 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 1, Jan, 2010 Research Article Genotypic variation in morphology and freezing resistance of Eucalyptus globulus seedlings subjected to drought hardening in nursery Rafael E. Coopman*1 , Jorge C. Jara2, Rene Escobar3, Luis J. Corcuera4, Leon A. Bravo5 1Instituto de Silvicultura,
Facultad de Ciencias Forestales y Recursos Naturales,
Universidad Austral de
Chile,
Valdivia, Chile
E-mail: rafael.coopman@docentes.uach.cl Received March 20, 2009 / Accepted August 31, 2009 Code Number: ej10005 Abstract Eucalyptus globulus Labill is one of the most planted species in Chile, because of its fast growth and superior pulp qualities. Nevertheless, the incidence of drought and frost damage immediately after planting is frequent. The purpose of this work was to study the effect of drought hardening on frost resistance and on variations in morphological traits that may increase drought resistance at nursery phase in four genotypes of E. globulus Labill. Drought hardening treatments consisted in induced water stress by watering restriction, until pre-dawn stem xylem water potentials (Ψpd) reached -0.2, -1.8 and -2.6 MPa. Two water stress-rewatering cycles were applied during 54 days of hardening. Plant and root biomasses were affected by the interaction of drought hardening and genotypes. The rest of morphological and alometrical traits were affected independently by drought or genotype. Plant height, leaf area, specific leaf area (SLA), stem, and leaf biomasses decreased with drought hardening, while collar diameter was not affected. Genotypes responded differentially to drought hardening in plant height, leaf area, SLA, and stem, and leaf biomasses. Ice nucleation temperature (INT), and freezing temperatures (FRT), and 50% freezing damage index of leaves (LT50) were affected by the interaction between drought hardening and genotypes. EG-13, EG-23 and EG-22 genotypes became freezing tolerant with drought hardening (-2.6 MPa). Additionally, EG-14 genotype increased its freezing resistance at -1.8 MPa. Therefore, freezing resistance levels and mechanism depend on genotype and drought hardening treatment. The success in tree breeding by genetic selection should be facilitated by improved understanding of the physiology of stress resistance development and survival during water supply limitations. The knowledge of morphological and freezing resistance dependency on the interaction between genotype and drought hardening may be useful nursery management information to improve plantation success. Keywords: biomass allocation, cross hardening, genotypic selection, planting stock quality, water stress.
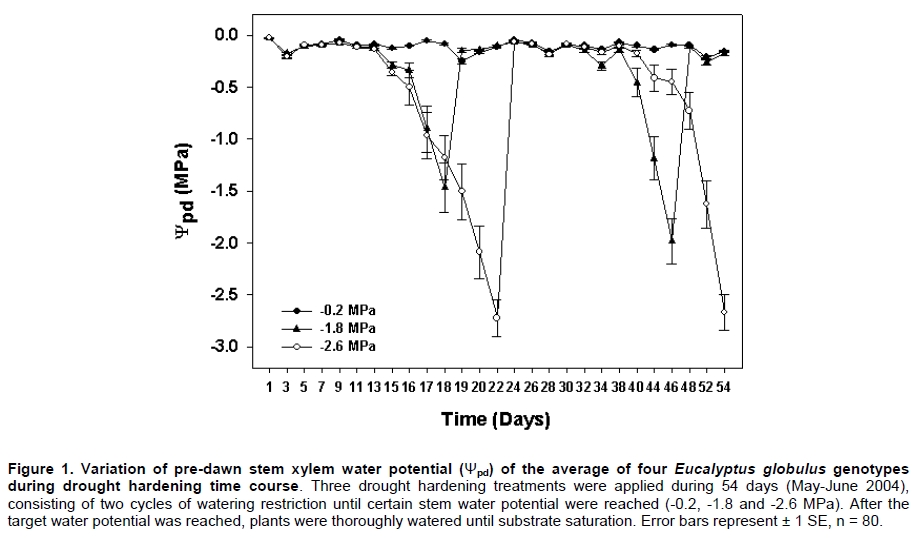
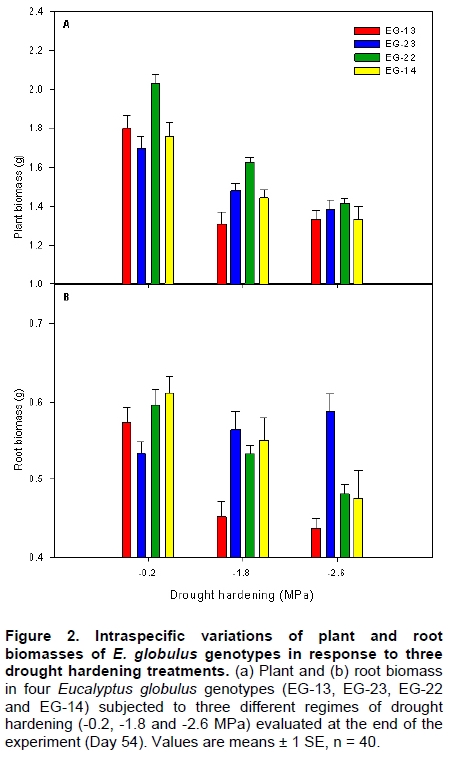
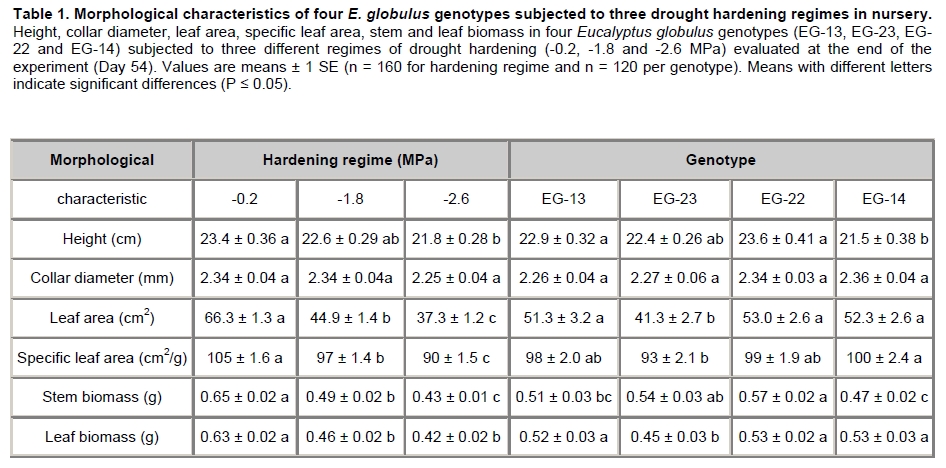
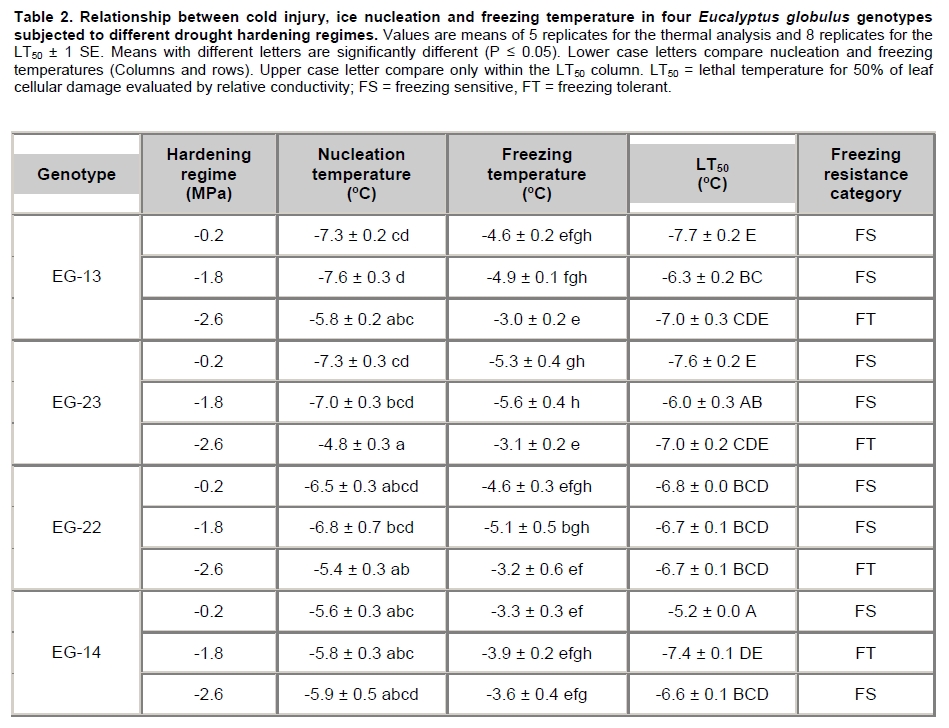
Eucalyptus globulus Labill is one of the most planted species in Chile, because of its fast growth and superior pulp qualities. However, seedlings are inherently susceptible to drought, low temperature, and frost. These factors largely contribute to seedling mortality soon after planting (Close et al. 2004). The challenges to improve E. globulus capacity to withstand drought and freezing temperatures by resistant genotypes selection have received major attention (Pita and Pardos, 2001; Costa e Silva et al. 2007; Wikberg and Ögren, 2007). Chilean forest companies have expanded and developed intensively in the last two decades. This has led to tree forest nurseries to incorporate the most modern container nurseries and vegetative plant propagation procedures. Plants develop succulent stem and leaves and high shoot:root ratios in response to the high nitrogen fertilization rates