 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 12, No. 3, July 15, 2009 Group III PLA2 from the scorpion, Mesobuthus tamulus: cloning and recombinant expression in E. coli Gururao Hariprasad1 , Kolandaivelu Saravanan2, Sundararajan Baskar Singh3, Utpal Das4, Sujata Sharma5, Punit Kaur6, Tej Pal Singh7, Alagiri Srinivasan*8,
1Department of Biophysics,
All India Institute of
Medical Sciences,
Ansari Nagar, New Delhi
- 110 029,
India Financial support: Department of Science and Technology as a grant to GH (No. SR/FT/L-18/2005). Code Number: ej09026 Phospholipases A2 (PLA2) are enzymes that specifically hydrolyze the sn-2 fatty acid acyl bond of phospholipids, producing a free fatty acid and a lyso-phospholipid. We report the cloning and expression of a secretory phospholipase A2 (sPLA2) from Mesobuthus tamulus, Indian red scorpion. The nucleotide sequence codes for a 167 residue enzyme. The open reading frame codes for a 31 amino acid signal peptide followed by a mature portion of the protein. The primary structure shows the calcium binding motif, catalytic residues, 8 highly-conserved cysteines and C-terminal extension which classify it as a group III PLA2. The entire transcript was expressed in Escherichia coli and was purified by metal affinity chromatography under denaturing conditions. The protein was refolded by serial dilutions in the refolding buffer to its active form. Hemolytic assays indicate that the protein adopts a functional conformation. The functional requisites such as optimum pH of 8 and calcium dependency are shown. This report provides a simple but robust methodology for recombinant expression of toxic proteins. Keywords: group III phospholipase A2, Mesobuthus tamulus, recombinant expression.
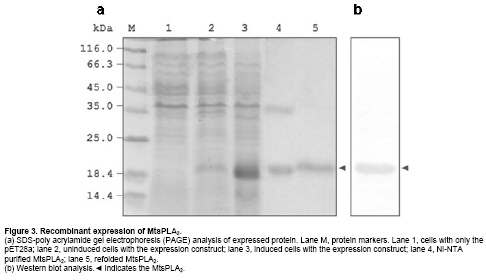
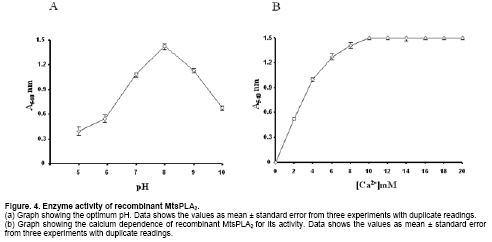
Phospholipase A2 (PLA2) (EC 3.1.1.4) catalyzes the hydrolysis of glycerophospholipids at sn-2 position producing free fatty acids and lysophospholipids. One such reaction product arachidonic acid is the precursor in the biosynthesis of prostaglandins. Prostaglandins are important in the inflammatory process and are implicated in numerous pathophysiological conditions. It, therefore, holds importance in the field of biology and medicine (Kaiser, 1999). PLA2 is a widely distributed enzyme across the species and classified into ten groups on the basis of their sequence homology and disulfide pattern (Six and Dennis, 2000). Group I and group II phospholipase A2s have been well characterized in terms of their structure and function. They share sequence and structural similarity among themselves. The other groups have not been so well characterized. Many group III PLA2s have been identified across the species: insect (Kuchler et al. 1989; Scott et al. 1990; Ryu et al. 2003), scorpion (Zamudio et al. 1997; Valdez-Cruz et al. 2004; Hariprasad et al. 2007), gila monster (Gomez et al. 1989) and human (Valentin et al. 2000). Though they share many enzymatic properties, they do not share any significant amino acid homology. Apart from their involvement in inflammation (Touqui and Alaoui-El-Azer, 2001; Murakami et al. 2003), they have been implicated in apoptosis (De Coster, 2003) and cancer (Murakami et al. 2005) necessitating the study of similar secretory phospholipase A2 (sPLA2s). In the context of heterologous gene expression being very useful in modern biotechnology (Manzur et al. 2006), the aim of the present work was to clone and recombinantly express in E. coli a group III PLA2 from the scorpion, Mesobuthus tamulus. Materials and Methods Venom glands were removed from ten Indian red scorpions obtained from Irula Snake Catchers Industrial Society Limited, with the permission of the Department of Forests and Wild life, Government of Tamil Nadu, India. The