 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 1, Jan, 2010 Research Article Normalized embryoid cDNA library of oil palm (Elaeis guineensis) Pek-Lan Chan1* , Lay-Sun Ma2* , Eng-Ti L. Low3*, Elyana M. Shariff4, Leslie Cheng-Li Ooi5, Suan-Choo Cheah6, Rajinder Singh*7 1Advanced Biotechnology and Breeding
Centre,
Malaysian Palm Oil Board,
No. 6, Persiaran Institusi,
Bandar Baru Bangi,
43000 Kajang, Selangor, Malaysia Note: The sequences have been deposited in GenBank. Accession numbers: DW247764-DW248770. * These authors contributed equally to this work. Financial support: Ministry of Science, Technology and Innovation (MOSTI), Malaysia under the Malaysia-MIT Biotechnology Partnership Programme (MMBPP). Code Number: ej10010 Abstract A normalized embryoid cDNA library (EON) was constructed based on reassociation kinetics reaction. Results from dot blot hybridization and sequencing of EON cDNA clones clearly indicated that the normalization process reduced the frequency of high abundance transcripts and increased the frequency of low abundance gene transcripts. A total of 553 non-redundant expressed sequence tags (ESTs) were identified, 325 of these were not observed in the standard oil palm cDNA libraries sequenced previously. A total of 10 EON cDNA clones were chosen for expression