
|
Electronic Journal of Biotechnology
Universidad Católica de Valparaíso
ISSN: 0717-3458
Vol. 12, Num. 2, 2009
|
Electronic Journal of Biotechnology, Vol. 12, No. 2, April 15, 2009
Review Article
AgNO3 - a potential regulator of ethylene activity and
plant growth modulator
Vinod Kumar1 , Giridhar Parvatam2, Gokare Aswathanarayana
Ravishankar*3
1Plant Cell Biotechnology
Department,
Central Food Technological
Research Institute,
Mysore-570 020,
Karnataka State, India
2Plant Cell Biotechnology
Department,
Central Food Technological
Research Institute,
Mysore-570 020,
Karnataka State, India
3Plant Cell Biotechnology
Department,
Central Food Technological
Research Institute,
Mysore-570 020,
Karnataka State, India
Tel: 91 821 2516501
Fax: 91 821 2517 233
E-mail: pcbt@cftri.res.in
*Corresponding author
Financial support: Department of Biotechnology, Government of India.
Received March 6, 2006
/ Accepted August 7,2008
Code Number: ej09015
Abstract
The aim of
this review is to critically analyze the role of silver nitrate (AgNO3)
in modulating plant growth and development. In recent years, basic studies on
ethylene regulation opened new vistas for applied research in the area of micro-propagation,
somatic embryogenesis, in vitro flowering, growth promotion, fruit
ripening, and sex expression. Silver nitrate has proved to be a very potent
inhibitor of ethylene action and is widely used in plant tissue culture. Few
properties of silver nitrate such as easy availability, solubility in water,
specificity and stability make it very useful for various applications in
exploiting plant growth regulation and morphogenesis in vivo and in
vitro. Silver ion mediated responses seem to be involved in polyamines,
ethylene- and calcium- mediated pathways, and play a crucial role in regulating
physiological process including morphogenesis. The molecular basis for
regulation of morphogenesis under the influence of silver nitrate is completely
lacking. This review compiles published reports of silver nitrate-mediated in
vitro and in vivo studies and focuses on fundamental and applied
aspects of plant growth modulation under the influence of silver nitrate.
Keywords: calcium, ethylene, morphogenesis, polyamines, silver nitrate, somatic
embryogenesis.
Abbreviations: |
ACC: 1-Amino-cyclopropane-1-carboxylic
acid
ADC: arginine decarboxylase
AgNO3: silver nitrate
AVG: aminoethoxyvinylglycine
BA: 6-benzylaminopurine
DFMA: α-DL-difluromethyl arginine
DFMO: α-DL-difluromethyl ornithine
IAA: indole acetic acid
NAA: α-naphthalene acetic acid
ODC: ornithine decarboxylase
PA: polyamine
Put: putrescine
SAM: S-adenosyl-L- methionine
STS: silver thiosulphate
Spd: spermidine
Spm: spermine |
In recent
years, advances in plant genetic engineering have opened new avenues for crop
improvement and various plants with novel agronomic traits have been produced.
The success of plant genetic engineering relies on several factors which
include an efficient tissue culture system, for regeneration of plants from
cultured cells and tissues (Pua et al. 1996). Shoot generation and rooting are
important in the realization of the potential of the cell and tissue culture
techniques for plant improvement (Purnhauser et al. 1987). Silver ions in the
form of nitrate, such as AgNO3, play a major role in influencing
somatic embryogenesis, shoot formation and efficient root formation which are
the prerequisites for successful genetic transformation (Bais et al. 1999; Bais et al. 2000a; Bais et al. 2000b; Bais et al.
2001a; Bais et al. 2001b; Bais et al. 2001c). Silver ions are also
employed in the form of silver thiosulphate in several tissue culture studies (Eapen
and George, 1997).
Ethylene
is recognized as a ubiquitous plant hormone (Lieberman,
1979; Yang,
1985), which influences growth
and development of plants (Abeles, 1973; Yang
and Hoffman, 1984; Mattoo and
Suttle, 1991). In vitro studies have indicated that ethylene
can affect callus growth, shoot regeneration and somatic embryogenesis in
vitro (Purnhauser et al. 1987; Songstad
et al. 1988; Roustan et al. 1989; Roustan
et al. 1990; Biddington, 1992; Pua
and Chi, 1993). Thus, by regulating the
production or action of ethylene, the growth and development of some
tissue cultures can be controlled to a certain extent (Beyer,
1976c; Davies, 1987; Purnhauser
et al. 1987; Songstad
et al. 1988; Chi and Pua, 1989; Bais
et al. 2000a; Giridhar et al. 2003).
AgNO3 has
been known to inhibit ethylene action (Beyer, 1976a)
and cobaltous ions are known to inhibit ethylene synthesis (Lau
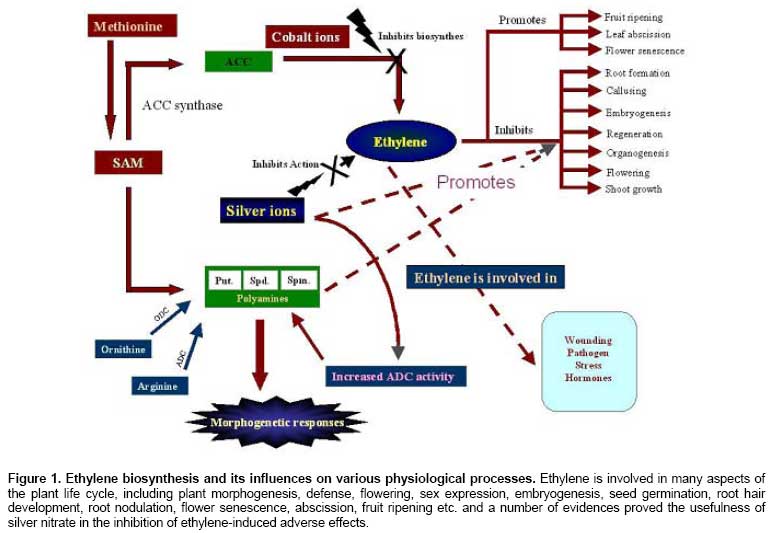
and Yang, 1976) (Figure 1). Silver ion
is capable of specifically blocking the action of exogenously applied
ethylene in classical responses such as abscission, senescence
and growth retardation (Beyer, 1976c). These
observations led to its application in tissue culture. Addition
of AgNO3 to the culture media greatly improved the
regeneration of both dicot and monocot plant tissue cultures (Beyer,
1976c; Duncan et al. 1985; Davies, 1987; Purnhauser
et al. 1987; Songstad et al.
1988; Chi and
Pua, 1989; Veen
and Over Beek, 1989; Bais
et al. 2000a; Giridhar et al. 2003). The exact mechanism of AgNO3 action
on plants is unclear. However, few existing evidences suggest its
interference in ethylene perception mechanism (Beyer,
1976c). In recent years, AgNO3 has been employed in
tissue culture studies for inhibiting ethylene action because of
its water solubility and lack of phytotoxicity at effective concentrations
(Beyer, 1976a).
Ethylene
Ethylene
is a gaseous plant hormone involved in many aspects of plant life
cycle (Figure 1) such as seed
germination, root hair development, root nodulation, flower senescence,
abscission, and fruit ripening (Johnson and Ecker,
1998; Bleecker and Kende,
2000). Its biosynthesis (Wang et al. 2002)
is tightly regulated by internal signals and environmental stimuli
from biotic and abiotic stresses, such as pathogen attack, wounding,
hypoxia, ozone, chilling, or freezing (Wang et al.
2002). The role of ethylene in morphogenesis has been well documented
in an earlier review (Kumar et al. 1998a). Mutants
have also been identified that display a constitutive triple response
in the absence of ethylene (Kieber et al.
1993). This can be divided into subgroups based on whether or
not the constitutive triple response can be suppressed by inhibitors
of ethylene perception and biosynthesis, such as silver ions and
aminoethoxyvinyl glycine (AVG). Mutants that are unaffected by these
inhibitors are termed constitutive triple-response (ctr)
mutants, whereas mutants whose phenotype reverts to normal morphology
are termed ethylene-overproducer (eto) mutants, which are defective
in the regulation of hormone biosynthesis. To date, data is lacking
on the molecular basis for silver ion interaction with the mutants,
which are insensitive to ethylene.
Ethylene biosynthesis
To understand
the role of silver ions in regulating morphogenesis, it is important to know
the aspects of ethylene biosynthesis (Figure 1). The biochemistry of
ethylene biosynthesis has been a subject of intensive study in plant hormone
physiology (reviewed by Wang et al. 2002). In brief, the biosynthesis of
ethylene starts with conversion of the amino acid methionine to S-adenosyl-L-
methionine (SAM, also called Adomet) by the enzyme Met Adenosyltransferase. SAM
is subsequently converted to 1-aminocyclopropane-1-carboxylic-acid (ACC) by the
enzyme ACC synthase (ACS). The activity of ACS is the rate-limiting step in
ethylene synthesis. The final step requires oxygen and involves the action of
the enzyme ACC-oxidase (ACO), formerly known as the ethylene forming enzyme
(EFE) (Wang et al. 2002).