total RNA was isolated using guanidium isothiocyanate (Chomczynski and Sacchi, 1987). cDNA library was prepared in pBluescriptSK+ using cDNA synthesis kit according to the manufacturer’s protocol (Amersham Biosciences, Hong Kong). Forward primer 5’-ATGCATACTCCAAAACACGCTAT-3’ was designed based on the cDNA sequence of imperatoxin I (Zamudio et al. 1997) and used in conjunction with oligo (dT) primer to amplify PLA2 gene from the cDNA library. Polymerase chain reaction (PCR) was performed with thermal cycler instrument (PTC 100, USA) for 30 cycles of 1 min at 94ºC, 2 min at 54ºC, 2 min at 72ºC followed by a final extension of 10 min at 72ºC. The amplified product was isolated and cloned into pGEMT-Easy vector (Promega, USA) and both strands from at least four clones were sequenced by dye termination method using automatic sequencer (ABI, USA). The homology search using BLAST, alignment using ClustalW and furin-type cleavage site prediction (Duckert et al. 2004) on ExPASY on line were carried out. Construction of expression plasmid From the complete PLA2 sequence obtained, a new forward primer 5’-ACTATGTGGGGAACCAAATGGTG-3’ corresponding to the N-terminus of mature protein and a reverse primer, 5’-TTAACCATTCAGTTTGTTAATCGC-3’, corresponding to the C-terminal end of the protein was synthesized to amplify the mature MtsPLA2. The amplified product was cloned in pGEMT-Easy vector to obtain pGEMT-MtsPLA2. The plasmid pGEMT-MtsPLA2 and expression vector pET28a were digested with NotI restriction enzyme. After dephosphorylation of the expression vector with calf intestinal alkaline phosphatase for 1 hr at 50ºC, the insert was ligated into pET28a to obtain an expression construct, pET28a-MtsPLA2 which was used in the transformation of E. coli BL21 (DE3) cells for protein expression. The pET28a-MtsPLA2 transformant was grown in luria-bertini (LB) medium containing 100 µg/ml ampicillin at 37ºC to an A600 of about 0.6 and iso propyl thio-β-D-galactopyranoside (IPTG) was added to a final concentration of 1 mM for induction. Cells were incubated for an additional 4 hrs at 37ºC and harvested by centrifugation for 15 min at 8000 g. The cell pellet was suspended in buffer A (25 mM Tris-HCl buffer, pH 8.0 and 100 mM NaCl) containing 1 mM ethylene diamine tetra acetic acid (EDTA), 1 mM phenyl methane sulphonyl fluoride (PMSF), 50 μg/ml DNaseI, 20 μg/ml RNaseA and 1 mM MgCl2 and sonicated for 15 min. The inclusion body obtained by centrifugation at 20,000 g at 4ºC for 30 min was washed with buffer A, containing 2 M urea and solubilised with buffer B (25 mM Tris-HCl buffer, pH 8.0, 100 mM NaCl and 8 M urea) with continuous stirring at room temperature for 4 hrs. After centrifugation, the supernatant solution was applied to pre-equilibrated nickel nitrilo tri aceticacid (Ni-NTA) resin column (Qiagen, Germany). The column was washed with 25 mM immidazole in buffer B and the protein was eluted with 300 mM immidazole in buffer B. The eluted protein was refolded by step-wise dialysis against buffer A containing decreasing concentrations of urea starting from 7 M to 0 M. 5% (v/v) β-mercaptoethanol was present during this urea removal step. The β-mercaptoethanol was removed by dialyzing against buffer A, containing reduced (1 mM) and oxidized (0.1 mM) glutathione. Each round of dialysis was done at 16ºC for 8 hrs. The dialyzed sample was centrifuged at 12,000 g at 4ºC for 15 min and the supernatant was lyophilized. The protein quantity was determined by Bradford colorimetric assay. Sodium dodecyl sulphate-polyacrylamide gel electrophoresis and Western blot Samples were run on 12% poly-acrylamide gel under reducing conditions using Lammeli system of buffers and stained with coomassie blue. The refolded His-MtsPLA2 protein was transferred to a nitrocellulose membrane. The membrane was blocked with 3% bovine serum albumin in PBS for 1 hr at room temperature and incubated with horse radish peroxidase (HRP)-conjugated Ni-NTA (Qiagen, Germany) diluted in transfer solution (1 : 1000) for 1 hr. The nitrocellulose membrane was washed four times with PBS containing 0.05% Tween20. The protein was localized using 2 mM 3, 3’-diaminobenzidine and 0.1% hydrogen peroxide in 0.1 M Tris-HCl, pH 7.5. The pH dependence of MtsPLA2 activity was assayed by RBC lysis method (Shiloah et al. 1973). 