 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 12, No. 3, July 15, 2009 Diversity of marine gliding bacteria in Thailand and their cytotoxicity Yutthapong Sangnoi1 , Pornpoj Srisukchayakul2 , Vullapa Arunpairojana3 , Akkharawit Kanjana-Opas*4
1 Department of Industrial Biotechnology,
Faculty of Agro-Industry,
Prince of Songkla University,
Hat Yai, Songkhla 90112, Thailand Financial support: Thailand Research Fund (MRG4880153) and a Biodiversity Research and Training Grant (BRTR_149011). Scholarship for YS from the Graduate School, Prince of Songkla University. Code Number: ej09023 Abstract Eighty-four marine gliding bacteria were isolated from specimens collected in the Gulf of Thailand and the Andaman Sea. All exhibited gliding motility and swarm colonies on cultivation plates and they were purified by subculturing and micromanipulator techniques. Their 16S rRNA genes were amplified by the polymerase chain reaction (PCR) technique. The phylogenetic analysis indicated that the represented isolates can be separated into six different clads (gr 1 - gr 6) within the Cytophaga-Flavobacterium-Bacteriodes (CFB) group. Group 1 formed a remote linear, with only 90% sequence similarity, from Flavobacteriaceae bacterium which indicated a potentially novel taxonomic group. Groups 2 and 3 were identified as the recently proposed Tenacibaculum mesophilum and Fulvivirga kasyanovii respectively. Groups 4, 5 and 6, consisting of the largest number of the members, were identified as Rapidithrix thailandica, Aureispira marina and Aureispira maritima respectively. The isolates were cultivated in four different cultivation media (Vy/2, RL 1, CY and SK) and the crude extracts were submitted to screen cytotoxicity using a sulphorodamine B (SRB) assay. The results from cytotoxic screening showed that groups 2, 4 and 6 were capable of producing the cytotoxic metabolites against selected human cell lines (breast adenocarcinoma (MCF-7), colon cancer (HT-29), cervical cancer (HeLa) and oral cancer (KB)). However, groups 1, 3 and 5 did not produce metabolites with cytotoxicity when cultivated in the same cultivation media as the previous groups. CY medium was the only cultivation medium which could yield the cytotoxic metabolites against MCF-7. Keywords: Aureispira marina, Aureispira maritime, Fulvivirga kasyanovii, human cell lines, Rapidithrix thailandica, Tenacibaculum mesophilum.
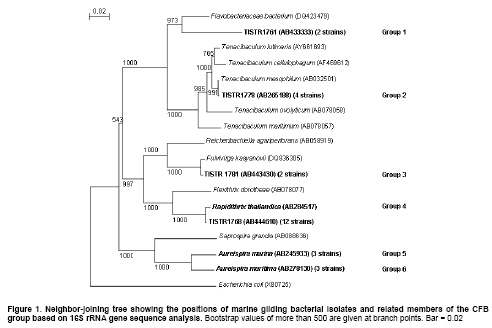
Research to discover anticancer drugs have been the major focus of natural product researchers worldwide. This is due to the increasing number of cancer patients, the lack of selectivity of most anticancer agents (Miot-Noirault et al. 2004), the development of resistance to drugs (Hutson et al. 2004) and the serious adverse side effects of currently available anticancer drugs (Schilder et al. 2004). Therefore, the need for new cancer drugs is very crucial. In the past thirty years, several researchers have demonstrated that marine organisms, especially marine invertebrates are capable of producing cytotoxic compounds (Izbicka et al. 1998; Boyd et al. 2001; Curiel et al. 2001). However, marine microorganisms are understudied and only a few cytotoxic compounds have been reported so far. The examples are palau’amide (Williams et al. 2003), salinosporamide A (Feling et al. 2003), micromide (Williams et al. 2004), resistoflavine (Gorajana et al. 2007), and trichodermamide B (Garo et al. 2003). Interestingly, cytotoxic compounds from marine microorganisms have been produced mainly by marine fungi and actinomycetes. Gliding bacteria are the group of Gram negative bacteria with unique gliding motility and distributed in various habitats (Johansen et al. 1999; Barbeyron et al. 2001; Nakagawa et al. 2001; Hosoya et al. 2006; Hosoya et al. 2007; Srisukchayakul et al. 2007). Despite the increasing number of reports on isolation of novel marine gliding bacteria, little is known about their secondary metabolites and their potential uses as sources of bioactive compounds. So far only few metabolites, haliangicin (Fudou et al. 2001), neoverrucosane diterpenoids (Spyere et al. 2003), miuraenamides (Iizuka et al. 2006) and