A major
breakthrough in the ethylene synthesis pathway was the establishment of
S-adenosylmethionine (S-AdoMet) and ACC as the precursors of ethylene (reviewed
in Yang and Hoffman, 1984; Kende, 1993). On the basis of this knowledge, the
enzymes that catalyze these reactions were characterized and purified. The
first successes in molecular cloning of the ACC (Sato and Theologis, 1989) and
ACO (Hamilton et al. 1991; Spanu et al. 1991) genes led to the demonstration of
these enzymes belonging to a multi-gene family and are regulated by a complex
network of developmental and environmental signals responding to both internal
and external stimuli (reviewed by Johnson and Ecker, 1998). In addition to
being an essential building block of protein synthesis, nearly 80% of cellular
methionine is converted to S-AdoMet by S-AdoMet synthetase (SAM synthetase) at
the expense of ATP utilization (Ravanel et al. 1998). S-AdoMet is the major
methyl donor in plants and is used as a substrate for many biochemical
pathways, including polyamines and ethylene biosynthesis (Ravanel et al. 1998).
Ethylene
signal perception
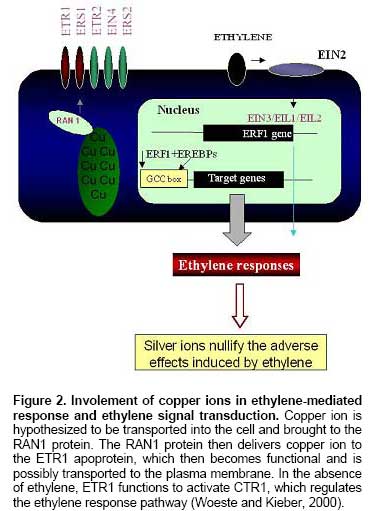
Ethylene is
perceived by a family of five membrane-localized receptors that are homologous
to bacterial histidine kinases involved in sensing environmental changes (Figure 2). Ethylene binding occurs at the N-terminal transmembrane domain of the
receptors, and a copper co-factor is required for the binding. The system
typically consists of a histidine kinase as the sensor that autophosphorylates
an internal histidine residue in response to environmental signals, and a
response regulator that activates the downstream components upon receiving a
phosphate from the histidine residue of the sensor on its aspartate residue (Wurgler-Murphy
and Saito, 1997). Five ethylene receptors exist in Arabiodpsis: ETR1,
ETR2, ERS1, ERS2, and EIN4 (Chang et al. 1993; Hua et al. 1995; Hua and
Meyerowitz, 1998; Sakai et al. 1998). Further characterization of ethylene
binding to ETR1 has revealed that it occurs at the hydrophobic pocket located
at the N- terminus of the receptors and requires a transition metal, copper, as
a co-factor (Figure 2) (Schaller and Bleecker, 1995; Rodriguez et al.
1999; Wang et al. 2002). Further findings indicated that
RAN1 is involved in the delivery of copper to the ethylene receptor and
that this copper-delivery pathway is required to create functional ethylene
receptors in plants (Figure 2) (Wang et al. 2002). Cu ions are also
known to form complexes with ethylene (Coates et al. 1968). But the studies of Beyer (1976c) revealed that the
effect of silver ions could be explained on the basis that silver ions
substitute for Cu ions, thereby interfering with ethylene action. This may be
due to the similarity in size, the same oxidation state, and the ability of
both Cu ion and Ag ion to form complexes with ethylene (Coates et al. 1968).
The possibility of the anti-ethylene property of silver was later well explored
in various plant systems. At present there are no concrete evidences to show
the involvement of silver ions with signaling networks which leads to down
regulation of physiological responses governed by ethylene. Therefore, focus on
the elucidation of molecular basis for diverse developmental process in plants
such as abscission, flowering, fruit ripening, morphogenesis and sex
expression, that are known to be regulated by silver ions, would be
interesting.
Possible
mechanisms of action of silver nitrate on ethylene action inhibition
Silver ions are capable
of generating ethylene insensitivity in plants (Zhao et al. 2002).
Ethylene-insensitive mutations (Hall et al. 1999) and silver ions are thought
to perturb the ethylene binding sites (Rodriguez et al. 1999). The ethylene
receptor, ETR1, contains one ethylene-binding site per homodimer and binding is
mediated by a single copper ion (Cu) present in the ethylene-binding site. The
replacement of the copper co-factor by silver also serves to lock the receptor
into a conformation such that it continuously represses ethylene responses
(Zhao et al. 2002).
There are
different views and experimental evidences on this subject. According to one
view, the ethylene action in plants is inhibited by week antagonists such as CO2 and strong antagonists like silver compounds. This is possibly due to oxidation
of ethylene by a metal-ion enzyme system (Abeles, 1973). In Arabidopsis,
insensitivity to ethylene is conferred by dominant mutation in receptors
(Bleecker et al. 1988). Another hypothesis is that AgNO3 inhibits
ethylene action by means of silver ions by reducing the receptor capacity to
bind ethylene (Yang, 1985), which would result in higher titers of ethylene in
the tissues, thus inhibiting the earlier steps of its own pathway. Miyazaki and
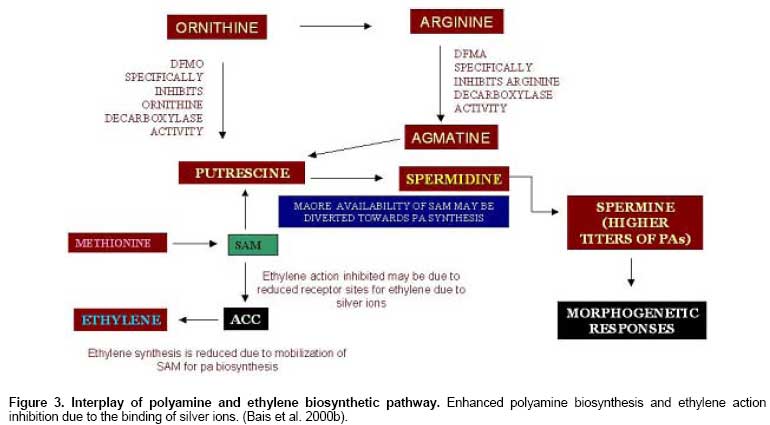
Yang (1987) reported the influence of putrescine and AgNO3 on the
competitive utilization of SAM. Bais et al. (2000b) also postulated that the
utilization of SAM by putrescine for its conversion to spermidine would
possibly result in a lower availability of SAM for ethylene biosynthesis (Figure 3). The introduction of ethylene antagonists into the culture media affects
the level of ACC, thereby affecting ethylene levels (Gong et al. 2005).
Polyamines
Other
important substances responsible for regulation of morphogenesis
are polyamines. The polyamines (PAs) are organic compounds having
two or more primary amino groups. Polyamines have been implicated
in several important cellular processes like cell division, morphogenesis,
protein synthesis, DNA replication, and plant response to abiotic
stress (Tabor and Tabor, 1984; Smith,
1985; Smith, 1993; Van Den
Broeck et al. 1994; Walden
et al. 1997; Kumar and Rajam, 2004). They bind
to DNA, and are essential for cell viability (Flink
and Pettijohn, 1975). Polyamines
are also known to be involved in DNA helix stabilization, stabilization
of loops in RNA molecules, membrane permeability, DNA replication,
cell division, gene expression, regulation of enzyme activities,
membrane stabilization, morphogenesis, fruit ripening etc. (Bais
and Ravishankar, 2002; Kumar and
Rajam, 2004). It has been postulated that polyamines and related
compounds are a type of growth regulator or secondary hormonal messenger
(Galston, 1983; Davies, 1987). PAs are found in plant cells at
significantly higher levels than plant hormones. There is evidence
that PAs are taken up by cell suspension cultures (Evans
and Malmberg, 1989). Interestingly, it seems that there is a
strong link between ethylene, polyamines, and calcium-mediated signaling.
This triangle is expected to be a potential target for silver ions.
This is because both ethylene and polyamines are metabolically related
(Figure 3)
and utilize the same precursor, SAM, for their synthesis (Evans
and Malmberg, 1989; Bais and Ravishankar, 2002).
It has also been suggested that polyamines and ethylene may regulate
each other’s synthesis. For instance, ethylene has been shown
to inhibit arginine decarboxylase and S-adenosyl methionine
decarboxylase activities in pea seedlings (Apelbaum
et al. 1985). These enzymes are necessary
for polyamine synthesis (Smith, 1985). It has
been proved beyond doubt that polyamines play crucial roles in plant
growth and development as well as basic biological process (reviewed
by Kumar and Rajam, 2004). Since polyamines have
been reported to promote embryogenesis (Feirer et al.
1984), the promotive
effect of ethylene inhibitors, such as AgNO3, on regeneration
was thought to be due to enhanced polyamine synthesis rather than
reduced ethylene production. Pua et al. (1996) clearly
described the synergistic effect of AgNO3 and putrescine
on shoot regeneration in Chinese radish. Miyazaki and
Yang (1987) reported the influence of putrescine and AgNO3 on
the competitive utilization of SAM. Bais et al. (2000b) postulated
that, utilization of SAM by putrescene for its conversion to spermidine
would possibly result in a lower availability of SAM for ethylene
biosynthesis (Figure 3). On the other hand, Pua
and Chi (1993) also
reported the same stimulatory effect of AgNO3 feeding
on ethylene production and its contribution to increased titers of
polyamines in mustard. Polyamines also regulate the growth and secondary
metabolism (Bais et al. 1999; Bais et al. 2001b; Bais and
Ravishankar, 2002). Reports on somatic embryogenesis in carrot
(Roustan et al. 1990; Nissen, 1994) indicate that the potent ethylene
action inhibitor, AgNO3,
causes the increase of ADC activity, which in turn increases the
levels of endogenous polyamines in carrot embryogenic cultures.