106 RBCs were resuspended in 1 ml of 0.15 M NaCl in eppendorf tubes. Different buffers (100 mM glycine-HCl, pH 5.0 and pH 6.0; 100 mM Tris-HCl, pH 7.0 and pH 8.0; 100 mM glycine-NaOH, pH 9.0 and pH 10) were used to maintain different pHs. For controls, the RBCs were suspended in respective buffered saline solution without the enzyme. 10 µg of MtsPLA2 was used for hydrolysis. The tubes were put on end-over-end shaker at 25ºC for 5 min. After the incubation, the tubes were centrifuged at 5000 g at 25ºC for 10 min. Activity was determined by the relative amount of hemoglobin liberated from the RBC by measuring the supernatant color at 540 nm. Calcium dependence was determined using the same set of experimental conditions with different concentrations of CaCl2 (0-20 mM) at pH 8.0. The nucleotide sequence data reported here is available in EMBL, GenBank and DDJB databases (GenBank accession number: AY443497). Results and Discussion Gene specific forward primer from the nucleotide sequence of imperatoxin I (Zamudio et al. 1997) was used for the amplification of M. tamulus venom gland cDNA library to get a 656 bp gene. This was the only scorpion PLA2 nucleotide sequence available in the literature when this work was started. Oligo (dT)25N was used as the reverse primer. Thus, the first 23 nucleotides in the sequence are primer dictated. The salient features of the nucleotide sequence are given in Figure 1. Blast analysis did not show any significant identity (> 30%) at the nucleotide level with any group of PLA2. Complete degeneracy is observed even at the conserved amino acid positions. The only exception is the 85% identity it shares with the cDNA sequence of imperatoxin I of Pandinus imperator (Zamudio et al. 1997). The families, Buthidae and Scorpionidae to which Mesobuthus tamulus and Pandinus imperator belong, are closely related phylogenetically (Petrunkevitch, 1955). Amino acid sequence homology search for the translated protein indicates that the amplified product codes for group III sPLA2. Sequence comparison with other selected group III sPLA2s showed conservation of all the essential motifs (Figure 2). The calcium binding residues Trp7, Gly9, Gly11, Asp34 and the His/Asp dyad at position 33/34 is conserved and preceded by Asp-X-Cys-Cys-Arg-X motif in all the sequences. A key residue aspartic acid which is needed to stabilize the catalytic histidine in most sPLA2s is replaced by glutamic acid at position 62 in MtsPLA2, as in imperatoxin I (Zamudio et al. 1997) and D. melanogaster sPLA2 (Ryu et al. 2003). This homologous amino acid change has not resulted in the loss of enzymatic activity. Another highlight of the homology study is that the first eight of the ten cysteines are conserved in all group III sPLA2s while the last two at the C-terminal extension are at less conserved positions. It appears that all native scorpion PLA2s have C-terminal extensions which are subsequently cleaved and held by disulfide linkages to form heterodimers (Zamudio et al. 1997; Conde et al. 1999; Valdez-Cruz et al. 2004). This type of sub unit association is similar to that seen in β-bungarotoxins of snake PLA2s (Wu and Chang, 2000) but differ in two aspects: (1) the entire MtsPLA2 is the product of a single transcript and (2) the second