marinoquinoline A (Kanjana-Opas et al. 2006), have been reported from marine gliding bacteria indicating they are untapped resources awaiting exploitation. Thailand is located in the high biodiversities zone having uniquely different marine habitats both in the Indian Ocean (Andaman Sea) and the Pacific Ocean (Gulf of Thailand). Although the biodiversity of marine gliding bacteria are understudied in Thailand and elsewhere, our reports on discovery of marine gliding bacteria in Thailand in the past decade have shown the promising results (Hosoya et al. 2006; Hosoya et al. 2007; Srisukchayakul et al. 2007). In this paper we aim to report the isolation, identification, and classification of novel groups of marine gliding bacteria from Thailand and their potential as sources of cytotoxic compounds. Materials and Methods In this study, a modified seawater glutamate medium (SWG) containing L-Glutamic acid monosodium salt, 1.0 g; NH4NO3, 0.01 g; K2HPO4, 0.01 g; agar 15 g and seawater, 1 L (Hosoya et al. 2006) was used for the isolation of marine gliding bacteria from marine specimens such as biofilms, invertebrates and plant materials. These were collected in Thai sea water. All specimens, after rinsing with sterile seawater, were cut into small pieces (0.5 x 0.5 cm) with a sterilized sharp blade and placed on the isolation media in Petri dishes and incubated at room temperature. Swarm colony formation was observed under a stereomicroscope and a clean edge of swarm colony was cut into a small agar blocks by a sterilized sharp blade. These were transferred to a new cultivation plate containing modified SAP2 medium (tryptone, 1 g; yeast extract, 1 g; agar, 15 g; sea water, 1 L) (Reichenbach, 1991) and incubated at room temperature. The subculturing was repeated until a pure culture could be obtained. For some isolates, the isolation of a single cell by a micromanipulator was performed in order to obtain a pure culture (Skerman, 1968; Sly and Arunpairojana, 1987). 16S rRNA gene sequences and phylogenetic analyse The genomic DNA of all isolates was extracted by a Wizard® genomic DNA purification kit (Promega). The 16S rRNA genes were amplified by the polymerase chain reaction (PCR) technique, using universal primers BF1 (5’-GAGTTTGATCATGGCTCAG-3’) and BR1 (5’-CGGTTACCTTGTTACGACTT-3’). The PCR products were purified by a GFX PCR DNA and gel band purification kit (Amersham). Sequencing reactions were performed with an ABI PRISM BigDye Terminator cycle sequencing ready-made kit (Applied Biosystem) (Srisukchayakul et al. 2007). The 16S rRNA gene sequences were edited and assembled with the BioEdit program (Hall, 1999). The nearly completed 16S rDNA sequences were compared with the sequences within the GenBank/EMBL/DDBJ databases using the BLAST search. The alignment of the 16S rRNA gene sequences of the isolates and related genera were carried out by using the CLUSTAL X program ver 1.83 (Thompson et al. 1997). Nucleotide substitution rates (Knuc values) (Kimura, 1980) were determined and a distance matrix tree was constructed by using the neighbor-joining method (Saitou and Nei, 1987). Alignment gaps and unidentified base positions were not taken into consideration in the calculations. The topology of the phylogenetic tree was evaluated by performing a bootstrap analysis with 1000 trials. Cultivation and crude extract preparation Ten milliliters of seed cultures (24 hrs) in SAP2 medium were transferred into 250-ml Erlenmeyer flasks containing 100 ml of four different cultivation media including modified Vy/2 medium (baker’s yeast paste, 5 g; seawater, 1 L) (Iizuka et al. 2003); modified RL 1 medium (peptone, 3 g; yeast extract, 2 g; KNO3, 0.5 g; vitamin B12, 0.5 µg/L; seawater, 1 L) (Spyere et al. 2003); modified CY medium (casitone, 10 g; malt extract, 2 g; yeast extract, 1 g; seawater, 1 L) (Suzuki et al. 1998) and SK medium (skim milk, 5 g; yeast extract, 3 g; MgSO4, 1 g; seawater, 1 L). Each flask contained two grams of amberlite XAD-16 resins used for absorbing of secondary metabolites produced during the cultivation (Spyere et al. 2003). The cultivating flasks were incubated and shaken at 25ºC; 200 rpm for 7 