Involvement
of calcium in polyamine-mediated response
Polyamines
are associated with Ca2+ ions in signaling events (Majewska-Sawka
et al. 1998). They supported the hypothesis of transportation
of spermidine/spermine within protoplasts through a carrier-mediated
mechanism (Antognoni et al. 1994, Majewska-Sawka
et al. 1997). Majewska-Sawka et al.
(1998) found that spermidine/spermine may result in change in
distribution of Ca2+ ions. It is reasonable to conclude
that Ca2+ ions may be involved in the
mechanism of polyamine action in plant cells (Bush,
1995). This aspect is very
relevant here because, apart from ethylene regulation, silver nitrate
is known to regulate the polyamine pool in plant systems.
Application
of silver nitrate in plant tissue culture
So far we have
discussed the possible mechanisms of regulation of morphogenesis by silver
nitrate. Interestingly, a large number of reports are accumulating on the
utility of silver nitrate in tissue culture and other applications, with
significant contributions towards the development of plant biotechnology and
transgenic research. The following section deals with a brief compilation of
published research pertaining to the effect of silver nitrate in plant
morphogenesis (Table 1).
Table
1. Composition of medium in transformation experiment. |
Plant |
Response |
Reference |
Albizzia julibrissin |
In vitro shoot formation |
Sankhla et al. 1995 |
Andrographis paniculata |
Somatic embryogenesis |
Martin, 2004 |
Apple |
Higher efficiency of regeneration and transformation |
Seong et al. 2005 |
Arachis hypogea |
Regeneration |
Pestana, 1999 |
Arachis hypogea |
Multiple shoot formation |
Ozudogru et al. 2005 |
Albizzia procera |
Plant regeneration |
Kumar et al. 1998b |
Bactris gasipaes |
Enhance embryogenic competence |
Steinmacher
et al. 2007 |
Brassica campastris |
Shoot regeneration |
Palmer, 1992 |
Brassica juncea |
Microspore embryogenesis |
Prem et al. 2005 |
Capsicum annuum |
Shoot development and plant regeneration |
Hyde and Phillips, 1996 |
Cicer arietinum |
Somatic embryo induction |
Patil et al. 1999 |
Cichorium intybus |
Shoot length, shoot number, flowering |
Bais
et al. 2001a; Bais et al. 2001b; Bais et al.
2001c |
Coffea arabica |
Shoot growth
Somatic embryogenesis
Direct somatic embryogenesis |
Ganesh
and Sreenath, 1996
Giridhar et al. 2003
Giridhar et al. 2004 |
Coffea canephora |
Shoot growth
Somatic embryogenesis
Direct somatic embryogenesis |
Giridhar et al. 2003
Giridhar
et al. 2004; Kumar et al. 2007
Fuentes et al. 2000
|
Cucumis sativus |
Shoot regeneration
Sex expression |
Mohiuddin et al. 1997
Atsmon and Tabbak, 1979 |
Daucus carota |
Somatic embryogenesis |
Nissen et al. 1994 |
Decalepis hamiltonii |
In vitro root formation and shoot formation |
Bais
et al. 2000a; Bais et al. 2000b; Reddy et al.
2001 |
Egyptian maize |
Enhanced the formation of embryogenic type II callus |
El-Itriby et al. 2003 |
Eleusine coracana |
Plant regeration |
Kothari-Chajer et al. 2008 |
Glycine max |
Shoot formation on hypocotyls |
Wang and Xu, 2008 |
Gossypium sp. |
Leaf abscission
Enhance multiple shoot production from hypocotyl
segments |
Beyer,
1976a
Divya et al. 2008 |
Helianthus annuus |
Regeneration and shoot organogenesis |
Chraibi et al. 1991 |
Hordium vulgare |
Somatic embryogenesis
Plant regeneration |
Castillo
et al. 1998
Jha et al. 2007 |
Ipomoea batatas |
Shoot
regeneration |
Gong
et al. 2005 |
Manihot esculenta |
Shoot
organogenesis |
Zang et al. 2001 |
Morus Alba |
Modification of sex expression |
Thomas, 2004 |
Nicotiana plumbaginifolia |
Shoot regeneration |
Purnahauser et al. 1987 |
Oryza sativa |
Androgenesis |
Lentini et al. 1995 |
Paspalum scrobiculatum |
Plant regenerarion |
Kothari-Chajer et al. 2008 |
Passion fruit |
Regeneration |
Reis et al. 2003 |
Penisittum glaucum |
Plant regeneration |
Oldach et al. 2001 |
Penisittum glaucum |
Regeneration from zygotic embryos |
O'Kennedy et al. 2004 |
Pennisettum americanum |
Plant regeneration |
Plus et al. 1993 |
Phaseolus vulgaris |
Shoot development |
Cruz de Carvalho et al. 2000 |
Phoenix dactylifera |
Somatic embryogenesis |
Al-Khayri
and Al-Bahrany, 2001; Al-Khayri and
Al-Bahrany, 2004 |
Picea glauca |
Somatic embryogenesis |
Kong and Yeung, 1994 |
Picea glauca |
Maturation of somatic embryos |
El Meskaoui et al. 2000 |
Pisum sativum |
Shoot growth |
Beyer, 1975 |
Punica granatum |
Adventitious shoot regeneration. |
Naik and Chand, 2003 |
Quassia amara |
somatic embryogenesis |
Martin and Madassery, 2005 |
Raphanus sativus |
Shoot regeneration |
Pua et al. 1996 |
Rubus sp |
Inhibits callous formation during shoot
multiplication |
Tsao and Reed, 2002 |
Sorghum bicolor |
Plant regeneration |
Oldach et al. 2001 |
Stenotaphrum secundatum |
Embryogenic callus shoot regeneration. |
Fei et al. 2000 |
Tagetes erecta |
Plant growth flowering and seed viability |
|
Triticum aestivum |
Plant regeneration from callus
Improved embryogenic callus frequency |
Yu et al. 2008
Wu et al. 2006 |
Vanilla planifolia |
Shoot growth and in vitro root formation |
Giridhar et al. 2001 |
Vicia faba |
Promoted root formation |
Khalafalla and Hattori, 2000 |
Vigna ungiculata |
In vitro regeneration |
Brar
et al. 1999 |
Zea mays |
Regenerable
callus
Embryogenic
calli from embryo scutellum |
Songstad
et al. 1988
Valdez-Ortiz,
2007 |
Somatic
embryogenesis
Theoretically,
each living plant cell is capable of forming somatic embryos. Somatic
embryos are formed from vegetative plant cells. Applications of
this process include: clonal propagation of genetically uniform
plant material, elimination of viruses, provision of source tissue
for genetic transformation, generation of whole plants from single
cells called protoplasts, and development of synthetic seed technology.
Plant growth regulators in the tissue culture medium can be manipulated
to induce callus formation and subsequently changed to induce embryos
from the callus or directly from intact tissues. The ratio of different
plant growth regulators required to induce callus or embryo formation
varies with the type of plant. The use of silver nitrate
improved somatic embryogenesis in several plant species such as buffalograss
(Fei
et al. 2000), Coffea sp. (Fuentes et al.
2000; Giridhar
et al. 2004),
carrot (Nissen, 1994), white spruce (Kong
and Yeung, 1994), Triticum durum (Fernandez
et al. 1999), and Zea mays (Vain Hort and
Flament, 1989; Vain
Hort et al. 1989; Songstad et al. 1991).
Multiple
shoot induction and shoot regeneration
Silver
nitrate is known to promote multiple shoot formation in different
plants. In vitro shoot formation was improved by incorporating
silver nitrate in the culture medium. Ganesh and
Sreenath (1996) reported in vitro sprouting of apical
buds of Coffea under the influence of AgNO3. The
addition of N6-benzyladenine with AgNO3 greatly enhanced
the rate of sprouting. At low concentration, AgNO3 was
found to cause delayed senescence resulting in improved growth of
the proliferated shoots in Coffea canephora (Fuentes
et al. 2000). AgNO3 enhanced in vitro shoot
growth of C. arabica and C. canephora (Giridhar
et al. 2003). Shoot
regeneration of Chinese radish Cv Red coat was improved when
cultured in media supplemented with 2030 µM AgNO3 (Pua
et al. 1996). Brassica sp. are poorly responsive to tissue
culture manipulations (Narasimhulu
and Chopra, 1988). B. campestris produces high levels
of ethylene causing abnormal growth and development of the plant
in tissue culture conditions (Lentini et al. 1988),
and also inhibits de novo shoot
regeneration in vitro (Chi et al. 1990; Chi
et al. 1991; Palmer, 1992; Pua and Chi, 1993). The cotyledons and hypocotyls
of 7 cultivars belonging to B.
campestris spp. chinensis, spp. pekinensis and spp. parachinensis
exhibited improved shoot regeneration on culture media supplemented
with growth regulators and AgNO3.
The effects of the ethylene precursor,
1-aminocyclopropane-1-carboxylic acid (ACC), and two inhibitors, silver
thiosulfate and aminoethoxyvinylglycine (AVG), were tested in yellow passionfruit (Passiflora
edulis) axillary buds cultured in vitro (Reis et al. 2003). The
organogenesis was assessed by the number of buds per explant, mean leaf area
per explant, and shoot length. ACC-supplemented medium significantly inhibited
all evaluated responses. When ethylene action and biosynthesis were inhibited,
a significant enhancement of buds and leaf area was observed. The results
suggest beneficial effects of silver nitrate on in vitro development of
axillary buds.