profiling across samples from different stages of the tissue culture process. Two of the genes exhibited promising expression patterns for predicting the embryogenic potential in callus. Some of these genes were also differentially expressed in the various tissues of oil palm. This study showed that normalization of the existing embryoid library improved the chances of identifying transcripts not captured in the standard libraries, some of which could be associated with embryogenesis. This collection of ESTs is particularly well suited for use as candidate genes for development of an oil palm DNA chip, which can be used to obtain a more comprehensive view of the molecular mechanism associated with oil palm tissue culture. Keywords: auxin, expressed sequence tags, gene expression, low abundance genes, tissue culture.
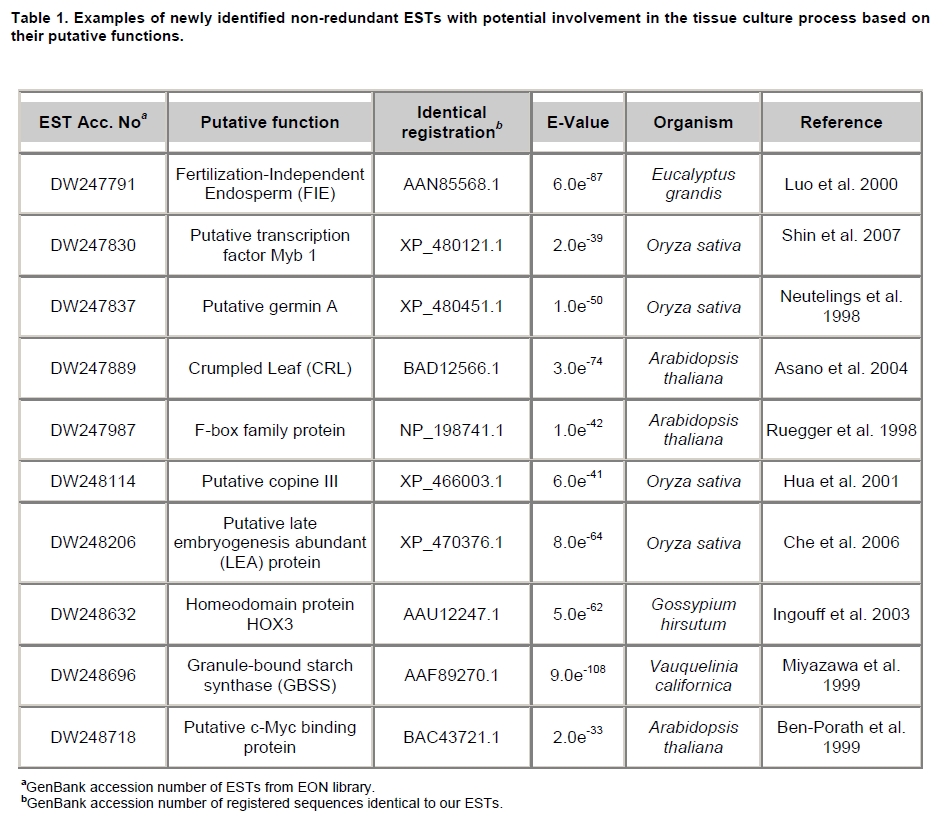
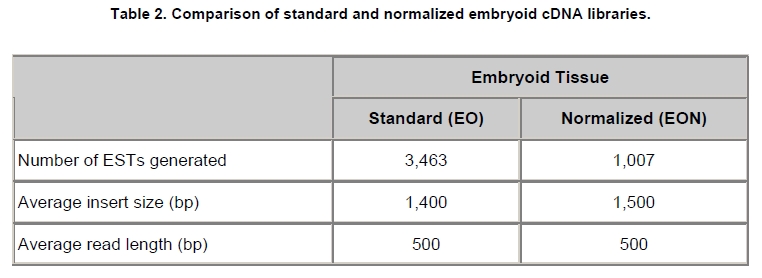
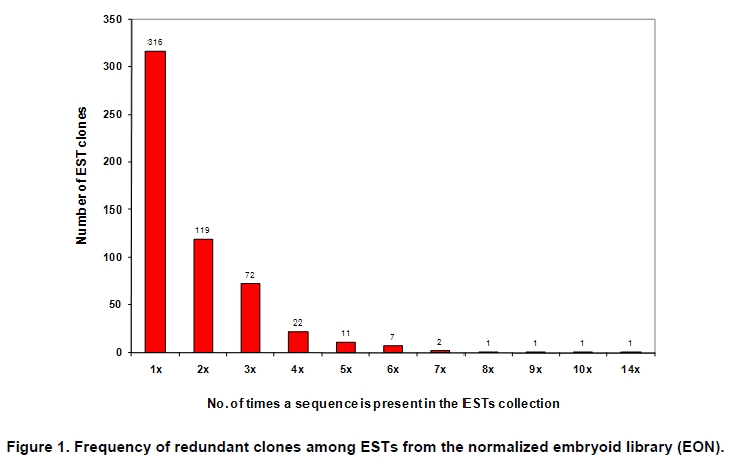
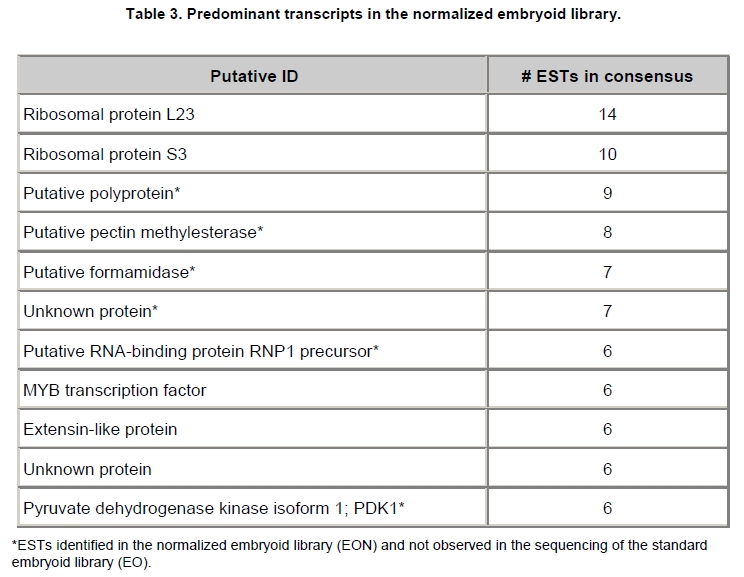
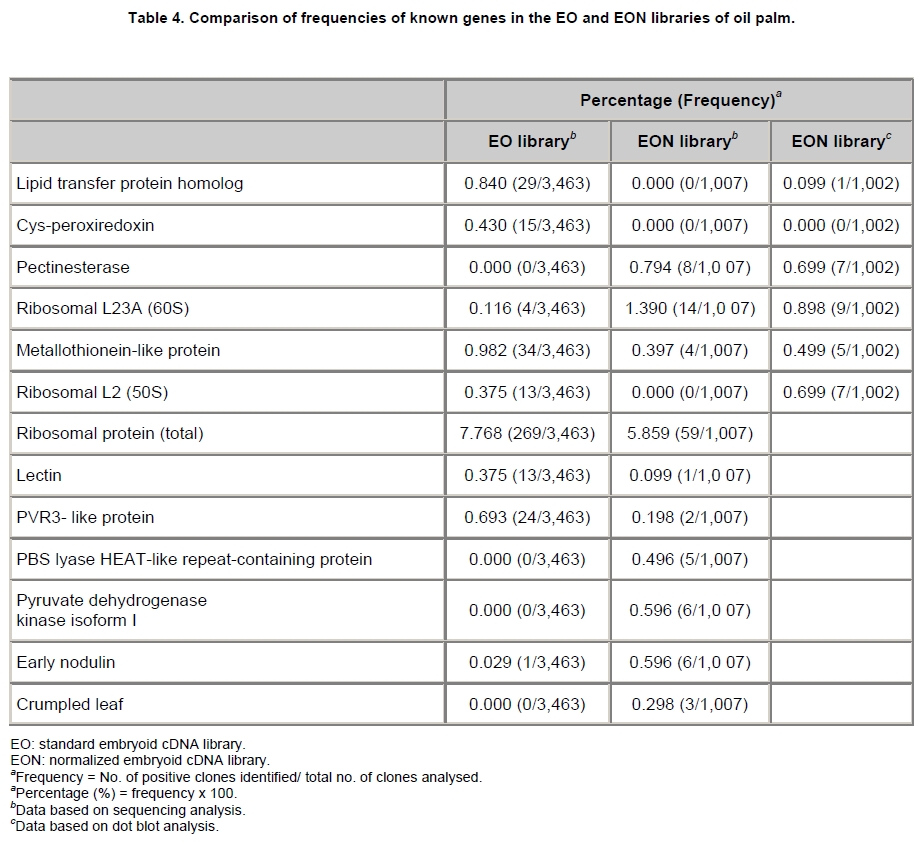
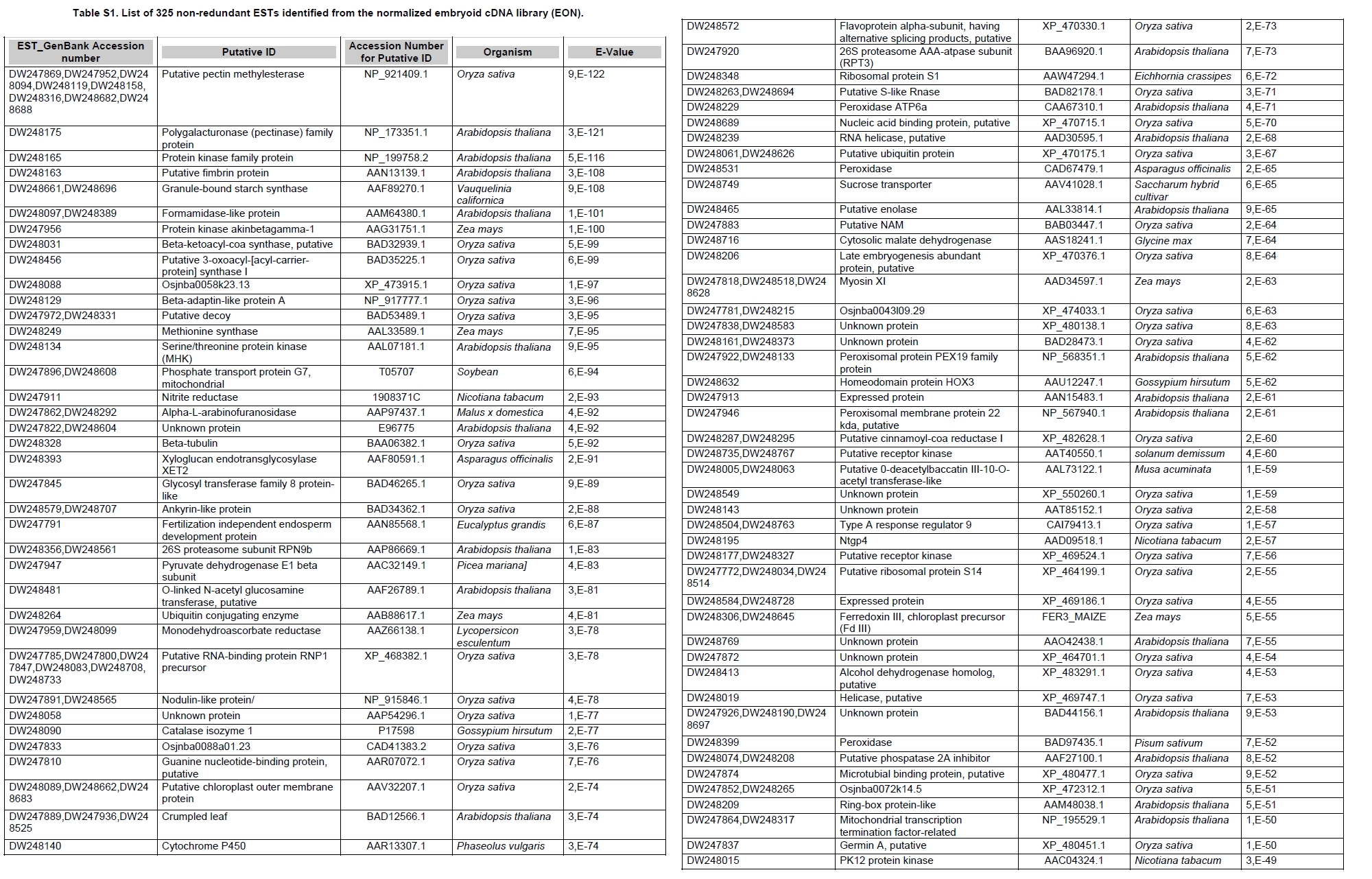
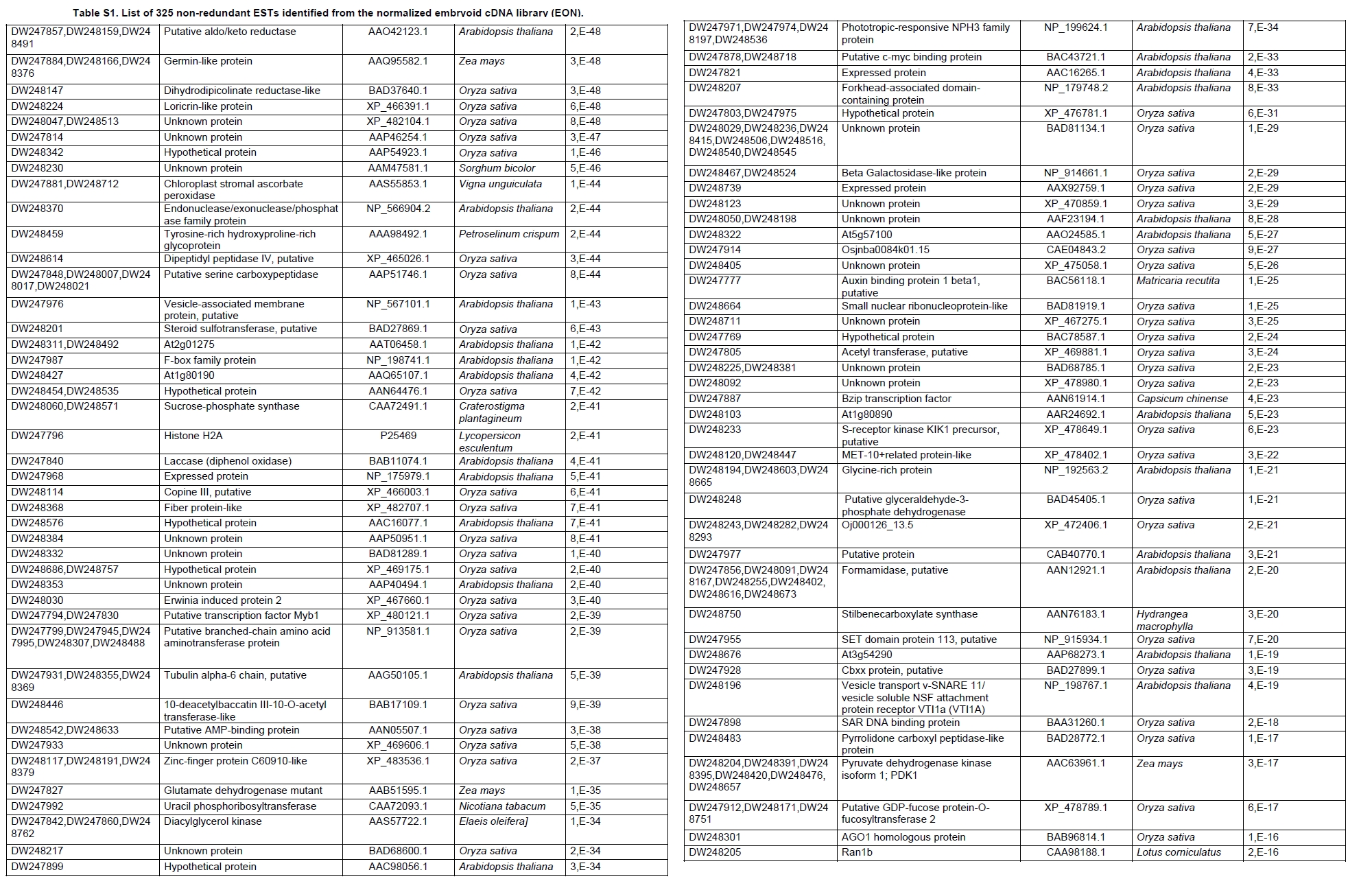
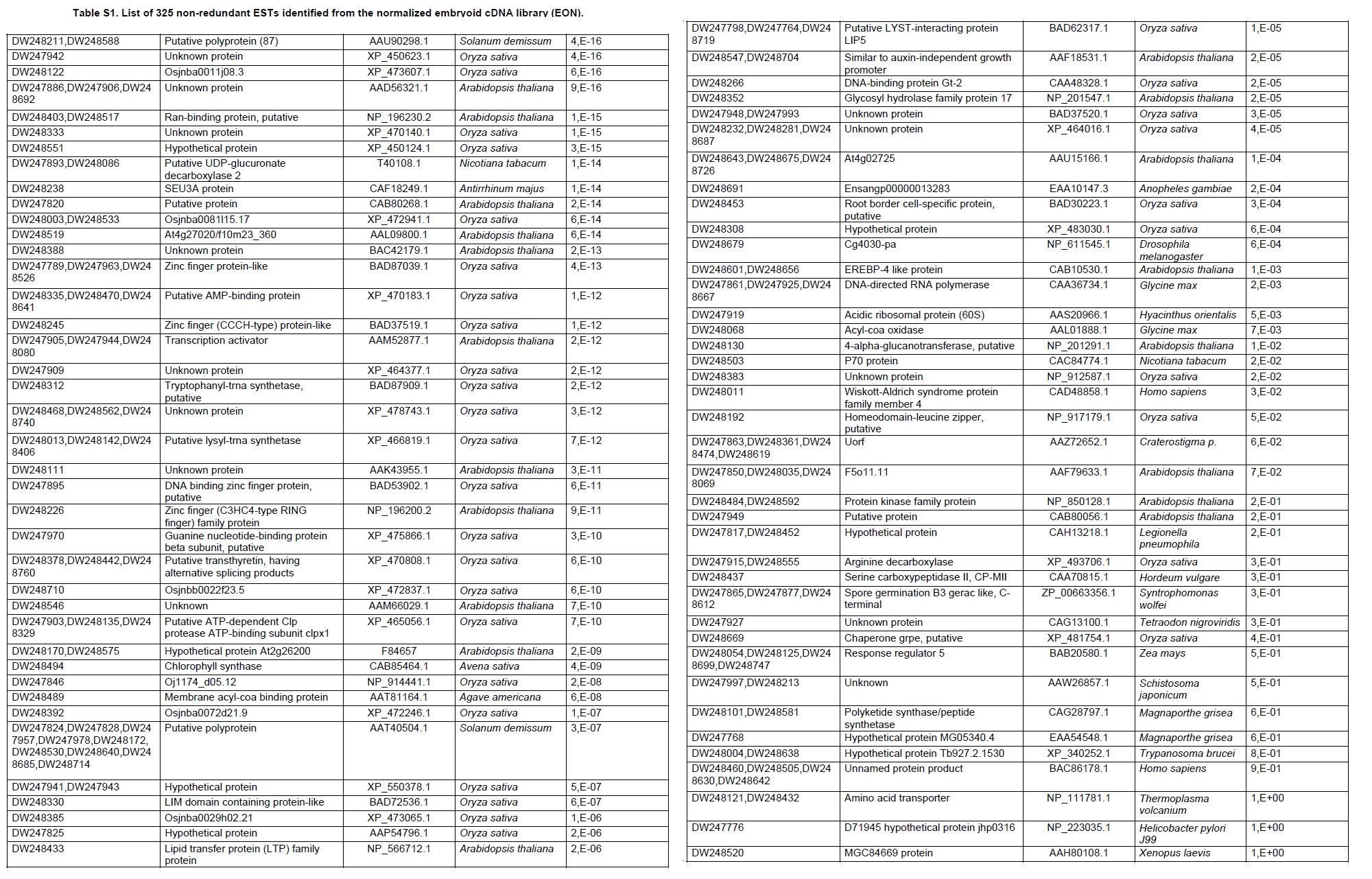
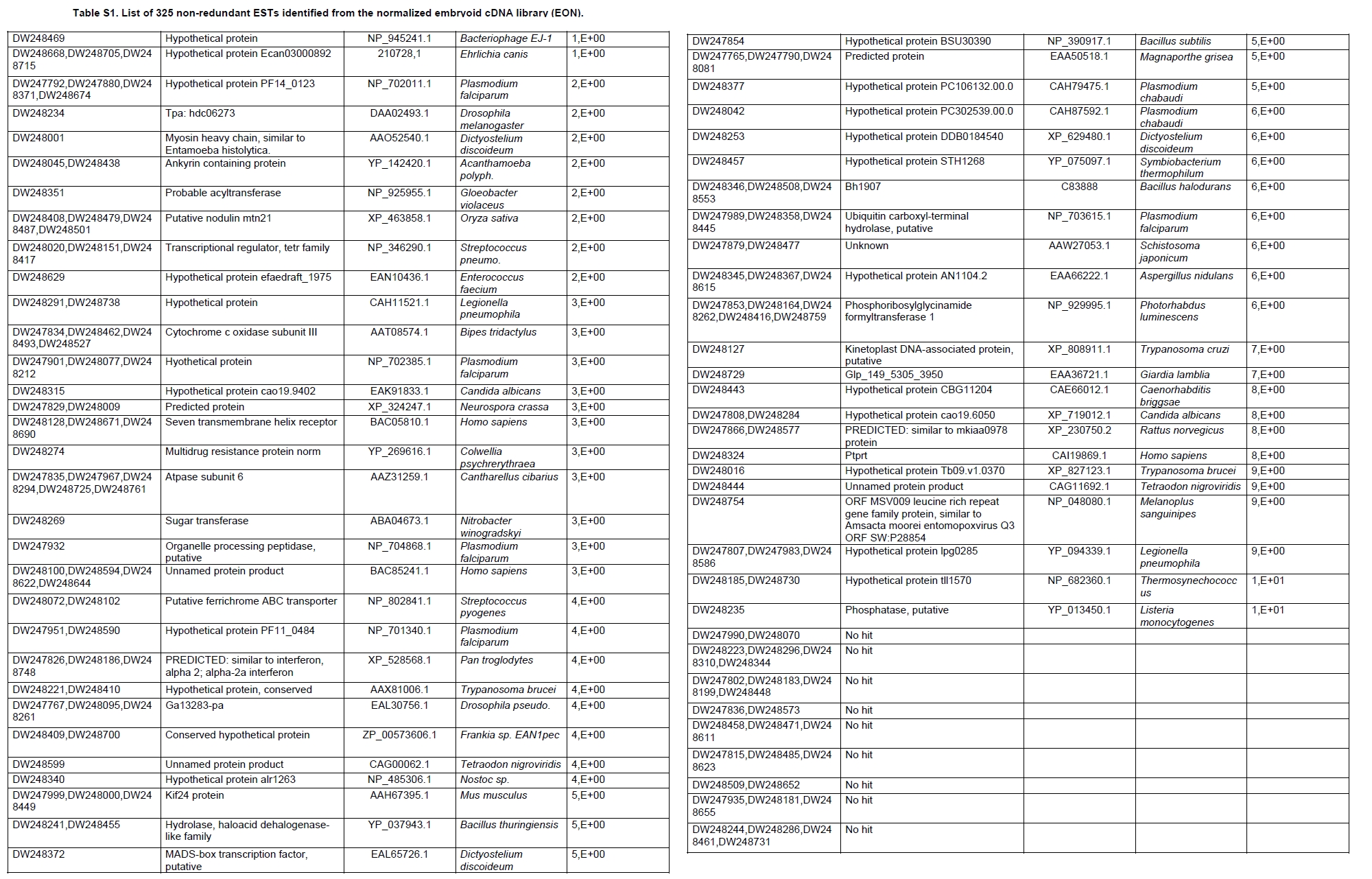
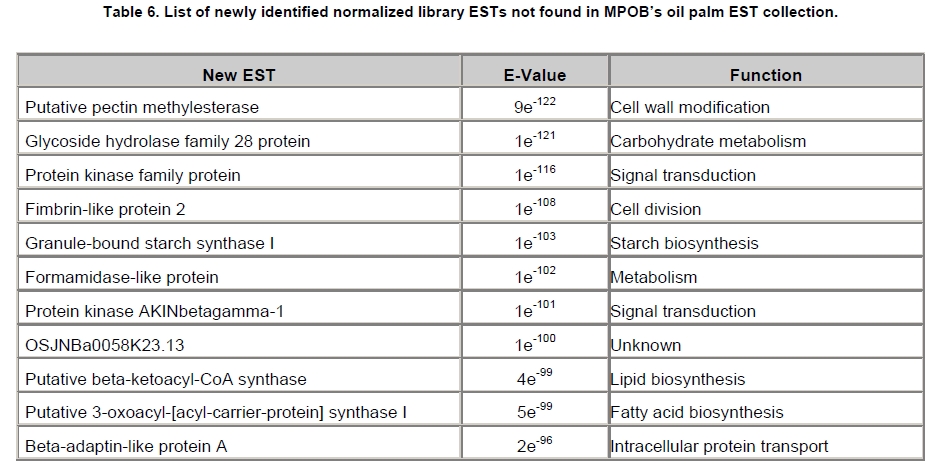
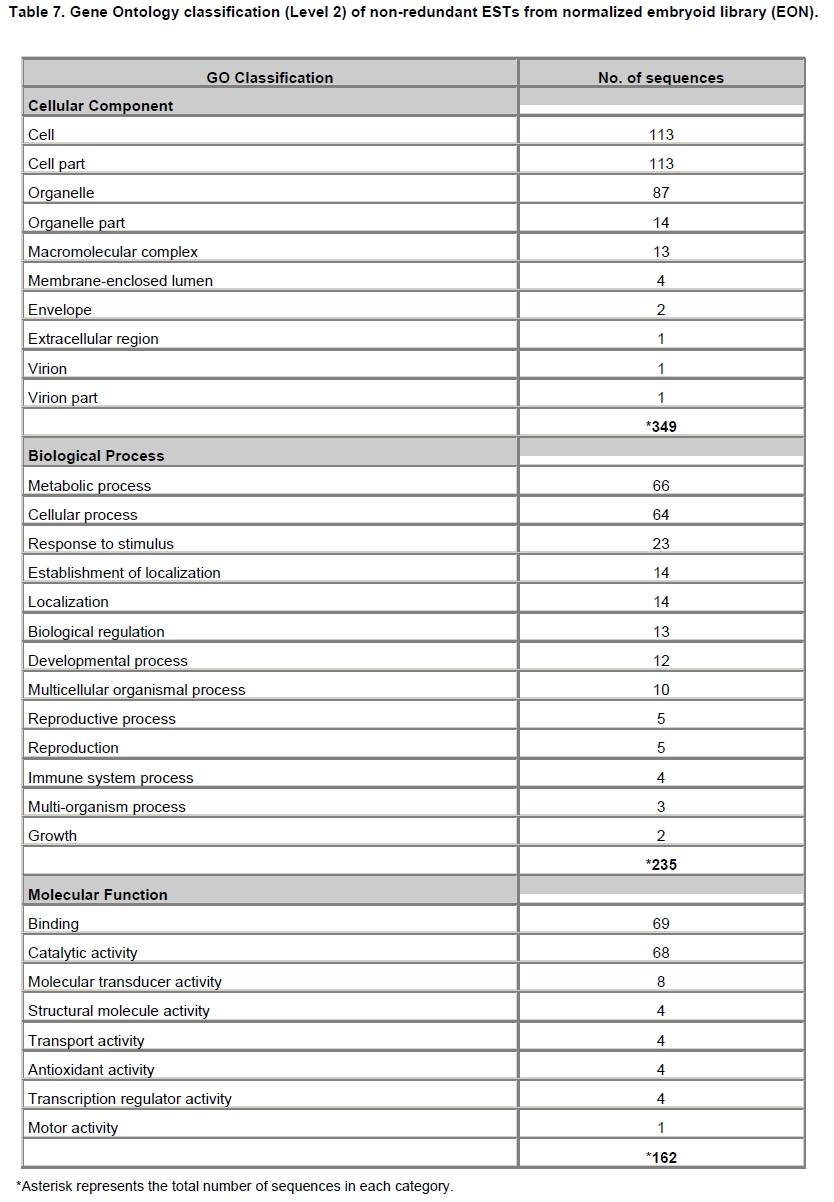
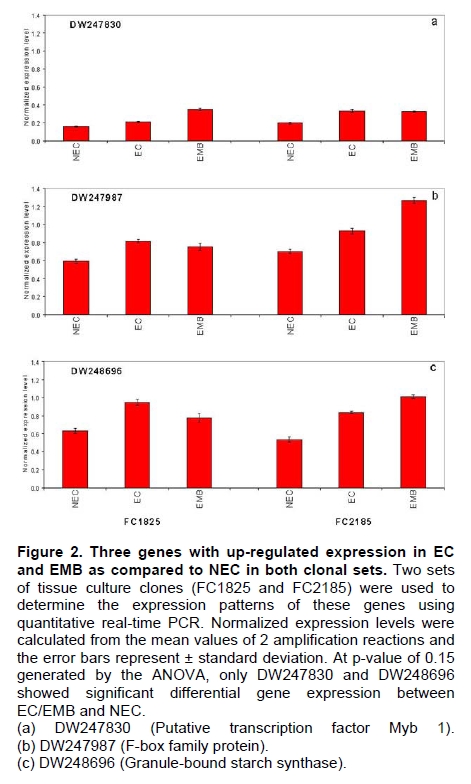
