50ºC for 15 min, during which the solution was mixed every 3 min to ensure gel dissolution. The DNA was recovered through a cartridge based protocol (Life Technologies). Automated fluorescent DNA sequencing/Sanger-Dideoxy Nucleoside Triphosphate Chain Termination DNA Sequencing method was used. Big Dye chemistry with ABI PRISM® BigDye™ Terminators v 3.0 and AmpliTaq® DNA polymerase Dyes attached to the four di-deoxynucleotide terminators were used for sequencing. The cycling reaction was done on a GeneAmp® PCR System with a MicroAmp® 96-Well Reaction Plate. The PCR fragments obtained were then separated on a denaturing polyacrylamide gel on Perkin Elmer ABI PRISM® 377 DNA sequencer. The dyes were detected by fluorescence after excitation. The sequence information was decoded by a 3100-Avant Genetic Analyzer soft ware. The PCR product was sequenced directly without cloning it in vectors. Purification of sequencing template This procedure was performed to remove primers and dNTPs, A G-50 w/spin column was rinsed by adding 300 µL H2O to spin column and span in a microfuge. 600 µL Sephadex G-50 slurry (1 g / 15 mL in ddH20) was added to the spin column and microfuge at 4900 RPM for 60 sec. 20 µL PCR products was loaded to the center of the Sephadex and microfuged at 4900 RPM for 60 sec and the product was transferred into fresh 1.5 mL eppendorf tube (QIAGEN Guide, 1998; Applied Biosystems, 2002). Extension/termination reactionsA reaction mixture consisting of template DNA 30-90 ng, primer 2.0 -3.0 pmol, buffer 4.0 µL, T and terminator Ready Reaction mix 2.0 µL was added into a 500 µL eppendorf tube and made to final volume of 20 µL with sterile H2O (Perkin-Elmer Corporation, 1997; Applied Biosystems, 2002). The reaction mixture was briefly mixed by pipetting. It was overlaid with 2 drops of mineral oil and spun down. The reaction tubes were placed into a thermalcycler (ThermoHybaid Omni-E) that had been preheated to 95ºC in a start the cycling program. The preheating was to prevent non-specifically annealed primers from being extended (and thus stabilized) by Amplitaq DNA polymerase. Thermalcycler was programmed to run at 95ºC for 1 min 30 sec preheating, then 95ºC for 30 sec, 59ºC for 15 sec (sequence-specific annealing temperature), 60ºC for 4 min and 25 cycles. The sample was stored at 4ºC at the end of the cycles. The PCR products were purified by running the products through Centri-Sep Columns to remove excess dye terminators (Perkin-Elmer Corporation, 1997; Applied Biosystems, 2002). 1.5 µL of the purified PCR products were loaded on 5.0% Long Ranger, 6 M-urea gel. DNA sequencing and homology comparisonsThe DNA was sequenced in a 48 cm ABI sequencing gel (5.0% long ranger acrylamide, 6 m urea). The gel consisted of 18.0 g urea, 5.0 mL 10 X TBE, 5.0 mL Long Ranger, 25.0 µL TEMED, 250.0 µL 10% APS, 27 mL ddH2O (up to 50 mL) (Perkin-Elmer Corporation, 1997; Applied Biosystems, 2002). After polymerization for 10-15 min the gel was positioned and warmed for 15 min to 51ºC. Samples were heated to 95ºC for 2 min and cooled on ice and 1.5 µL loaded in each well. Buffer was loaded into any empty wells to the right and left of the samples. The gel was run at a constant 48 watt for 8 hrs and the ABI base-caller results were received in ASCII format and were subjected to homology comparisons with other published sequences in the GenBank, EMBL, DDBJ, PDB Database using the BLAST (basic logical alignment sequencing tool) (Altschul et al. 1997), on the National Center for biotechnology information (NCBI) Genbank website. Phylogeny analysis for the L10/L12 rDNABootstrap test of phylogeny using the neighbourhood joining method and molecular evolutionary analyses were conducted using MEGA version 4 (Tamura et al. 2007). The L10/L12 rDNA sequences and the L10/L12 intergenic regions of ‘Candidatus Liberibacter' species from different agro-ecological zones in Kenya and those reported on the National Centre for Biotechnology Information (NCBI) Genbank were used in the analysis. The evolutionary distances were computed using the Maximum Composite Likelihood method (Tamura et al. 2004) and are in the units of the number of base substitutions per site. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 99 positions in the final dataset. Seven L10 rDNA, L10/L12 intergenic rDNA and partial L12rDNA sequences from 'Candidatus Liberibacter asiaticus' species were utilized in the analysis, they included isolate “Thai” (accession no. DQ852665), isolate “Japan” (accession no. AY342001), isolate “India” (accession no. AY266352), isolate “Pakistan” (accession no. EF493182), isolate “Asia” (accession no. M94319), isolate “China” (accession no. DQ157277), and isolate “Florida” from the USA (accession no. EU265648). Two sequences were from South Africa 'Candidatus Liberibacter africanus subsp. capensis' (accession no. AF248498) and 'Candidatus Liberibacter africanus str. Nelspruit' (accession no. U09675). Other Proteobacteria belonging to the α subdivision used included, Agrobacterium tumefaciens str. C58 (accession no. AEOO7869), Rickettsia rickettsii str. 'Sheila Smith' (accession no. CP000848), Brucella abortus (accession no. AF169147), and Bartonella tribocorum CIP105476 (accession no. AM260525). Escherichia coli O157.H7 str. Sakai (accession no. NC_002695) was used as an outgroup and belongs to the γ subdivision of Proteobacteria. Six isolates from Kenya (Kenya (lm3) isolate, Kenya (um3) isolate, Kenya (lm4) isolate, Kenya (lm5) isolate, Kenya (lh4) isolate and Kenya (um4) isolate were utilized in the analysis (Figure 1). Phylogeny analysis for the ribosomal protein L10The amino acid sequence of the L10 protein of 'Candidatus Liberibacter' species from Kenya and those reported were also used in determining phylogeny within the species. The phylogenetic history was inferred using the Minimum Evolution method with a bootstrap test (500 replicates). The evolutionary distances were computed using the Poisson correction method and the ME tree was searched using the Close-Neighbor-Interchange (CNI) algorithm (Nei and Kumar, 2000). The Neighbor-joining algorithm was used to generate the initial tree in MEGA4 (Tamura et al. 2007). Six L10 protein amino acid sequences from Kenyan isolates (Kenya (lm3) isolate, Kenya (um3) isolate, Kenya (lm4) isolate, Kenya (lm5) isolate, Kenya (lh4) isolate and Kenya (um4) isolate) were utilized in the analysis with L10 proteins from South African isolates 'Candidatus Liberibacter africanus subsp. capensis' (accession no. Q9L5W5) and 'Candidatus Liberibacter africanus str. Nelspruit' (accession no. P41191). The 'Candidatus Liberibacter asiaticus' isolates included isolate “Thai” (accession no. ABI35994), isolate “Japan” (accession no. AAR13469), isolate “India” (accession no. AAP22421), isolate “Asia” (accession no. P36249), isolate “China” (accession no. AAZ81564) and isolate “Florida” from the USA (accession no. ABY25857). Other Proteobacteria belonging to the α subdivision used included, Agrobacterium tumefaciens str. C58 (accession no. NP_354933), Rhizobium leguminosarum bv. viciae 3841 (accession no. YP_767368), Bartonella henselae str. Houston-1(accession no. YP_033437), Brucella abortus biovar 1 str. 9-941 (accession no.YP_221949), Ochrobactrum anthropi ATCC 49188 (accession no.YP_001370490). Escherichia coli O157.H7 str. Sakai (accession no. NP_312935) was used as an outgroup and belongs to the γ subdivision of Proteobacteria (Figure 2). Results and Discussion DNA sequences of L10 and L12 ribosomal protein genes A 7I6-basepair DNA fragment was amplified (Figure 3) and sequenced, producing 536 basepairs of DNA encoding the L10 protein, 44 basepairs of DNA intergenic region and 136 basepairs of DNA that partially encodes the L12 protein. Genes encoding the L10 protein of alpha subgroup proteobacteria have been sequenced and characterized in ‘Candidatus L. africanus strain Nelspruit' from South Africa (Planet et al. 1995), ‘Candidatus L. africanus subsp. capensis' (Garnier et al. 2000) in ‘Candidatus L. asiaticus' (Asiatic strains) (Villechanoux et al. 1993; Okuda et al. 2005) and in ‘Candidatus L. americanus' (Teixeira et al. 2008). The L10 and L12 protein are transcribed from the rplKAJL-rpoBC ribosomal protein-RNA polymerase gene cluster. The most abundant transcript is the 2600 nucleotide tetracistronic L11-L1-L10-L12 mRNA