used during the full growth phase in nursery. These features make seedlings highly susceptible to drought and frost damage (Thomas and Ahlers, 1999; Sundheim Fløistad and Kohmann, 2004; Villar-Salvador et al. 2004b). The final drought hardening phase under nursery conditions induces leaf morphological and seedling biomass allocation modifications which are closely linked with the survival of plants in unfavourable sites with seasonal drought (Royo et al. 2001; South et al. 2001; Coopman et al. 2008). Seedlings exposed to sub-lethal water stress cycles in nurseries exhibit less drought injury in field than non stressed plants (van den Driessche, 1992; Villar-Salvador et al. 2004a). Changes in drought tolerance depend on the degree of water stress, the length of the hardening period, and the species (Royo et al. 2001; Wikberg and Ögren, 2007). There is limited information about morpho-physiological variations induced by a drought hardening phase under nursery conditions of E. globulus genotypes adapted to ambient conditions in Chile (Coopman et al. 2008). Plants respond to water deficit by reducing growth rates and water loss. During acclimation to water stress, adjustments in architecture and intrinsic physiology can occur (Ewers et al. 2000). Leaf shedding and root growth triggered by water stress (Linder et al. 1987) result in decreased shoot:root ratio, which favours a high water balance during drought (Ewers et al. 2000; Hsiao and Xu, 2000). Plant morphological attributes such as collar diameter and plant height have been consistently reported as good indicators for survival and growth potential after planting (Mason, 2001; Haase et al. 2006). Finally, there is evidence that cold acclimation and water stress induce cell dehydration. This water loss triggers a series of common physiological responses, such as accumulation of solutes (Beck et al. 2004). These solutes decrease protoplast osmotic potential and may cryoprotect freezing labile cell structures. Furthermore, products of drought induced genes may also be involved in freezing resistance (Pearce, 2001; Beck et al. 2007). Development of cold hardiness is initially related to a reduction in water content and then to changes in the physical state of water (Shvaleva et al. 2008). Solutes that accumulate during water stress may bind to water, affecting even more the physical properties of water. It has been proposed that drought resistant genotypes of Eucalyptus globulus have a greater cold tolerance than drought susceptible ones (Costa e Silva et al. 2009). Moreover, physiological and morphological changes are differentially induced in different genotypes (Pita and Pardos, 2001). Differences in leaf anatomy influenced the formation of ice and the pattern of frost injury in three evergreen tree species from New Zealand (Neuner and Banister, 1995). Also, genetic variation in frost resistance of Eucalyptus globulus have been reported (Tibbits et al. 2006). The purpose of this work was to study the effect of drought hardening on frost resistance and on variations in morphological traits that may increase drought resistance at nursery phase in four genotypes of E. globulus Labill. Materials and Methods A total of 11,200 eight month old rooted cuttings of Eucalyptus globulus Labill, divided in 4 sets of 2,800 plants of each genotype evaluated (EG-13, EG-23, EG-14 and EG-22), were obtained from the Forestry Company Bosques Arauco S.A. Chile. Cuttings were rooted in trays of 88 containers of approximately 130 ml each, using a mixture of turf:perlite 1:1 (v/v) with a total porosity, aeration and water retention of 75%, 32%, and 42% respectively. The growth density was 393 plants m-2, in a greenhouse with mist