Inhibition of ethylene action by
AgNO3 stimulated regeneration of shoots from cotyledon explants of Helianthus
annus (Chraibi et al. 1991). In many plants, the regeneration potential of
cultured cells and tissues decreases with increased cycles of subcultures
(Ogura and Shimada, 1978; Vasil, 1987). This phenomenon is evident in Pennisetum
americanum (Pearl millet) and Plus et al. (1993) effectively addressed this
issue by incorporating AgNO3 in the culture medium to restore the
regeneration potential. AgNO3 enhanced shoot regeneration frequency
in silk tree (Albizzia julibrissin) and Nicotiana plumbaginifolia.
The
work done in our laboratory has shown that exogenous feeding of
putrescine and silver nitrate influenced morphogenesis in chicory
(Chichorium intybus) shoot
cultures (Bais et al. 2000b). Putrescine and AgNO3 induced
shoot multiplication and in vitro flowering. The chicory plants,
which flower biennially, could be forced to flower experimentally
for studies on in
vitro pollination and seed development (Bais et
al. 2000b).
Silver
nitrate was found to be beneficial in the regeneration and clonal
propagation of several economically important plants (Table
1) such as peanut (Pestana et al. 1999),
cowpea (Brar et
al. 1999), Brassica sp. (Eapen and George,
1997; Pua et al. 1999), Capscicum sp.
(Hyde and Philips, 1996, Kumar et
al. 2003a), watermelon (Lim
and Song, 1993), Coffea canephora (Kumar
et al. 2003b), Cucumber (Mohiuddin et al. 1997),
Pomegranate (Naik and Chand, 2003), White marrygold
(Misra and Datta, 2001),
Cassava (Zang et al. 2001), Petunia (Gavinlertvatana
et al. 1980), etc.
In vitro rooting
Decalepis
hamiltonii Wight, Arn (swallow root), belonging to
Asclepiadaceae, is a monogeneric climbing shrub, native of the Deccan peninsula
and forest areas of Western Ghats in India. It is used as a culinary spice due
to its aromatic roots. In vitro root formation is a major issue in the
tissue culture of this plant. Effects of AgNO3 on in vitro root formation of Decalepis hamiltonii were studied. Addition of 40 µM AgNO3 resulted in root
initiation and elongation (Bais et al. 2000a; Reddy
et al. 2001).
Vanilla
is an important spice crop of commercial value. The effect of AgNO3 on
rooting and shooting was elucidated in Vanilla
planifolia (Giridhar et al. 2001). Maximum
number of shoots and highest shoot length was obtained on medium
containing 20 µM AgNO3. AgNO3 not only
induced shoot multiplication but also influenced rooting of vanilla
explants. The plantlets obtained on medium containing 40 µM
AgNO3 exhibited 100% survival. Silver nitrate also
induced rooting and flowering in vitro on the rare, rhoeophytic
woody medicinal plant, Rotula aquatica Lour. Dipping of the
basal end of shoots in NAA (2.69 µM) and silver nitrate (11.7 µM)
solution improved rooting efficiency (Sunandakumari
et al. 2004).
Modification
of sex expression
The inhibition
of ethylene action by silver nitrate was employed to suppress the development
of female flowers and induce male flowers (Beyer, 1976c; Atsmon and Tabbak,
1979; Takahashi and Jaffe, 1984). Mulberry (Morus alba L.) is a dioecious
plant and the male and female flowers are seen in separate plants. Bisexual
flowers never occur under natural conditions (Thomas, 2004). By treating the
nodal cuttings with silver nitrate, bisexual flowers could be induced in female
plants. The histological analysis of these bisexual flowers showed both ovule
and anther in the same flower (Thomas, 2004). Bisexual flowers were also
induced in cucumber by silver nitrate treatment (Stankovic and Prodanovic,
2002). Ethylene and auxin promote the formation of female flowers whereas
gibberellins promote the formation of male flowers (Mohan Ram and Jaiswal,
1970; Saito and Takahashi, 1986). The enhancement of femaleness by auxin
possibly occurs through the induction of ethylene biosynthesis (Takahashi and
Jaffe, 1984; Trebitsh et al. 1987). Ethylene evolution is highly correlated
with sex expression in plants and dioecious plants produce more ethylene than
monoecious ones (Rudich et al. 1972; Trebitsh et al. 1987). In view of all
these evidences, silver nitrate may possibly be a potent candidate compound to
regulate the sex expression in plants.
Fruit
ripening
Ethylene plays
a crucial role in initiating and accelerating the ripening-related process.
Treatment of tomato with silver ions has been shown to inhibit ethylene action
and fruit ripening (Hobson et al. 1984). Furthermore, if silver ions were
applied at stages of ripeness well after the breaker stage, ripening can be
arrested (Tucker and Brady, 1987). The growth
regulator 1-methylcyclopropane (1-MCP), like silver ions, is an extremely
effective antagonist for plants or harvested plant products (Serek et al.
1995a; Serek et al. 1995b; Serek et al. 1995c; Sisler et al. 1996).
Leaf
abscission
Ethylene
that stimulated leaf abscission in cotton is blocked by the silver
ion (Beyer,
1976c). Without AgNO3, all the leaves had abscised
on the 7th day in ethylene. Plants treated with increasing
concentrations of AgNO3 and placed in ethylene showed
progressively less leaf abscission. Treatment with 25 mg/l of AgNO3 reduced
the time required to reach 100% leaf abscission by 2 days. Silver
nitrate treatment also reduced the growth retarding effects of ethylene.
Other similar experiments with mature cotton plants have demonstrated
a similar ability of AgNO3 to prevent young fruit and
flower abscission (Beyer, 1976c).
Concluding Remarks
In this review, an
attempt has been made to discuss the role of ethylene, polyamines, and silver
ions as potent regulators of morphogenesis in plants. The interplay of
polyamines, ethylene, and calcium signaling is also discussed. The influence of
exogenously applied silver ions in the form of AgNO3 in plant tissue
culture media significantly regulates the ethylene activity in most of the
plant systems. We have clearly brought out the major physiological effects of
AgNO3 in plant systems viz direct or indirect organogenesis,
somatic embryogenesis, in vitro rooting of micro shoots, induction of
flowering, early flowering, sex expression, and control of leaf abscission.
However, there is a gap in information on the molecular mechanisms of
interaction between silver ions and the ethylene receptors. Further research on
the regulation of morphogenesis through the use of metal ions like silver would
throw light on an array of functions of these relatively simple molecules that
play a marvelous role in influencing growth, development, and adaptation of
plants to the environment. This opens new dimensions
in understanding plant morphogenesis. Hence, it is necessary to elucidate the
physiological mechanisms at the gene regulation level to find out the actual
role of silver ions in signaling and to see how they influence
regulation of ethylene action in plants.
Acknowledgments
VK is grateful to the CSIR, New Delhi for the award
of Research Fellowship. Authors thank Mr. Rithesh Narayanpur for his technical
assistance in preparation of the manuscript.
References
- ABELES, F.B. Ethylene
in plant biology. Academic press New York, 1973. 302 p. ISBN 978-0120414505.
- AL-KHAYRI, J.M. and AL-BAHRANY, A.M.
Silver nitrate
and 2-isopentyladenine promote somatic embryogenesis in date palm (Phoenix
dactylifera L.). Scientia Horticulturae, July 2001, vol. 89, no. 4, p. 291-298. [CrossRef]
- AL-KHAYRI, J.M. and Al-BAHRANY, A.M.
Genotype-dependent in vitro response of date palm (Phoenix dactylifera L.) cultivars to silver nitrate. Scientia Horticulturae, February 2004,
vol. 99, no. 2, p. 153-162. [CrossRef]
- ANTOGNONI, F.; CASALI,
P.; PISTOCCHI, R. and BAGNI, N. Kinetics and calcium-specificity of polyamine
uptake in carrot protoplasts. Amino acids, December 1994, vol. 6, no. 3,
p. 301-309. [CrossRef]
- APELBAUM,
A.; GOLDLUST, A. and ICEKSON, I. Control by ethylene of arginine
decarboxylase activity in pea seedlings and its implication for
hormonal regulation of plant growth. Plant
Physiology, 1985, vol. 79, no. 3, p. 635-640.
- ATSMON, D. and TABBAK, C.
Comparitive effects of gibberellin, silver nitrate and aminoethoxyvinyl glycine
on sexual tendency and ethylene evolution in the cucumber plant (Cucumis
sativus L). Plant Cell Physiology, 1979, vol. 20, no. 8, p. 1547-1555.
- BAIS, H.P.; GEORGE, J. and
RAVISHANKAR, G.A. Influence of polyamines on growth of hairy root cultures of
witl of chiocory (Chichorium intybus L cv Lucknow local) and formation
of coumarins. Journal of Plant Growth Regulation, 1999, vol. 18, no. 1, p.
33-37. [CrossRef]
- BAIS, H.P.; SUDHA, G.; SURESH, B.
and RAVISHANKAR, G.A. AgNO3 influences in vitro root
formation in Decalepis hamiltonii Wight, Arn. Current Science, 2000a, vol. 79, p. 894-898.
- BAIS,
H.P.; SUDHA, G.S. and RAVISHANKAR, G.A. Putrescine and AgNO3 influences
shoot multiplication In vitro flowering and endogenous titres
of polyamines in Chichorium intybus L cv Lucknow Local. Journal
Plant Growth Regulation, 2000b, vol. 19, no. 2, p. 238-248.