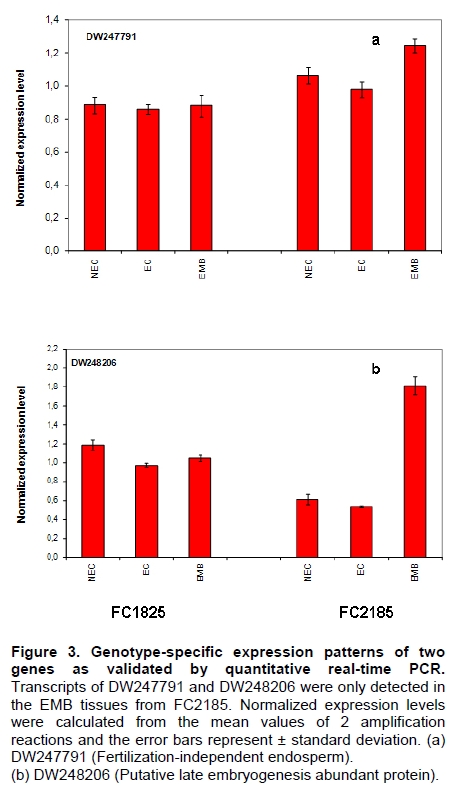
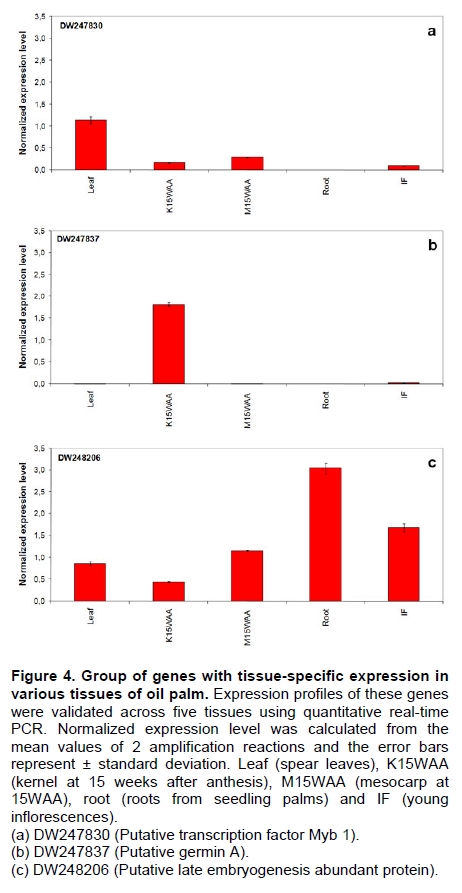
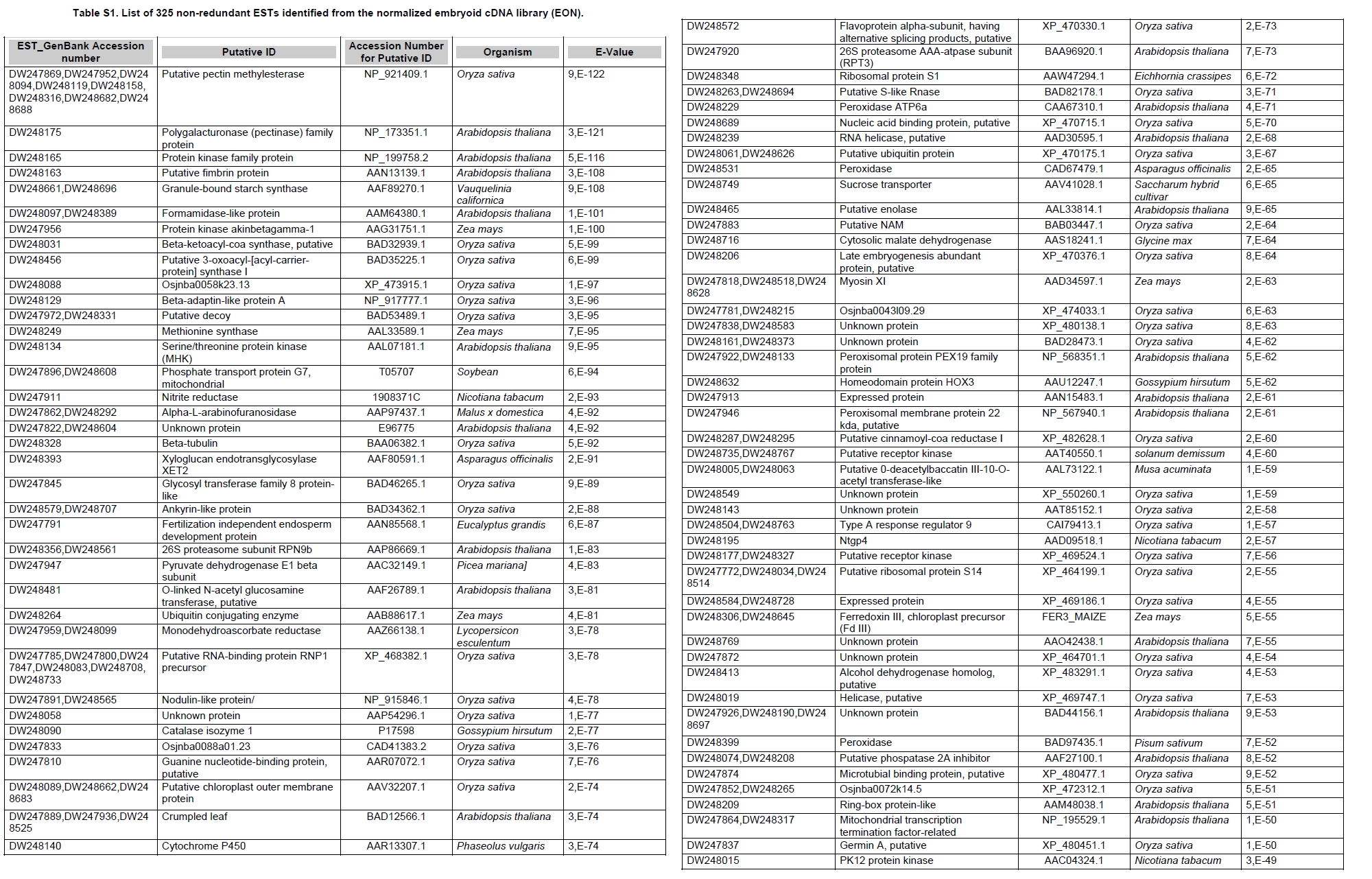
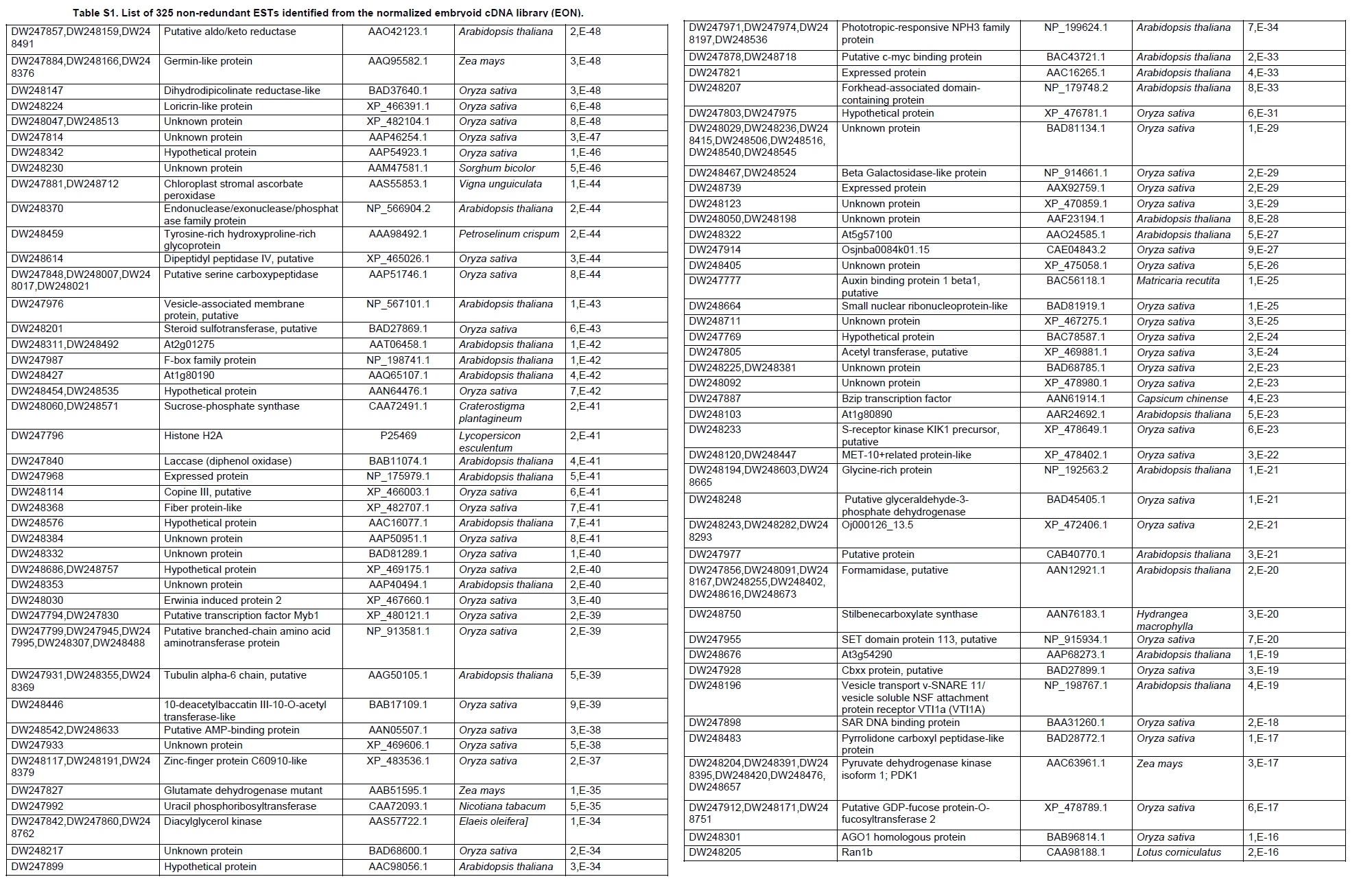
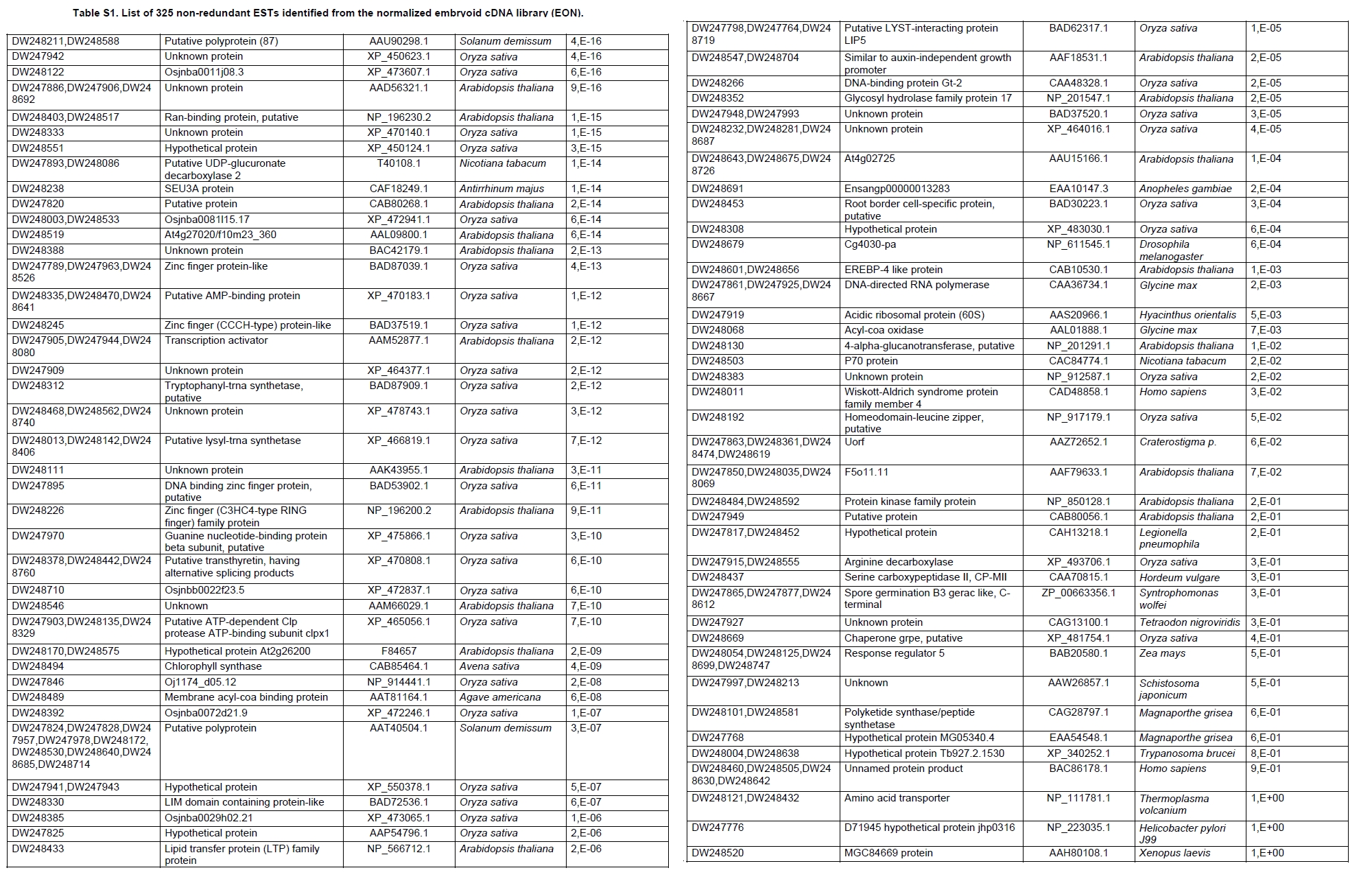
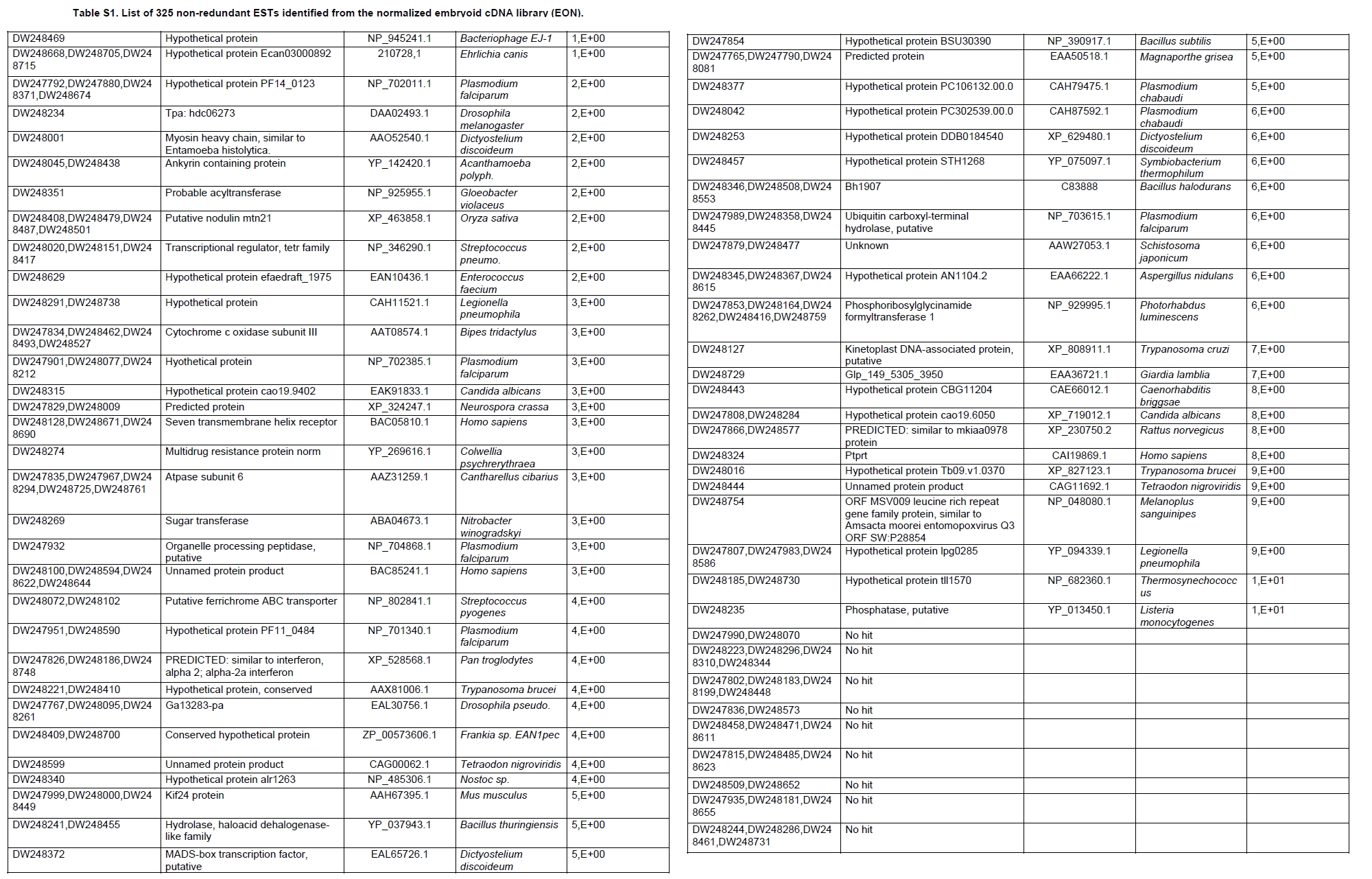
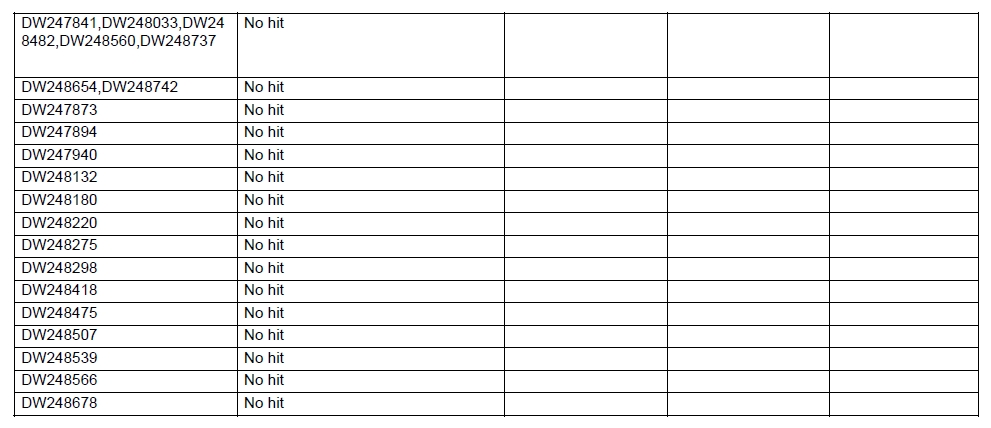
Oil palm is the most important source of vegetable oil globally. Its propagation by tissue culture was first described in the 1970s (Jones, 1974). Since then, the commercial advantage of tissue culture planting materials over conventional seedlings has been well established. However, the tissue culture process remains fraught with difficulties. The rate of callogenesis of oil palm explants remains low, at about 19% (Corley and Tinker, 2003), while the rate of embryogenesis from proliferating callus cultures is only 6 percent (Wooi, 1995). Attempts to overcome the problem via culture media and environmental modifications (Jones, 1974; Wong et al. 1996; Muniran et al. 2008), development of suspension culture (Tarmizi et al. 2004) and bioreactor system (Gorret et al. 2004) have somewhat improved the efficiency rate of oil palm tissue culture. However, low embryogenesis rate remains a recalcitrant problem. Therefore, understanding the molecular basis of oil palm tissue culture could provide