irrigation. After rooting, plants were grown outdoors for 5 months. For drought hardening, these plants were kept under a transparent roof in order to avoid water from rainfall. Daily maximum and minimum air temperatures and relative humidity, wind speed, and maximum PPFD were determined at foliage level with a weather station (Datalogger 21 X, Campbell Scientific Inc., Logan, Utah) with temperature and relative humidity sensor (CS500-L, Campbell Scientific Inc., Logan, Utah), and an anemometer (Vector Instruments Type A 100 L2, N. Wales, United Kingdom) and piranometer (LI-COR LY-200 Biosciences, Lincoln, NE USA). Three different treatments were applied and consisted in watering restriction until plants reached a pre-dawn stem xylem water potential of (Ψpd:-0.2, -1.8 and -2.6 MPa). Plants were then watered to reach substrate saturation and a second cycle of drought started. The treatment with Ψpd = -0.2 MPa was considered as a non-stressed control; the -1.3 MPa (moderate stress) value is in the center of the target range (-1.0 to -1.5 MPa) for drought hardening processes that has been commonly used in most USA forest nurseries (Landis, 1989). It is just below to the wilting point of the experiment’s seedlings. Finally, -2.6 MPa (severe stress) corresponds to a permanent wilting point of experiment’s seedlings. The hardening phase lasted 54 days (May-June 2004) and included two cycles of low water potential (Figure 1). Ψpd was measured in plants of each genotype and treatment (n = 20) with the Scholander pressure chamber (Soilmoisture Equipment Corp, Santa Barbara, CA, USA), approximately every two days. The treatment with Ψpd = -0.2 MPa was watered with a frequency of about 7 days. Ψpd did not change until day 13 and then decreased at a rate of -0.27 MPa day-1 (Figure 1). The desired water potentials were reached at 18 and 22 days in the first cycle, respectively. After rewatering, the plants reached again the desired water potentials after 46 and 54 days, respectively (Figure 1). In order to uniform the humidity content of the substrate, before the beginning of the assay and immediately after the hardening treatment, plants were all watered as treatment -0.2 MPa. Non significant differences in the Ψpd behaviour between genotypes were found during hardening (P ≤ 0.05). Forty plants from each treatment and genotype were collected at the end of the assay to determine plant height, stem diameter, and root, stem, leaf, and plant dry biomasses. Additionally, leaf area and specific leaf area were determined using a leaf area meter Li-3100 (Li-Cor Inc., Lincon, Ne USA). Three weeks after the end of the hardening phase, 12 plants from each hardening treatment and clone were collected. Two leaf disks (8 mm diameter) were obtained from each third pair of leaves, which were the first totally expanded leaves. The leaf disks were placed into polycarbonate test tubes and exposed to freezing temperatures (-3.0º, -4.5º, -6.0º, -7.5º, and -9.0ºC), without ice nucleating agents in a chamber cooled by forced air. The above zero cooling rate was 30ºC h-1 and below cero 2.1ºC h-1. The time of exposure to each target freezing temperature was 1.5 hrs. Then, leaf disks were left to thaw at 4ºC in darkness for 24 hrs. After this period the leaf injury index was evaluated (LT50) in leaf disks by measuring changes in relative electrolyte conductivity caused by ion leakage (RC) according to Raymond et al. (1986). Thermal analyses were conducted with material from the same leaves in order to determine ice nucleation temperature (INT) and freezing temperature (FRT) (Larcher, 2003). With these values and LT50, it was possible to determine if a particular genotype was freezing sensitive (FS) or freezing