- BAIS, H.P.; SUDHA, G.S. and
RAVISHANKAR, G.A. Influence of putrescine AgNO3 and polyamine
inhibitors on the morphogenetic response in untransformed and transformed
tissues of Chichorium intybus and their regenerants. Plant Cell
Reports, 2001a, vol. 20, no. 6, p. 547-555. [CrossRef]
- BAIS, H.P.; SUDHA G.S. and
RAVISHANKAR, G.A. Putrescine influences growth and production of coumarins in
transformed and untransformed root cultures of witloof chicory (Chichorium
intybus L cv Lucknow Local). Acta Physiologia Plantarum, 2001b, vol.
23, p. 319-327.
- BAIS, H.P.; VENKATESH, R.T.; CHANDRASHEKAR,
A. and RAVISHANKAR, G.A. Agrobacterium rhizogenes mediated
transformation of witl of chiocory in vitro shoot regeneration and
induction of flowering. Current Science, 2001c, vol. 80, p. 83-87.
- BAIS, H.P. and RAVISHANKAR G.A. Role
of polyamines in the ontogeny of plants and their biotechnological
applications. Plant Cell Tissue and Organ Culture, 2002, vol. 69, no. 1,
p. 1-34. [CrossRef]
- BEYER, E.M. 14C-ethylene
incorporation and metabolism in pea seedlings. Nature, 1975, vol. 255,
no. 5504, p. 144-147. [CrossRef]
- BEYER, E.M. A potent inhibitor of
ethylene action in plants. Plant Physiology, 1976a, vol. 58, no. 3, p. 268-271.
- BEYER, E.M. Ethylene antidote. HortScience, 1976b, vol. 11, no. 3, p.
174-175.
- BEYER, E.M. Silver ion: a potent anti-ethylene agent in cucumber and
tomato. HortScience, 1976c, vol. 11, no. 3, p. 175-196.
- BEYER, E.M. Effect of silver ion
carbon dioxide and oxygen on ethylene action and metabolism. Plant
Physiology, 1979, vol. 63, p. 163-173.
- BIDDINGTON, N.L. The influence of
ethylene in plant tissue culture. Plant Growth Regulation, 1992, vol. 11,
no. 2, p. 173-178. [CrossRef]
- BLEECKER, A.B.; ESTELLE, M.A.;
SOMERVILLE, C. and KENDE, H. Insensitivity to ethylene conferred by a dominant
mutation in Arabidopsis thaliana. Science, 1988, vol. 241,
no. 4869, p. 1086-1089. [CrossRef]
- BLEECKER, A.B. and KENDE, H. Ethylene:
A gaseous signal molecule in plants. Annual Review of Cell and Developmental
Biology, 2000, vol. 16, p. 1-18. [CrossRef]
- BRAR, Mohanjeet S.; MOORE, Misty J.; AL
KHAYRI, Jameel M.; MORELOCK, Teddy E. and ANDERSON, Edwin J. Ethylene inhibitors promote in vitro regeneration
of cowpea (Vigna ungiculata L.). In Vitro Cellular and
Developmental Biology-Plant, May 1999, vol. 35, no. 3, p. 222-225. [CrossRef]
- BUSH, D.S. Calcium
regulation in plant cells and its role in signaling. Annual Review of Plant
Physiology and Plant Molecular Biology, 1995, vol. 46, p. 95-122. [CrossRef]
- CASTILLO, A.M.; EGAÑA, B.;
SANZ, J.M. and CISTUE, L. Somatic embryogenesis and plant regeneration from
barley cultivars grown in Spain. Plant Cell Reports, 1998, vol. 17, no.
11, p. 902-906. [CrossRef]
- CHANG, C.; KWOK, S.F.; BLEECKER,
A.B. and MEYEROWITZ, E.M. Arabidopsis ethylene-response gene ETR1:
Similarity of product to two-component regulators. Science, 1993, vol. 262,
no. 5133, p. 539-544. [CrossRef]
- CHI,
G.L. and PUA, E.C. Ethylene inhibitors enhanced de novo shoot
regeneration from cotyledons of Brassica
campastris spp in vitro. Plant Science, 1989, vol.
64, p. 243-250. [CrossRef]
- CHI, G.L.; BARFIELD, D.G.; SIM, G.E.
and PUA, E.C. Effect of AgNO3 and amino-ethoxyvinylglycine on in
vitro shoot organogenesis from seedling explants of recalcitrant Brassica genotypes. Plant Cell Reports, 1990, vol. 9, no. 4, p. 195-198. [CrossRef]
- CHI
G.L.; PUA, E.C. and GOH C.J. Role of ethylene on de novo shoot
regeneration from cotyledons of Brassica
campastris spp Pekinensis(Lour) Olsson in vitro. Plant
Physiology, May 1991, vol. 96, no. 1, p. 178-183.
- CHRAIBI, B.K.M.; LATCHE, A.;
RAUSTAN, J.P. and FALLOT, J. Stimulation of shoot regeneration from cotyledons
of Helianthus annuus by ethylene inhibitors silver and cobalt. Plant
Cell Reports, 1991, vol. 10, no. 4, p. 204-207. [CrossRef]
- COATES, G.E.; GREEN, M.L.H. and WADE,
K. Organo metallic compounds II. Methuen and Co Ltd London, 1968, p. 1-376.
- CRUZ
DE CARVALHO, M.H.; VAN LE, B.; ZUILY-FODIL, Y.; PHAM THI, A.T.
and TRAN THANH VAN, K. Efficient whole plant regeneration of common
bean (Phaseolus vulgaris L.) using
thin-cell-layer culture and silver nitrate. Plant Science, 2000,
vol. 159, no. 2, p. 223-232. [CrossRef]
- DAVIES, P.J. The plant
hormones: their nature, occurrence, and functions. In: DAVIES P.J. ed. Plant
Hormones and Their role in Plant Growth and Development, Ch. Al. Boston:
Martinus Nijhoff, 1987.
- DIVYA,
K.; ANURADHA, T.S.; JAMI, S.K. and KIRTI, P.B. Efficient regeneration
from hypocotyl explants in three cotton cultivars. Biologia
Plantarum, 2008, vol. 52, no. 2, p. 201-208. [CrossRef]
- DUNCAN,
D.R.; WILLIAMS, M.E.; ZEHR B. and WIDHOLM, J.M. The production
of callus capable of plant regeneration from immature embryos of
numerous Zea mays genotypes. Planta, 1985, vol. 165, no. 3,
p. 322-332. [CrossRef]
- EAPEN,
S. and GEORGE L. Plant regeneration from peduncle segments of oil
seed brassica species: Influence of AgNO3 and silver
thiosulphate. Plant Cell Tissue and Organ
Culture, 1997, vol. 51, no. 3, p. 229-232. [CrossRef]
- EL MESKAOUI, A.; DESJARDINS, Y. and TREMBLAY,
F.M. Kinetics of ethylene biosynthesis and its effects during maturation of
white spruce somatic embryos. Physiologia Plantarum, 2000, vol. 109, p.
333-342. [CrossRef]
- EL-ITRIBY, H.A.; ASSEM, S.K.; HUSSEIN,
E.H.A.; ABDEL-CALIL, F.M. and MADKOUR, M.A. Regeneration and transformation of
egyptian maize inbred lines via immature embryo culture and a biolistic
particle delivery system. In Vitro Cellular and Development Biology-Plant,
2003, vol. 39, no. 5, p. 524-531. [CrossRef]
- EVANS, P.T. and MALMBERG, R.L. Do
polyamines have role in plant development? Annual Review of Plant Physiology
and Plant Molecular Biology, 1989, vol. 40, p. 235-269. [CrossRef]
- EVANS, J.M. and BATTY, N.P. Ethylene
precursors and antagonists increase embryogenesis of Hordeum vulgare L anther culture. Plant Cell Reports, 1994, vol. 13, no. 11, p. 676-678. [CrossRef]
- FEI, S.; READ, P.E. and
RIORDAN, T.P. Improvement of embryogenic callus induction and shoot
regeneration of buffalo grass by AgNO3. Plant Cell Tissue Organ
Culture, 2000, vol. 60, no. 3, p. 197-203. [CrossRef]
- FEIRER, R.P.; MIGNON, G. and
LITVAY, J.D. Arginine decarboxylase and polyamines required for embryogenesis
in wild carrot. Science, 1984, vol. 223, no. 4643, p. 1433-1435. [CrossRef]
- FERNANDEZ, S.;
MICHAUX-FERRIÈRE, N. and COUMANS, M. The embryogenic response of immature
embryo cultures of durum wheat (Triticum durum): histology and
improvement by AgNO3. Plant Growth Regulation, 1999, vol. 28,
no. 3, p. 147-155. [CrossRef]
- FLINK, L. and PETTIJOHN,
D.E. Polyamines stabilize DNA folds. Nature, 1975, vol. 253, no. 5486,
p. 62-63. [CrossRef]
- FUENTES, S.R.L.; CALHEIROS, M.B.P.; MANETTI-FILHO, J. and
VIEIRA, L.G.E. The effects of silver nitrate and different carbohydrate sources
on somatic embryogenesis in Coffea canephora. Plant Cell Tissue Organ
Culture, 2000, vol. 60, no. 1, p. 5-13. [CrossRef]
- GALSTON, A.W.
Polyamines as modulators of plant development. Bioscience, 1983, vol.
33, p. 382-388.
- GANESH, S.D.
and SREENATH, H.L. Silver nitrate enhanced shoot development in cultured apical
shoot buds of Coffea arabica Cv Cauvery (S4347). Journal of
Plantation Crops, 1996, vol. 24, p. 577-580.