the necessary information needed to improve the efficiency of the process further. The first step towards this would be to isolate and evaluate the genes that are expressed during oil palm tissue culture. The number of genes expressed in plants has been estimated at between 26,500 and 45,000 (Sterck et al. 2007). Typically, messenger RNAs are divided into three frequency classes: super-prevalent (10-15 mRNA species, each with about 5,000 copies, representing 10-20% of the total mRNA mass); intermediate (1,000-2,000 mRNA species, 225 copies per species, 40-45% of the total mass) and complex (15,000-20,000 mRNA species, 15 copies per species, 40-45% of the total mass) (Bishop et al. 1974; Zhang et al. 2005). However, the distribution of the expressed genes among the different tissues is highly variable. The massive presence of super-prevalent mRNAs in a tissue often hampers large-scale expressed sequence tag (EST) sequencing (Chu et al. 2003). As EST sequencing continues, the remaining unidentified genes become progressively harder to find because they are of lower abundance and restricted to only certain cell types. The most scarce and tissue-specific genes need to be catalogued as these genes play an essential role in plant development (Reddy et al. 2002; Shary and Guha-Mukherjee, 2004). Sequencing from standard cDNA libraries is ineffective for discovering rarely expressed genes, as the intermediate and highly expressed genes are sequenced more frequently. Therefore, a sequencing programme for expressed genes needs to be strategised in order to isolate all the classes of genes. This can be achieved by constructing a normalized cDNA library containing equal representations of genes expressed in a particular cell, tissue or organ. At least two main approaches for constructing normalized cDNA libraries have been reported (Weissman, 1987; Bonaldo et al. 1996). Weissman (1987) proposed to normalize a cDNA library through saturation hybridization of cDNAs with genomic DNA. This technique depends on the presence of relatively even copies of most genes in a genome. Although normalization can be achieved by this approach, it has been suggested that obtaining sufficient rare cDNAs to achieve saturation in the hybridization process is difficult (Soares et al. 1994). The rare cDNAs may also not hybridize to genomic DNA efficiently based on their reassociation kinetics. The other approach depends on reannealing of cDNAs following the principle of second-order kinetics. It is postulated that rare species anneal less rapidly and that the single-stranded fraction of the cDNA becomes