toletant (FT). If ice nucleation temperature is higher than LT50, then the genotype is FT. Similarly, if ice nucletion temperature is lower than LT50, the genotype is FS. Essentially, this procedure was the same as used by Reyes-Díaz et al. (2005). Briefly, a piece of leaf was attached to a fine copper-constantan type T thermocouple, gauge 30, junction diameter about 0.5 mm (Cole Palmer, Vernon Hills, IL USA). This sample was immediately enclosed in a cryotube to avoid tissue dehydration. Cryotubes were immersed in a cryostat (C25P, ThermoHaake, Karlsruhe, Germany) programmed to perform a linear descend from 0º to -15ºC at a rate of 2ºC h-1. Temperature was continuously monitored and recorded every 1 sec with a USB data acquisition system (Personal Daq/56, IOtech. Inc, Cleveland, Ohio USA) connected to a personal computer. There were no genotypic differences between Ψpd during hardening. The variation in Ψpd of the average of genotypes (n = 80; 20 by each genotype) was considered for the enforcement of drought hardening treatments. A factorial randomized block designs was use to examine the effect of drought hardening treatments(-0.2, -1.8 and -2.6 MPa) and genotypes (EG-13, EG-23, EG-22 and EG-14) on morphological characteristics (n = 160 for hardening regime and n = 120 per genotype), and intraspecific variations of plant and root biomasses (n = 40), and cold injury, ice nucleation, and freezing temperatures (n = 5 for thermal analysis and n = 8 for the LT50). The factorial design was arranged in 4 spatial blocks (culture platforms) with 233 replicates (seedlings) of each combination of drought hardening treatmentsand genotypes. The experimental unit was the seedling. The sampling within each block was totally random. Data were analyzed by two-way ANOVA. All variables were subjected to normality and variance homogeneity tests prior to ANOVA. A post-hoc Tukey test was applied when ANOVAS gave significant differences (P ≤ 0.05). All these analyses were performed using the software STATISTICA (Version 6, 2001, StatSoft, Tulsa, OK). Results Drought hardening treatments generated differential genotypes effects on seedlings allometry. At the end of the experiment, the interaction between genotypes and hardening treatment was significant only for plant (P ≤ 0.05) and root biomass (P ≤ 0.01). Plant biomass decreased in the four genotypes as the stress treatment increased. However, there were no significant differences in plant biomass between genotypes hardened at -1.8 and -2.6 MPa (Figure 2A). The plant biomass of the EG-22 genotype only differed between the -0.2 and -2.6 MPa treatments. The plant biomass of EG-22 genotype hardened at -0.2 MPa was higher than the rest (Figure 2A). In the -1.8 MPa drought hardening treatment the EG-22 and EG-13 genotypes generated the highest and lowest plant biomass, respectively. The root biomass of EG-13 genotype showed the highest sensitivity to water stress, diminishing 22% in the -1.8 MPa treatment. The root biomass of EG-22 and EG-14 genotypes decreased only with the most severe hardening treatment (-2.6 MPa). The genotypes root biomass did not differ within the control treatment (-0.2 MPa). However, root biomasses of almost all genotypes differed with the hardening treatments at -1.8 and -2.6 MPa. The EG-23 genotype showed a different trend: it’s root biomass increased with the exposure to the most severe water stress treatment (Figure 2B). Mortality of plants by dehydration was low and did not differ among genotypes, but it was significantly different within treatments (P ≤ 0.01). Plants exposed to -1.8 and -2.6 MPa only exhibited a mortality of 2.5% and 5.7%, respectively. The hardening treatments resulted in a significant