days. The amberlite XAD-16 resins were collected by filtering through a nylon mesh and washed with deionized water to remove salt and remaining cultivation media. The resins were set to dry at room temperature for 15 min and then soaked twice with 100 ml of methanol for 30 min. The methanol fractions were combined and evaporated to yield the dry crude extracts. Four different human cell lines including breast adenocarcinoma (MCF-7), colon cancer (HT-29), cervical cancer (HeLa) and oral cancer (KB) were used for the screening of cytotoxic activity using a sulphorodamine B (SRB) assay (Skehan et al. 1990). In brief, monolayer cultures of each cell line in 96-well microtiter plate (2 x 103 cells/well) were treated with marine gliding bacterial extracts dissolved in dimethyl sulfoxide (DMSO). The plate was incubated in a CO2 incubator at 37ºC for 7 days. At the mid point of cultivation, the medium was refreshed once. The percentage of survival of the cancer cells was measured colorimetrically by SRB dye as described by Skehan et al. (1990). The absorbance was measured by a microplate reader at the wavelength of 492 nm. One column of 96-well microtiter plate containing 2 x 103 cells/well of each cancer cell line was left untreated with a crude extract in order to use it as an internal control. Cytotoxicity was reported as the percentage of inhibition, calculated from the optical densities of treated wells and control, correlating to the viability of the cells as described in the equation below. % inhibition = [(OD492 Control - OD492 Sample) / (OD492 Control)] x 100 The DDBJ/EMBL/GenBank accession numbers for the 16S rRNA gene sequences of strains TISTR1761, TISTR1763, TISTR1768, TISTR1780 and TISTR1781 are AB433333, AB433334, AB444610, AB433335 and AB443430, respectively. Results and Discussion Isolation, 16S rRNA gene and phylogenetic analyses In this study, eighty-four marine gliding bacterial isolates were obtained from different specimens collected in the Gulf of Thailand and the Andaman Sea. It was interesting to note that biofilms and macro-algae were among the most prolific sources of marine gliding bacteria (data not shown). Gliding bacteria are known to play important roles in the biodegradation of organic matter in seawaters such as the exopolysaccharides commonly present in the biofilm and algae (Johansen et al. 1999). Moreover, biofilm which is a consortium of microorganisms could also serve as nutritional sources for certain gliding bacteria with predatory behavior (Kadouri and O’Toole, 2005). Initially all eighty four isolates were supposed to be screened for cytotoxicity using Glutamate (NH4NO3, 0.01 g; K2HPO4.3H2O, 0.01 g; sodium glutamate, 1.0 g; seawater, 1 L) and Glycerol (NH4NO3, 0.01 g; K2HPO4.3H2O, 0.01 g; glycerol, 1.0 g; seawater, 1 L) media for cultivation. However, the results showed that the crude extracts obtained from cultivation of forty nine isolates in these two media yielded no inhibitory activity against all four cancer cell lines which could be due to the compositions of the media. Therefore, Glutamate and Glycerol media were replaced with Vy/2, RL 1, CY and SK media for cultivation and only 27 isolates were selected for screening study based on their morphological characteristics. Almost complete 16S rRNA gene sequences of the twenty-seven selected strains and other neighboring species were used to construct the phylogenetic tree as shown in Figure 1. The results demonstrated that all marine gliding bacterial isolates belong to the Cytophaga-Flavobacterium-Bacteroides (CFB) group and can be further divided into six different groups. Group 1 (TISTR1761 = AB433333 and TISTR1763 = AB433334) formed a distinct lineage separated from the closest neighbor, Flavobacteriaceae bacterium (DQ423479), with only 90.0% sequence similarity. It is decidedly possible to consider this group as a novel genus. Cell morphology of TISTR1761 is shown as long threads (Figure 2a and Figure 2b). However, more descriptive details such as G + C mol content, biochemical and chemotaxonomic characteristics of this candidate novel genus are currently under investigation. The 16S rRNA gene sequences (4 