progressively more normalized during the course of the hybridization (Soares et al. 1994; Kopczynski et al. 1998). For example, when 50% of the very rare species are annealed, the most abundant species in the single-stranded fraction would not be more than twice as abundant as the rarest species. Although this reassociation-kinetics based approach is considerably more complex, libraries can be readily produced in which the minor mRNA species from a small fraction of the cells of an organ are nearly as abundantly represented as the most abundant RNA species from the predominant cell population. Sequencing of tissue culture-related genes from oil palm standard cDNA libraries was described previously by Ho et al. (2007) and Low et al. (2008). A total of 1,723 ESTs were obtained from suspension cell culture (Ho et al. 2007), while another 17,599 ESTs were generated through the sequencing of standard cDNA libraries from leaf-derived embryogenic callus (EC), non-embryogenic callus (NEC) and embryoid (EMB) (Low et al. 2008). In order to capture rarely expressed genes associated with somatic embryogenesis, the existing EMB library was chosen for normalization following the method described by Bonaldo et al. (1996). The method is simple and has certain advantages over other procedures. It is based on the reassociation-kinetics principle involving hybridization of a 10-fold excess of driver over tracer. The DNA driver is generated by PCR amplification of the tracer (single-stranded DNA of the starting cDNA library) produced in vivo using the helper phage M13K07. The driver hybridizes with the tracer in the form of single-stranded circles, followed by hydroxyapatite (HAP) column purification of the remaining unbound single-stranded plasmids. The single-stranded plasmids are converted to double-strands and electroporated into bacterial host cells. This technique preserves the longest cDNAs and minimizes redundancies while increasing the representation of rare cDNAs by about three-fold on average (Bonaldo et al. 1996). The discovery of rarely expressed genes from the normalized embryoid cDNA library (EON) will enrich the existing EST collection of genes expressed during oil palm tissue culture. Availability of these non-redundant ESTs by the normalization process will aid in the development of a comprehensive cDNA microarray for identification of candidate genes that may be regulated during oil palm tissue culture. Materials and Methods Different stages of oil palm (Elaeis guineensis Jacq.) tissue culture materials (NEC, EC, EMB) were provided by FELDA Agricultural Services