decrease in plant height, leaf area, SLA, stem, and leaf biomasses, while collar diameter was not affected. The genotypes were not different in size at the beginning of the essay. However, they showed differential responses to drought hardening treatments in plant height, leaf area, SLA, and stem, and leaf biomasses (Table 1). The interaction between drought hardening treatment and genotypes was significant for ice nucleation, freezing temperatures (P ≤ 0.05) and LT50 (P ≤ 0.01). Hardening treatments at -0.2 and -1.8 MPa did not modify ice nucleation and freezing temperatures of EG-23 and EG-13 genotypes. However, both temperatures increased with the -2.6 MPa treatment in EG-13, EG-23 and EG-22 genotypes. These temperatures did not change with the hardening treatment in the EG-22 y EG-14 genotypes, with the exception of freezing temperature of EG-22 that was significantly reduced in the plants exposed to -1.8 MPa. Ice nucleation temperatures were significantly lower than freezing ones in all cases, except for the EG-22 clone after hardening at -1.8 MPa (Table 2). The LT50 of EG-13 and EG-23 genotypes at -0.2 and -2.6 MPa were similar and lower than at -1.8 MPa. The -1.8 and -2.6 MPa drought hardening treatments reduced the LT50 of EG-14 genotype, while the EG-22 genotype LT50 was not affected. When comparing genotypes in the -0.2 MPa drought hardening treatment, the genotypes decreased their freezing resistance in the following order EG-14, EG-22, EG-23 and EG-13. However, within genotypes hardened at -1.8 MPa, EG-14 showed the lowest LT50. The LT50 of all genotypes subjected to -2.6 MPa did not differ (Table 2). Water stress effects on growth and morphology Different intraspecific morphological responses to water stress have been reported in E. globulus seedlings (Osorio and Pereira, 1994; Costa e Silva et al. 2004). Drought hardening treatments, -1.3 MPa and -2.4 MPa, applied under nursery conditions, significantly reduced plant height, leaf area, and specific leaf area, stem and leaf biomass in all evaluated E. globulus genotypes. Plant height, which is considered a predictor of growth potential in the field, was reduced 3% and 7% in treatments at -1.8 MPa and -2.6 MPa respectively. Nevertheless, excessive plant height is a disadvantage in water stressed sites (South and Zwolinski, 1997). A quality seedling planting stock is composed of the biggest possible seedlings which maintain acceptable levels of survival in a designated site (South et al. 2005). Collar diameter of nursery seedlings, which is considered the best predictor of survival and initial growth of plants in a plantation (South et al. 1995), did not differ within our drought hardening treatments neither among genotypes. The low responsiveness found in this morphological trait, could be beneficial because genotypes can be hardened without a trade off with morphological indicators of seedling quality. However, collar diameter can be effectively controlled by plant density (Mason, 2001). A particularly efficient mechanism to reduce the transpiratory surface is partial leaf abscission (Larcher, 2003). Drought hardened genotypes at -1.8 and -2.6 MPa reduced their leaf area in a 32% and 44%, respectively. This variable is one of the most sensitive ones to water stress as reported by several studies in E. globulus (Osorio et al. 1998; Pita and Pardos, 2001; Costa e Silva et al. 2004). During our experiments, abscission of a significant foliage fraction (old and basal leaves) was observed, which is a known strategy to face drought in E. globulus (Coopman et al. 2008). The EG-23 genotype showed a lower leaf area than the other genotypes. According to Pita and Pardos (2001), the big leaf genotypes are of