initiated at the upstream major PL11 promoter and terminated at the transcription attenuator in the L12-beta intergenic space (Petterson, 1979). Comparison DNA sequence of the rpL10 protein gene showed that the Kenyan isolates from different agro-ecological zones were 100% homologous to one another. In contrast the rpL10 DNA sequence from the Kenyan strains was 99%, 83% and 83.18% similar to ‘Candidatus L. africanus strain Nelspruit', ‘Candidatus L. africanus subspecies capensis' and ‘Candidatus L. asiaticus' strains, respectively (Table 1). Comparisons of the partial 136 base sequence of the rpL12 gene showed 100% homology among isolates from the Kenya's agro-ecological zones, but 99.26%, 91.9% and 84.56% similarity to the ‘Candidatus L. africanus strain Nelspruit', ‘Candidatus L. asiaticus' and ‘Candidatus L. africanus subsp. capensis' strains, respectively (Table 1). The low similarity between the Kenyan isolates and ‘Candidatus L. africanus subsp. capensis' as compared to ‘Candidatus L. asiaticus' may be attributed to the incomplete sequence of the rpL12 gene. However, the Kenyan strains of the alpha subgroup of HLB disease causing proteobacteria were different from the strains characterized from South African strains and the Asiatic strains, but more closely related to the former than the later based on the rpL10 and the rpL12 protein gene.
Translation and comparison of L10 ribosomal protein In the current study, DNA sequence of L10 gene was translated into a 169 amino acid protein sequence, with a molecular weight of 18637.6 and an atomic formula: C829H1388N232O248S2. It had an estimated half-life of 30 hrs and an instability index of 32.85, which classifies the protein as stable. It had a start codon of TTG at the seventeenth base while the L12 protein has its start codon at the 580th base in Kenyan isolate LM4 (Figure 4). In prokaryotes the ribosomal protein L10 is one of the proteins from the large ribosomal subunit that belongs to the L10 protein family of ribosomal proteins. L10 proteins range in size from 135 amino acids in proteobacteria Brucella abortus (Oliveira et al. 1994) to 199 amino acids in both aquificae Aquifex aeolicus (Deckert et al. 1998) and A. pyrophilus (Klent et al. 1999) and they have only been found so far in eubacteria. The ribosomal protein L8 is a complex L7/L12 and L10 (Petterson et al. 1976). The acidic stalk protein complex, L10.L7/L12 in prokaryotes, binds to a region around residues 1030-1127 in domain II of 23S rRNA, termed the GTPase-associated domain (Beauclerk et al. 1984), and constitutes a partof the functional center, termed the GTPase center or factor-bindingcenter (Ban et al. 1999; Ban et al. 2000). The four copies of L7/L12 bind to L10 and form the pentameric complex (Liljas and Gudkov, 1987); the L10 moiety of the complex appears to bind directly to the rRNA domain (Petterson, 1979). In the current study, the L10 amino acid sequence of Proteobacteria isolates from LM4 and LH4 agro-ecological zones were identical to each other and to ‘Candidatus L. africanus Nelspruit', but had 88.18% homology to the isolate ‘Candidatus L. africanus subsp. capensis' and 81.64% similarity to ‘Candidatus L. asiaticus' (Table 1). Previous comparisons of the species on the 16S/23S intergenic regions and the tRNAsAla sequence identified in the 16S/23S intergenic regions of ‘Candidatus L. africanus' and ‘Candidatus L. asiaticus' region were found to be 79.46% and 87.8% respectively (Jagoueix et al. 1997). Furthermore, the outer membrane protein (omp) of ‘Candidatus L. africanus' (isolate ‘South Africa-Nelspruit‘) and ‘Candidatus L. asiaticus' nucleotide sequences exhibited 72.2% identity, whereas the encoded polypeptides had 58% identical and 86.5% similar amino acids (Bastianel et al. 2005). Comparison of the β operon of liberibacters shows that ‘Ca. L. africanus'and ‘Ca. L. asiaticus' shared 81.2% sequence identity, the percentage for ‘Ca. L. americanus' and ‘Ca.L. africanus' was only 72.2%, and identity for ‘Ca.L. americanus' and ‘Ca. L. asiaticus' was only 71.4% (Teixeira et al. 2008). Beyond showing variation on the L10 protein sequence in the species, these results further indicate that