- GAVINLERTVATANA, P.; READ, P.E. and
RIORDAN, T.P. Control of ethylene synthesis and action by silver nitrate and
rhizobitoxine in petunia leaf sections cultured in vitro. Journal of the
American Society for Horticultural Science, 1980, vol. 105, p. 304-307.
- GIRIDHAR, P.; OBUL REDDY, B. and
RAVISHANKAR, G.A. Silver nitrate influences in vitro shoot
multiplication and root formation in Vanilla planifolia Andr. Current
Science, November 2001, vol. 81, no. 9, p. 1166-1170.
- GIRIDHAR, P.; INDU, E.P.; VIJAYA RAMU, D. and
RAVISHANKAR, G.A. Effect of silver nitrate on in vitro shoot growth of Coffee. Tropical Science, 2003, vol. 43, no. 3, p. 144-146.[CrossRef]
- GIRIDHAR, P.; INDU, E.P.; VINOD, K.;
CHANDRASHEKAR, A. and RAVISHANKAR, G.A. Direct somatic embryogenesis from Coffea
arabica L and Coffea canephora P ex Fr. under the influence of
ethylene action inhibitor-silver nitrate. Acta Physiologiae Plantarum, September 2004, vol. 26, no. 3, p.
299-305. [CrossRef]
- GONG, Y.; GAO, F. and TANG, K. In vitro high frequency direct root and shoot regeneration in sweet potato using the
ethylene inhibitor silver nitrate. South African Journal of Botany, 2005, vol. 71, p. 110-113.
- HALL, A.E.; CHEN, Q.G.; FINDELL,
J.L.; SCHALLER, G.E. and BLEEKER, A.B. The relationship between ethylene
binding and dominant insensitivity conferred by mutant forms of the ETR1
ethylene receptor. Plant Physiology, 1999, vol. 121, no. 1, p. 291-299.
- HAMILTON, A.J.; BOUZAYEN, M. and
GRIERSON, D. Identification of a tomato gene for the ethylene-forming enzyme by
expression in yeast. Proceedings of the National Academy of Sciences of
the United States of America, 1991, vol. 88, no. 16, p. 7434-7437.
- HOBSON, G.E.;
NICHOLS, R.; DAVIES, J.N. and ATKEY, P.T. The inhibition of tomato fruit
ripening by silver. Journal of Plant Physiology, 1984, vol. 116, no. 1,
p. 21-29.
- HUA, J.; CHANG, C.; SUN, Q. and
MEYEROWITZ, E.M. Ethylene insensitivity conferred by Arabidopsis ERS
gene. Science 1995, vol. 269, p. 1712–1714.
- HUA, J. and MEYEROWITZ, E.M.
Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis
thaliana. Cell, 1998, vol. 94, no. 2, p. 261-271. [CrossRef]
- HYDE, Camille L. and PHILLIPS,
Gregory C. Silver nitrate
promotes shoot development and plant regeneration of chili pepper (Capsicum
annuum L.) via organogenesis. In Vitro Cellular and Developmental
Biology-Plant, April 1996, vol. 32, no. 2, p. 72-80. [CrossRef]
- JHA,
A.K.; DAHLEEN, L.S. and SUTTLE, J.C. Ethylene influences green
plant regeneration from barley callus. Plant Cell Reports, 2007, vol. 26, no. 3, p. 285-290. [CrossRef]
- JOHNSON, P.R. and ECKER, J.R. The
ethylene gas signal transduction pathway: A molecular perspective. Annual
Review of Genetics, 1998, vol. 32, p. 227-254. [CrossRef]
- KENDE, H. Ethylene biosynthesis. Annual
Review of Plant Physiology and Plant Molecular Biology, 1993, vol. 44, p.
283-307. [CrossRef]
- KHALAFALLA, M.M. and HATTORI, K. Ethylene
inhibitors enhance in vitro root formation on faba bean shoots
regenerated on medium containing thidiazuron. Plant Growth Regulation,
2000, vol. 32, no. 1, p. 59-63. [CrossRef]
- KIEBER, J.J.; ROTHENBERG, M.; ROMAN,
G.; FELDMANN, K.A. and ECKER, J.R. CTR1, a negative regulator of the ethylene response
pathway in Arabidopsis, encodes a member of the raf family of protein
kinases. Cell, 1993, vol. 72, no. 3, p. 427-441. [CrossRef]
- KONG, L and YEUNG, E.C. Effects of ethylene and ethylene
inhibitors on white spruce somatic embryo maturation. Plant Science, 1994, vol. 104, no. 1, p. 71-80. [CrossRef]
- KOTHARI-CHAJER,
A.; SHARMA, M.; KACHHWAHA, S. and KOTHARI S.L. Micronutrient optimization
results into highly improved in vitro plant regeneration
in kodo (Paspalum scrobiculatum L.) and finger (Eleusine
coracana (L.) Gaertn.) millets. Plant Cell
Tissue and Organ Culture, 2008, vol. 94, no. 6, p. 105-112. [CrossRef]
- KUMAR,
P.P.; LAKSHMANAN, P. and THORPE, T.A. Regulation of morphogenesis
in plant tissue culture by ethylene. In
Vitro Cellular and Developmental Biology-Plant, 1998a, vol.
34, no. 2, p. 94-103. [CrossRef]
- KUMAR, S.; SARKAR, A.K. and KUNHIKANNAN. Regeneration
of plants from leaflet explants of tissue culture raised Safed siris (Albizia
procera). Plant Cell Tissue Organ Culture, 1998b, vol. 54, no. 3, p.
137-143. [CrossRef]
- KUMAR, V.; GURURAJ, H.B.; PRASAD,
B.C.N.; SHARMA, A.; GIRIDHAR, P. RAJASEKARAN, T. and RAVISHANKAR, G.A. In vitro morphogenetic
responses and regeneration from pepper (Capsicum annuum) seedling
explants. In: Proceedings of 5th International Food Convention.
CFTRI, Mysore, India, 2003a, p. 41.
- KUMAR,
V.; SATHYANARAYANA, K.V.; INDU, E.P.; SARALA ITTY, S.; GIRIDHAR,
P.; CHANDRASHEKAR, A. and RAVISHANKAR, G.A. Stable transformation
and direct regeneration in Coffea
canephora by Agrobacterium rhizogenes mediated transformation.
In: Proceedings
of 10th Congress of Federation of Asian and Oceanian
Biochemists and Molecular Biologists. 2003b, p. 10.
- KUMAR, S.V. and RAJAM,
M.V. Polyamine ethylene nexus: A potential target for post harvest
biotechnology. Indian Journal of Biotechnology, 2004, vol. 3, no. 2, p.
299-304.
- KUMAR, V.; RAMAKRISHNA, A. and
RAVISHANKAR, G.A. Influence of different ethylene inhibitors on somatic
embryogenesis and secondary embryogenesis from Coffea canephora P ex Fr. Plant Cell Tissue and Organ Culture, 2007, vol. 43, no. 6, p. 602-607. [CrossRef]
- LAU, Oi-Lim and YANG, Shang F. Inhibition
of ethylene production by cobaltous ion. Plant Physiology, July 1976, vol. 58, no. 1, p. 114-117.
- LENTINI,
Z.; MUSSEL, H.; MUTSCHLER, M.A.; and EARLE, E.D. Ethylene generation
and reversal of ethylene effects during development of in vitro Brassica
compestris life. Plant
Science, 1988, vol. 54, no. 1, p. 75-81. [CrossRef]
- LENTINI, Z.; REYES, P.;
MARINEZ, C.P. and ROCA, W.M. Androgenesis of highly recalcitrant rice genotypes
with maltose and silver nitrate. Plant Science, 1995, vol. 110, no. 1,
p. 127-138. [CrossRef]
- LIEBERMAN, N.
Biosynthesis and action of ethylene. Annual Review of Plant Physiology,
1979, vol. 30, p. 533-591. [CrossRef]
- LIM,
H.T and SONG, Y.N. Effect of explant type and silver nitrate on
callus induction and organogenesis organogenesis of watermelon. Journal
of Agricultural Sciences, 1993,
vol. 5, p. 113-121.
- MAJEWSKA-SAWKA, A.;
NIKLAS, A. and JAZDZEWSKA. The effect of polyamines on the development of sugar
beet protoplasts. Biologia Plantarum, 1997, vol. 39, no. 4, p. 561-567. [CrossRef]
- MAJEWSKA-SAWKA, A.;
BUTOWT, R. and NIKLAS, A. Do polyamines release membrane-bound calcium in sugar
beet protoplasts? Journal of Plant Physiology, 1998, vol. 153, no. 1-2,
p. 247-250.
- MARTIN, K.P. Plant regeneration protocol of
medicinally important Andrographis paniculata (Burm. F.) Wallich ex Nees via somatic embryogenesis. In Vitro Cellular and Development Biology-Plant,
2004, vol. 40, no. 2, p. 204-209. [CrossRef]
- MARTIN,
K.P. and MADASSERY, J. Direct and indirect somatic embryogenesis
on cotyledon explants of Quassia amara L., an antileukaemic
drug plant. In Vitro Cellular and Development Biology-Plant,
2005, vol. 41, no. 1, p. 54-57. [CrossRef]
- MATTOO, A.K. and SUTTLE,
J.C. The plant hormone ethylene. CRC
press. Boca Raton, Florida, 1991. 337 p. ISBN 0-8493-4566-9.