subunit has no sequence similarity with any of the known proteins. Recombinant expression and enzymatic assay Having deciphered the structural features, it was important to know if this protein can be expressed in its functional form. MtsPLA2 gene was subcloned in pET28a expression vector. Electrophoretic analysis showed leaky basal expression in the uninduced cells and an expression of 20% in the induced cells. The protein was present in inclusion body. Lowering the inducer concentrations or the growth temperature did not produce the soluble enzyme. The inclusion body was solubilized using 8 M urea and the 6 x His-Tag-MtsPLA2 was purified by metal chelation chromatography under denaturing conditions. The purified protein existed as homodimer which is known to occur when inclusion bodies are solubilized with strong denaturants (Sambrook et al. 2001). The protein was refolded by stepwise removal of urea by dialysis. A large portion of the protein aggregated from the accumulation of high concentrations of folding intermediates during the process. The final yield of the purified active protein was 2.8 mg/liter as estimated by weight basis. The complete purification profile of recombinant His-MtsPLA2 is shown in Figure 3. His-MtsPLA2 migrated as a single protein of ~19 kDa on sodium dodecyl sulphate (SDS) gel under reducing conditions, indicating the existence of the entire protein as a single unit and not as a heterodimeric protein. During the bacterial expression, the entire transcript is expressed as a single protein. The furin-type cleavage required for the production of second small peptide unit has not taken place (Lane 5, Figure 3). As the enzyme is not cleaved and is present as a single molecule, the theoretical molecular weight and pI of the expressed MtsPLA2 including the connecting pentamer and vector dictated sequence is 19657 Da and 8.28 respectively. There is no experimental evidence for the heterodimeric form of the expressed protein. Under conditions of correct post translational processing, the expressed protein could give rise to a heterodimeric protein. The evidence for this conclusion comes from the amino acid sequence homology with other scorpion PLA2s (Zamudio et al. 1997; Conde et al. 1999; Valdez-Cruz et al. 2004) which all exist as a heterodimeric form in the native state. The optimal pH and calcium dependence for the refolded enzyme was determined by RBC lysis. The enzyme has an optimum pH of 8.0 (Figure 4a) and was calcium dependent showing a maximum activity with 10 mM of calcium (Figure 4b). These results are concurrent with previous results (Valentin et al. 2000; Valdez-Cruz et al. 2004). It is not determined if the observed hemolytic activity was a direct or an indirect effect. But previous studies have shown that the scorpion PLA2s can have direct hemolytic activity (Valdez-Cruz et al. 2004). We have determined the primary structure of phospholipase A2 from Indian red scorpion, Mesobuthus tamalus. The sequence analysis reveals that despite variation, the sequence is conserved with respect to the catalytic activity, calcium binding and structural integrity. Our protein-expression results indicate that the separation of the second peptide chain is not critical to the functional status of the enzyme. It is also established that the folding of PLA2 enzymatic domain is not affected by the presence of second peptide chain. Acknowledgements The technical assistance of Biomedical Informatics Center at AIIMS supported by Indian Council of Medical Research, Government of India, is acknowledged. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network © 2009 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej09026f4.jpg] [ej09026f3.jpg] [ej09026f1.jpg] [ej09026f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}