high risk for plants in drought prone areas. Drought hardening treatments of -1.8 MPa and -2.6 MPa significantly reduced SLA, 8% and 14%, respectively. Similar results have been reported by Pita and Pardos, (2001); Costa e Silva et al. (2004). For E. globulus genotypes, the maintenance of the balance between transpiration and absorption areas when water availability is reduced seems to be a determinant key of seedlings performance under drought conditions (Hsiao and Xu, 2000; Costa e Silva et al. 2004; McDowell et al. 2008). Finally, the biggest root biomass, smaller leaf area LA, and SLA of the EG-23 genotype are characteristics that would indicate a bigger capacity to tolerate drought relative to the other genotypes. In general, these changes are consistent with a higher water stress resistance after the acclimation period. Plants were re-watered after the acclimation period, but the morphological changes remained. For this reason, this material was considered fit to show that freezing resistance would be affected by morphological changes generated by water stress. Thermal analyses combined with a freezing injury index (LT50) allowed us to predict that the studied genotypes, at unstressed conditions, were not able to tolerate ice formation within their leaf tissues, showing incipient damage (LT10) at higher temperatures that INT (data not shown). For this reason, we classified these 4 E. globulus genotypes as freezing sensitive. Nevertheless, the severe drought hardening treatment (-2.6 MPa) increased the ice nucleation temperature of EG-13, EG-23 and EG-22 genotypes, resulting in an LT50 lower than INT. This indicates that these genotypes became freezing tolerant with drought hardening. Commonly, the initial and main frost damage mechanism in cold exposed plants is cellular dehydration, induced by aploplastic water freezing and its consequences (Neuner and Bannister, 1995). Since ice crystals are formed outside the cell, freezing and drought are considered causal of cellular dehydration (Shinozaki and Yamaguchi-Shinozaki, 2000; Beck et al. 2007). Then, it is reasonable to suggest that similar modifications are required in the cell to tolerate both stresses (Beck et al. 2007). Consistently, we detected a change in the freezing resistance mechanism in EG-13, EG-23 and EG-22 genotypes. However, their LT50 were minimally modified by the water stress acclimation period. An exception was EG-14, which increased its LT50 at the medium stress level. Therefore, freezing resistance depends on genotype and water stress level, as in other tree species (Costa e Silva et al. 2007; Costa e Silva et al. 2009; Gimeno et al. 2009). This physiological variation of the stress-response of genotypes is important for the development of breeding programmes and is essential to support decisions to allocate genotypes to different climatic planting sites (Shvaleva et al. 2008). In reforestation, seedlings are usually planted in autumn to avoid drought, but this may render the seedlings susceptible to autumn and winter frosts. Measuring freezing resistance 3 weeks after water stress was relieved appears to be an appropriate way to find semi-permanent changes induced by water stress. Water stress metabolites decrease during the deacclimation period (Dichio et al. 2006; Shvaleva et al. 2006; Dichio et al. 2009), but the more permanent seedlings morphological changes remain. Increasing drought resistance before plants are transferred from nurseries to the field seems a useful nursery silvicultural practice to improve plantation success. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10005f1.jpg] [ej10005t2.jpg] [ej10005f2.jpg] [ej10005t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}