the Kenyan strains of the proteobacterium were more closely related to ‘Candidatus L. africanus' than the ‘Candidatus L. asiaticus' in the amino acid sequences of the L10 and L12 ribosomal proteins. Intergenic region of L10/L12 ribosomal protein genes A 44-DNA base sequence comprising of the intergenic region of 'Candidatus L. africanus' strains from Kenya was obtained from UM3, UM4, and LM5, LM3 agro-ecological zones which were five bases shorter that the sequences obtained from ‘Candidatus L. asiaticus' and ‘Candidatus L. africanus subsp. capensis' and ‘Candidatus L. africanusstrain Nelspruit' (Figure 5). However, intergenic DNA sequences of Kenyan strain LH4 had an 11-basepair deletion compared to ‘Candidatus L. asiaticus' and ‘Candidatus L. africanus' strains. The intergenic regions from the LM4 isolates were 100%, 100%, 84.09%, 100%, 100%, 83.67%, 49% and 65.31% similar to isolates from LM3, LM5, LH4, UM3, UM4, ‘Candidatus L. africanus strain Nelspruit', ‘Candidatus L. asiaticus' and ‘Candidatus L. africanus subsp. capensis' strains, respectively (Table 1). The intergenic regions of ‘Candidatus L. africanus' strain from Kenya isolated from agro-ecological zones LM3, LM5, UM3 and UM4 were 100% homologous to one another. Similarly, the 595 basepair intergenic region of the 16S/23S of the ‘Candidatus L. asiaticus' strains from India and the People's Republic of China were 100% homologous to one another, although they were isolated distant geographically regions (Jagoueix et al. 1997). However, the level of homology of the entire 16S/23S intergenic regions of ‘Candidatus L. africanus' and ‘Candidatus L. asiaticus' was 87.46% and was only 498 base pairs long (Jagoueix et al. 1997). Because of the high homology between the ‘Candidatus L. asiaticus' strains from India and the People's Republic of China even though the two strains belonged two different serotypes (Jagoueix et al. 1997), concluded that probably in ‘Candidatus Liberibacter' the 16S/23S intergenic region does not vary much within a species. Hence monoclonal antibodies remain the only reagents that allow identification of ‘Candidatus Liberibacter' strains within a given species. However in the current study, the variability in the sequences of ‘Candidatus L. africanusstrain Nelspruit' were high, 49%, 65.31%, and 83.67% homology respectively, to ‘Candidatus L. asiaticus', ‘Candidatus L. africanus subsp. capensis' and ‘Candidatus L. africanus'strain from Kenya isolated from LM and UM. Results from the current study show that variability within the L10/L12 intergenic regions was high and may be used as a means of distinguishing between strains in the alpha subgroup of HLB disease causing proteobacteria. In addition the results from comparison of DNA sequences from the L10/L12 intergenic region further corroborate that the Kenyan strains of the proteobacteria are different from the South African strains and the Asiatic strains but more closely related to the former than the later. Phylogeny of the ‘Candidatus Liberibacter' species The ribosomal DNA sequence and the intergenic rDNA sequences have been previously used to study phylogeny in Proteobacteria (Hugenholtz, 2002), among the 21 Phloem-restricted, uncultured walled bacteria two proteobacteria have been phylogenetically characterized; the citrus huanglongbing (greening) (Jagoueix et al. 1994) and the strawberry chlorosis bacterium (Zreik et al. 1998). Both have been phylogenetically characterized based on the polymorphism within the rDNA sequences. The L10/L12 intergenic rDNA sequences were used to cluster the ‘Candidatus Liberibacter' species isolated in Kenya as compared to those posted in the genebank (Figure 1). The Kenyan strains of the ‘Candidatus Liberibacter' species clustered within the alpha (α) subdivision of proteobacteria. The Kenyan strains clustered away from the other African strains but with low bootstrap values of 19 (Figure 1), showing that the difference between the African strains is not wide. The Kenyan strains formed a cluster with the South African isolates ‘Capensis' (Garnier et al. 2000) and ‘Nelspruit' (Planet et al. 1995) with bootstrap values of 76 (Figure 1). This showed that the Kenyan strains are more closely related to the South African strains than the Asian strains. However the Asian isolates (‘Thai', ‘India', ‘China', ‘Japan', ‘Florida', ‘Asia' (Villechanoux et al. 1993), and ‘Pakistan‘ (Chohan et al. 2007) clustered within the α subdivision. Other members of the α subdivision including; Agrobacterium tumefaciens str. C58 (Wood et al. 2001), Rickettsia rickettsii str. 