- MISRA, P. and DATTA, S.K. Direct
differentiation of shoot buds in leaf segments of white merigold (Tagetes
erecta L). In Vitro Cellular Developmental Biology-Plant, 2001, vol. 37, no. 4, p. 466-470. [CrossRef]
- MIYAZAKI,
J.H. and YANG, S.F. The methionine salvage pathway in relation
to ethylene and polyamine biosynthesis. Physiologia Plantarum,
1987, vol. 69, no. 2, p. 366-370. [CrossRef]
- MOHAN
RAM, H.Y. and JAISWAL, V.S. Induction of female flowers on male
plants of Cannabis sativa L. by
2-chloroetahanephosphonic acid. Experientia, 1970, vol. 26,
p. 214-216.
- MOHIUDDIN, A.K.M.;
CHOWDHURY, M.K.U.; ABDULLA, Z.C. and NAPIS, S. Influence of silver nitrate
(ethylene inhibitor) on Cucumber shoot regeneration. Plant Cell, Tissue and
Organ Culture, 1997, vol. 51, no. 1, p. 75-78. [CrossRef]
- NAIK,
S.K. and CHAND, P.K. Silver nitrate and aminoethoxy venyl glycene
promotes in vitro adventitious
shoot regeneration of Pomegranate (Punica granatum L). Journal
of Plant Physiology, 2003, vol. 160, no. 4, p. 423-430. [CrossRef]
- NARASIMHULU,
S.B. and CHOPRA, V.L. Species specific shoot regeneration response
of cotyledonary explants of brassicas. Plant Cell Reports, 1988,
vol. 7, no. 2, p. 104-106. [CrossRef]
- NISSEN, P. Stimulation of
somatic embryogenesis in carrot by ethylene: Effects of modulators of ethylene
biosynthesis and action. Physiologia Plantarum, 1994, vol. 92, no. 3, p.
397-403. [CrossRef]
- OGURA, H. and SHIMADA,
T. Preliminary report on shoot differentiation from Wheat callus. Wheat
information Service, 1978, no. 45, p. 26-28.
- O'KENNEDY, M.M.; SMITH, G. and BOTHA, F.C.
Improved regeneration efficiency of a pearl millet (Pennisetum glaucum [L.]
R. Br.) breeding line. South African Journal of Botany, 2004, vol. 70,
no. 4, p. 502-508.
- OLDACH, K.; MORGENSTERN, A.; ROTHER,
S.; GIRGI, M.; O`KENNEDY, M. and LORZ, H. Efficient in vitro plant
regeneration from immature zygotic embryos of Pearl millet [Pennisettum
glaucum (L) R Br] and Sorgum bicolor (L) Moench. Plant Cell
Reports, 2001, vol. 20, no. 5, p. 416-421. [CrossRef]
- OZUDOGRU, E.; OZDEN TOKATLI, Y. and AKCIN, A. Effect of silver nitrate on
multiple shoot formation of Virginia-type peanut through shoot tip culture. In
Vitro Cellular and Development Biology-Plant, 2005, vol. 41, no. 2, p. 151-156. [CrossRef]
- PALMER, C.E. Enhanced shoot
regeneration from Brassica campestris by silver nitrate. Plant Cell Reports, 1992, vol. 11, no. 11, p.
541-545. [CrossRef]
- PATIL, P.; CHANDRA, R.;
SANGEETA, K. and RAGHUVEER, P. Influence of polyamines and ethylene inhibitors
on somatic embryo induction in chickpea (Cicer arietinum L.). Indian
Journal of Plant Physiology, 1999, vol. 3, p. 26-31.
- PESTANA,
M.C.; LACORTE, C.; DE FREITAS, V.G.; OLIVEIRA, D.E. and MANSUR,
E. In vitro regeneration of
peanut (Arachis hypogaea L) through organogenesis: Effect
of culture temperature and silver nitrate. In vitro Cellular and
Developmental Biology-Plant, 1999, vol. 35, no. 3, p. 214-216. [CrossRef]
- PLUS, J.L.; GEORGE, L.;
EAPEN, S. and RAO, P.S. Enhanced plant regeneration in pearl millet
(Pennisettum americanum). Plant Cell, Tissue and Organ Culture, 1993,
vol. 32, no. 1, p. 91-96. [CrossRef]
- PREM, D.; GUPTA, K. and AGNIHOTRI, A. Effect
of various exogenous and endogenous factors on microspore embryogenesis in
indian mustard (Brassica juncea (L.) Czern and Coss). In Vitro
Cellular and Developmental Biology-Plant, 2005, vol. 41, no. 3, p.
266-273. [CrossRef]
- PUA, E.C. and CHI, G.L. De novo shoot morphogenesis and plant growth of mustard (Brassica juncea) in
vitro in relation to ethylene. Physiologia Plantarum, 1993, vol. 88,
no. 3, p. 467-474. [CrossRef]
- PUA, E-C.; SIM, G-E.; CHI, G-L. and KONG,
L-F. Synergistic effects of ethylene inhibitors and putrescine on shoot
regeneration from hypocotyl explants of Chinese radish (Raphanus sativus L. var. longipinnatus Bailey) in vitro. Plant Cell Reports, May 1996, vol. 15, no. 9, p. 685-690. [CrossRef]
- PUA, E.C.; DENG, X. and KOH, A.T.C.
Genotypic variability of de novo shoot morphogenesis Brassica oleracea in vitro in response to ethylene inhibitors and Putrescine. Journal
of Plant Physiology, 1999, vol. 155, no. 4-5, p. 598-605.
- PURNHAUSER,
L.; MEDGYSEY, P.; CZAKO, M.; DIX, J.P. and MARTON, L. Stimulation
of shoot regeneration in Triticum
aestivum and Nicotiana plumbaginifolia Viv tissue cultures
using the ethylene inhibitor silver nitrate. Plant Cell Reports,
1987, vol. 6, no. 1, p. 1-4. [CrossRef]
- RAVANEL, S.; GAKIERE, B.; JOB, D.
and DOUCE, R. The specific features of methionine biosynthesis and metabolism
in plants. Proceedings of the National Academy of Sciences of the
United States of America, 1998, vol. 95, p. 7805-7812.
- REDDY, B.O.; GIRIDHAR, P. and
RAVISHANKAR, G.A. In vitro rooting of Decalepis hamiltonii Wight
and Arn., an endangered shrub by auxins and root-promoting agents. Current
Science, 2001, vol. 81, no. 11, p. 1479-1481.
- REIS,
L.B.; PAIVA NETO, V.B.; TOLEDO PICOLI, E.A.; COSTA, M.G.C.; REGO,
M.M.; CARVALHO, C.R.; FINGER, F.L. and
OTONI, W.C. Axially bud development of passion fruit as affected
by ethylene precursor and inhibitors. In Vitro Cellular Developmental
Biology-Plant,
2003, vol. 39, no. 6, p. 618-622. [CrossRef]
- RODRIGUEZ, F.I.; ESCH, J.J.; HALL,
A.E.; BINDER, B.M.; SCHALLER, G.E. and BLEECKER, A.B. A copper cofactor for the
ethylene receptor ETR1 from Arabidopsis. Science, 1999, vol. 283,
no. 5404, p. 996-998. [CrossRef]
- ROUSTAN,
J.P.; LATCHE, A. and FALLOT, J. Stimulation of Daucus carota somatic
embryogenesis by inhibitors of ethylene biosynthesis: cobalt and
nickel. Plant Cell Reports, March 1989, vol. 8, no. 3, p.
182-185. [CrossRef]
- ROUSTAN,
J.P.; LATCHE, A. and FALLOT, J. Control of carrot somatic embryogenesis
by AgNO3 an inhibitor of ethylene action effect on arginine
decarboxilase activity. Plant Science, 1990, vol. 67, no.
1, p. 89-95. [CrossRef]
- RUDICH, J.; HALEVY, A.H. and KEDAR,
N. Ethylene evolution from cucumber plants as related to sex expression. Plant
Physiology, 1972, vol. 49, no. 6, p. 998-999.
- SAITO,
T. and TAKAHASHI, H. Role of leaves in ethylene induced femaleness
in cucumber plants. Journal of the Japanese
Society for Horticultural Science, 1986, vol. 55, no. 4, p.
445-454.
- SAKAI, H.;
HUA, J.; CHEN, Q.G.; CHANG, C.; MEDRANO, L.J.; BLEECKER, A.B. and MEYEROWITZ,
E.M. ETR2 is an ETR1-like gene involved in ethylene signaling in Arabidopsis. Proceedings of the National Academy of Sciences of the United States
of America, 1998, vol. 95, p. 5812-5817.
- SANKHLA, D.; SANKHLA, N. and DAVIS,
D.T. Promotion of in vitro shoot formation from excised roots of
silktree (Albizzia julibrissim) by an oxime ether derivative and other
ethylene inhibitors. Plant Cell Reports, January 1995, vol. 15, no. 1-2,
p. 143-146. [CrossRef]
- SATO, T. and THEOLOGIS, A. Cloning
the mRNA encoding 1-aminocyclopropane-1-carboxylate synthase, the key enzyme
for ethylene biosynthesis in plants. Proceedings of the National Academy of
Sciences of the United States of America, 1989, vol. 86, no. 17, p.
6621-6625.
- SCHALLER, G.E. and BLEECKER, A.B. Ethylene-binding sites
generated in yeast expressing the Arabidopsis ETR1 gene. Science,
1995, vol. 270, no. 5243, p. 1809-1811. [CrossRef]
- SEONG, E.S.; SONG, K.; JEGAL, S.; YU, C. and
CHUNG, I. Silver nitrate and aminoethoxy vinylglycine affect Agrobacterium-mediated
apple. Plant Growth Regulation, 2005, vol. 45, no. 1, p. 75-82. [CrossRef]
- SEREK,
M.; SISLER, E.C. and REID, M.S. 1-methylcyclopropene a novel gaseous
inhibitor of ethylene action improves the life of fruits cut flowers
and potted plants. Acta Horticulturae, 1995a,
vol. 394, p. 337-347.