strains) and the phylogenetic analysis of group 2 confirmed that all members were identical to Tenacibaculum mesophilum, a marine gliding bacterium recently proposed by Suzuki et al. (2001). The 16S rRNA gene sequences of group 3 (TISTR1780 = AB433335 and TISTR1781 = AB443430) showed 98% similarity with Fulvivirga kasyanovii (DQ836305) (Nedashkovskaya et al. 2007). The sequence similarity was low enough to hypothesize that this could potentially be a novel species of Fulvivirga.Group 4, which is the largest group, showed 16S rRNA gene sequence similarity close to Rapidithrix thailandica (AB284517), which was recently proposed by Srisukchayakul et al. (2007). However, the similarity between group 4 (TISTR1768 = AB444610) and Rapidithrix thailandica was only 98%. It is, therefore, possible that they are different species of Rapidithrix, hence DNA-DNA hybridization is needed for confirmation. Groups 5 and 6 were identified as Aureispira marina (Hosoya et al. 2006) and Aureispira maritima (Hosoya et al. 2007) respectively. The 16S rRNA gene sequence similarities of groups 5 and 6 with A. marina and A. maritima were 99.0% and 99.0%, respectively. Cell morphology of new species TISTR1781 and TISTR1768 are shown as long threads (Figure 2c and Figure 2f). The total of 108 crude extracts, obtained from the cultivation of each isolate in four different cultivation media, were submitted for cytotoxicity assay against four cancer cells lines including HeLa, HT-29, KB and MCF-7 using an SRB method. The results (Table 1) showed that 13 out of 27 isolates (48.1%) could produce the crude extract which exhibited cytotoxicity, when tested the crude extract at a concentration of 25 µg/ml in SRB assay. Interestingly, the crude extracts obtained from group 2 (Tenacibaculum mesophilum) and group 4 (Rapidithrix thailandica) were capable of inhibiting all four cell lines whereas only one member of group 6 (Aureispira maritima) produced the crude extract which was active against HeLa and HT-29. There is only one strain of R. thailandica which could produce the extract with cytotoxicity against MCF-7. In contrast, the extracts of group 1 (the candidate novel genus), group 3 (Fulvivirga kasyanovii) and group 5 (Aureispira marina) exhibited no cytotoxicity. However, there has been no report on the secondary metabolite production by T. mesophilum and other gliding bacteria phylogenetically related to the strains obtained in this study. This suggests that marine gliding bacteria are still understudied and they could be as a potential source of cytotoxic compounds, especially the genera Tenacibaculum and Rapidithrix.
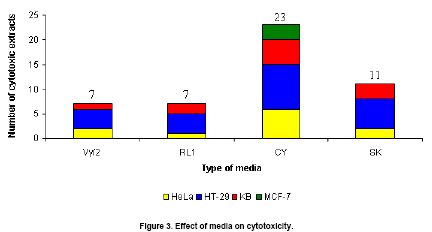
It is also important to note that the compositions of the cultivation media also played an important role in the production of the cytotoxic metabolites. From our observation, CY medium provided the faster growth of twenty-seven isolates than other cultivation media while Vy/2, RL 1 and SK media yield the same growth. As shown in Figure 3, CY medium yielded the highest number of crude extracts with cytotoxicity, followed by SK, RL 1 and Vy/2 media respectively. The CY medium was also the only medium which gave the crude extracts that could inhibit all four cell lines whereas other media could not yield the crude extract which inhibited MCF-7. This provided strong evidence that the media composition was one of the key factors for secondary metabolite production. Samarntarn et al. (1999) reported that amino acids and/or peptides in casitone, which was the key component in the CY medium, might serve as specific inducers for increasing the total alkaline protease production that causes the cytotoxic compound production. This study is the first to report the isolation of novel marine gliding bacteria in Thailand and screening for cytotoxicity. Acknowledgements Asst. Prof. Dr. Supreeya Yuenyongsawasdi and Mr. Theerasak Anantapong for their assistances with the cytotoxic assay. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2009 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej09023f2.jpg] [ej09023f3.jpg] [ej09023f1.jpg] [ej09023t1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}