'Sheila Smith', Brucella abortus (Campos et al. 1998), and Bartonella tribocorum CIP105476 (Saenz et al. 2007), clustered with all α-2 proteobacteria away from Escherichia coli O157.H7 str. Sakai (Makino et al. 1999; Yokoyama et al. 2000; Hayashi et al. 2001) that represented the gamma (γ) subdivision of proteobacteria. Ribosomal protein sequences have been utilized for phylogenetic analysis in eubacteria, due to their evolutionary important structural and functional properties and the existence of conserved domains (Müller and Wittmann-Liebold, 1997). The nusG-rplKAJL-rpoBC gene cluster has been previously used to study phylogenetic relationships in Liberibacters (Villechanoux et al. 1993). The L10 amino acid sequence phylogeny shows the Kenyan isolates (UM3, UM4, LM5, LM4, LM3, and LH4) clustering with other α proteobacteria. The Kenyan strains and the South African strain ‘Nelspruit' (Planet et al. 1995), form a distinct cluster with high bootsrap values away from the South African isolate ‘Capensis' (Garnier et al. 2000) (Figure 2). However, the Kenyan and the South African isolates share a common origin more recent than the one they share with the Asian isolates (‘Thai', ‘Japan', ‘India', ‘Asia', ‘China' and ‘Florida') denoting the close relationship between the Kenyan and South African isolates (Figure 2). Other members of the α subdivision of Proteobacteria Agrobacterium tumefaciens str. C58 (Goodner et al. 2001; Wood et al. 2001), Rhizobium leguminosarum bv. viciae 3841 (Young et al. 2006), Bartonella henselae str. Houston-1 (Alsmark et al. 2004), Brucella abortus biovar 1 str. 9-941 (Halling et al. 2005), Ochrobactrum anthropi ATCC 49188. Escherichia coli O157.H7 str. Sakai (Yokoyama et al. 2000; Hayashi et al. 2001), diverged from the Liberibacters earlier and forms a cluster with a bootstrap value of 93. The L10/L12 rDNA and the L10 protein phylogenetic clusters show that the Kenyan isolates and other Liberibacters are members of the α subdivision of proteobacteria. This is consistent with findings using the omp gene (Bastianel et al. 2005), the 16S rRNA (Jagoueix et al. 1994, Teixeira et al. 2005b), the 16S rDNA (Garnier et al. 2000) and the 16S/23S rDNA (Jagoueix et al. 1997). Therefore, the L10/L12 intergenic rDNA and the L10 protein serves as a good tool for phylogeny analysis among ‘Candidatus Liberibacter' species, however, the region cannot be used to cluster within a species as was reported for the 16S rDNA and 16/23S rDNA. (Jagoueix et al. 1997; Teixeira et al. 2005b). These results suggest that, the ‘Candidatus Liberibacters' active in the country maybe South African in origin. Concluding Remarks The isolated HLB causing proteobacteria in Kenya was more closely related to the ‘Candidatus L. africanus' strains from South Africa than the ‘Candidatus L. asiaticus' strains isolates from Asia with respect to the L10 and L12 rDNA and amino acid sequences and the L10/L12 intergenic DNA sequences. There was high homology in L10 and L12 rDNA and amino acid sequences and the L10/L12 intergenic DNA sequences of alpha proteobacteria isolated from different agro-ecological zones in Kenya. Phylogeny analysis on the L10/L12 rDNA and the L10 amino acid sequence shows significant clusters within the ‘Candidatus Liberibacter' species with the Kenyan strains being closely related to the South African strains. The HLB disease causing bacteria in Kenya is distinct from the South African but they have a common origin more recent than the one they share with the Asian HLB causing bacteria. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2009 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej09012f4.jpg] [ej09012f3.jpg] [ej09012f2.jpg] [ej09012f1.jpg] [ej09012f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}