- SEREK,
M.; SISLER, E.C. and REID, M.S. Effect of 1-MCP on the vase life
and ethylene response of cut flowers. Plant Growth Regulation,
1995b, vol. 16, no. 1, p. 93-97. [CrossRef]
- SEREK, M.; SISLER, E.C.; TIROSH, T.
and MAYAK, S. 1-methylcyclopropene prevents bud flower and leaf abscission of
Geraldton Wax flower. Hortscience, 1995c, vol. 30, no. 6, p. 1310.
- SISLER, E.C.; SEREK, M. and DUPILLE,
E. Comparison of cyclopropene 1 methylcyclopropene and 1-3 dimethyl
cyclopropene as ethylene antagonists in plants. Plant Growth Regulation, 1996, vol. 18, no. 3, p. 169-174. [CrossRef]
- SMITH,
T.A. Plant polyamines metabolism and function Polyamine synthesis.
In: FLORES, H.E.; ARTECA, R.N. and SHANNON, J.C. eds. Polyamines
and ethylene: biochemistry, physiology and interaction. American
Society of Plant Physiologists, Rockville, Md,
1985. p. 1-23.
- SMITH, T.A. Amines. In:
HARBORNE, J.B. and DEY, P.M. eds. Methods in Plant Biochemistry (Alkaloids
and Sulphur compounds). Academic Press, 1993, p. 17-49.
- SONGSTAD,
D.D.; DUNCAN, D.R. and WIDHOLM, J.M. Effect of 1-aminocycopropane-1-carboxilic
acid silver nitrate and norbornadiene on plant regeneration from
maize callus cultures. Plant Cell
Reports, 1988, vol. 7, no. 4, p. 262-265. [CrossRef]
- SONGSTAD, D.D.; ARMSTRONG, C.L. and
PETERSEN, W.L. Silver nitrate increase type II callus production from immature
embryos of maize inbred B73 and its derivatives. Plant Cell Reports,
1991, vol. 9, no. 12, p. 699-702. [CrossRef]
- SPANU, P.; REINHARDT, D. and BOLLER,
T. Analysis and cloning of the ethylene-forming enzyme from tomato by
functional expression of its mRNA in Xenopus laevis oocytes. EMBO
Journal, 1991, vol. 10, no. 8, p. 2007-2013.
- STANKOVIC,
L. and PRODANOVIC, S. Silver nitrate affects on sex expression
in cucumber (Cucumis sativus).
II Balkan symposium on Vegetables and Potatoes. Acta Hortoculturae, 2002,
vol. 579, p. 203-206.
- STEINMACHER, D.A.;
CANGAHUALA-INOCENTE, G.C.; CLEMENT, C.R. and GUERRA, M.P. Somatic embryogenesis
from peach palm zygotic embryos. Plant Cell Tissue and Organ Culture,
2007, vol. 43, no. 2, p. 124-132. [CrossRef]
- SUNANDAKUMARI, C.;
MARTIN, K.P.; CHITHRA, M. and MADHUSOODANAN, P.V. Silver nitrate induced
rooting and flowering in vitro on rare rhoeophytic woody medicinal
plant, Rotula aquatica Lour. Indian Journal of Biotechnology,
July 2004, vol. 3, no. 3, p. 418-421.
- TABOR,
C.W. and TABOR, H. Polyamines. Annual Review of Biochemistry,
1984, vol. 53, p. 749-790. [CrossRef]
- TAKAHASHI, H. and JAFFE, M.J.
Further studies of auxin and ACC induced feminization in cucumber plant using
ethylene inhibitors. Phyton, July 1984, vol. 44, no. 1, p. 81-86.
- THOMAS, T.D. In vitro modification of sex expression in Mulberry (Morus alba) by ethrel and
silver nitrate. Plant Cell Tissue Organ Culture, 2004, vol. 77, no. 3,
p. 277-281. [CrossRef]
- TREBITSH, T.; RIOV, J. and RUDICH,
J. Auxin, Biosynthesis of ethylene and sex expression in cucumber (Cucumis
sativus). Plant Growth Regulation, 1987, vol. 5, no. 2, p. 105-113. [CrossRef]
- TSAO, C.W.V. and REED, B.M. Gelling agents, silver
nitrate, and sequestrene iron influence adventitious shoot and callus formation
from Rubus leaves. In Vitro Cellular and Developmental Biology-Plant,
2002, vol. 38, no. 1, p. 29-32. [CrossRef]
- TUCKER, G.A. and BRADY, C.J. Silver
ions interrupt tomato fruit ripening. Journal of Plant Physiology, 1987,
vol. 127, no. 1-2, p. 165-169.
- VAIN HORT, Y.P. and FLAMENT,
P. Role of ethylene in embryogenic callus initiation and regeneration in Zea
mays L. Journal of Plant Physiology, 1989, vol. 135, no. 5, p.
537-540.
- VAIN
HORT, Y.P.; YEAN, H. and FLAMENT, P. Enhancement of production
and regeneration of embryogenic type II callus in Zea mays L
by AgNO3. Plant Cell Tissue Organ
Culture, 1989, vol. 18, no. 2, p. 143-142. [CrossRef]
- VALDEZ-ORTIZ, A.; MEDINA-GODOY, S.;
VALVERDE, M.E. and PAREDES-LOPEZ, O. A transgenic tropical maize line generated
by the direct transformation of the embryo-scutellum by A. tumefaciens. Plant
Cell Tissue and Organ Culture, 2007, vol. 91, no. 3, p. 201-214. [CrossRef]
- VAN DEN BROECK, D.; VAN DEN
STRAETEN, D.; VAN MONTAGUE, M. and CAPLAN, A. A group of chromosomal proteins is
specifically released by spermine and loses DNA-binding activity upon
phosphorylation. Plant Physiology, 1994, vol. 106, no. 2, p. 559-566.
- VASIL, I.K. Developing
cell and tissue culture system for the improvement of cereal and grass crops.
Journal Plant Physiology, 1987, vol. 128, no. 3, p.193-218.
- VEEN,
H. and OVER BEEK, J.H.M. The action of silver thiosulphate in carnation
petals. In: CLIGSTERS, H.D.; PROFT, M.; MARCELLE, R. and VAN POUCKE,
M. eds. Biochemical and
physiological aspects of ethylene production in lower and higher
plants. Kluwer Academic Publication. Dordedrecht, The Netherlands,
1989, p. 109-117.
- WALDEN, R.; CORDEIRO, A.
and TIBURIO, A. Polyamines: Small molecules triggering pathways in plant growth
and development. Plant Physiology, 1997, vol. 113, no. 4, p. 1009-1013.
- WANG, K.L.C.; HAI, L. and ECKER,
J.R. Ethylene biosynthesis and signaling networks. Plant Cell, 2002,
S131-S151 Supplement.
- WANG,
G.L. and XU, Y.N. Hypocotyl-based Agrobacterium-mediated
transformation of soybean (Glycine
max) and application for RNA interference. Plant Cell Reports,
2008, vol. 27, no. 7, p. 1177-1184. [CrossRef]
- WOESTE,
Keith E. and KIEBER, Joseph J. A strong loss-of-function mutation
in RAN1 results in constitutive activation of the ethylene response
pathway as well as a rosette-lethal phenotype. Plant Cell, 2000,
vol. 12, p. 443-455.
- WU,
L.M.; WEI, Y.M. and ZHENG, Y.L. Effects of silver nitrate on the
tissue culture of immature wheat embryos. Russian
Journal of Plant Physiology, 2006, vol. 53, no. 4, p. 530-534. [CrossRef]
- WURGLER-MURPHY, S.M. and
SAITO, H. Two-component signal transducers and MAPK cascades. Trends in
Biochemical Sciences, 1997, vol. 22, no. 5, p. 172-176. [CrossRef]
- YANG, S.F. and HOFFMAN, N.E.
Ethylene biosynthesis and its regulation in higher plants. Annual Review of
Plant Physiology, 1984, vol. 35, p. 155-189. [CrossRef]
- YANG, S.F. Biosynthesis and action
of ethylene. HortScience, 1985, vol. 20, no. 1, p. 41-45.
- YU,
Y.; WANG, J.; ZHU, M.L. and WEI, Z.M. Optimization of mature embryo-based
high frequency callus induction and plant regeneration from elite
wheat cultivars grown in China. Plant
Breeding, 2008, vol. 127, no. 3, p. 249-255. [CrossRef]
- ZANG, P.; PHANSIRI, S. and KAERLAS,
J.P. Improvement of cassava shoot organogenesis by the use of AgNO3 in
vitro. Plant Cell Tissue Organ Culture, 2001, vol. 67, no. 1, p. 47-54. [CrossRef]
- ZHAO, X.C.; QU, X.; MATHEWS, D.E.
and SCHALLER, G.E. Effect of ethylene-pathway mutations upon expression of the
ethylene receptor ETR1 from Arabidopsis. Plant Physiology, 2002, vol.
130, no. 4, p. 1983-1991.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication.
Supported by UNESCO / MIRCEN network
© 2009 by Pontificia Universidad Católica de Valparaíso -- Chile
The following images related to this document are available:
Photo images
[ej09015f3.jpg]
[ej09015f2.jpg]
[ej09015f1.jpg]
|

{kind=link}
{kind=link}
{kind=link}