Sdn. Bhd. Various tissues of oil palm (spear leaves, kernel at 15 weeks after anthesis (WAA), mesocarp at 15WAA, roots from seedling palms and young inflorescences) were harvested from tenera palms at Malaysian Palm Oil Board (MPOB)-UKM Research Station, Bangi, Selangor, Malaysia. All the samples were frozen at -80ºC prior to RNA extraction. Construction of standard embryoid cDNA library (EO library) Total RNA was isolated from oil palm embryoid cultures as described by Rochester et al. (1986). Poly (A)+ RNA was purified using oligo-dT cellulose chromatography according to Singh and Cheah (2000). The cDNA library was constructed using the ZAP-cDNA® Gigapack® III Gold Cloning kit (Stratagene). Mass excision of cDNA clones from an amplified library was performed using the ExAssist Helper phage following the manufacturer's instructions (Stratagene). The excised phagemids were transformed into SOLR cells and grown at 37ºC overnight on LB agar plates containing 100 µg/ml ampicillin. Double-stranded DNA (dsDNA) from the SOLR cells was extracted and transformed into XL1-Blue MRF' competent cells (Hanahan, 1985). Preparation of single-stranded DNA (ssDNA) Conversion of dsDNA into ssDNA was carried out by super infection of XL1-Blue MRF' transformants with the helper phage M13K07 (Vieira and Messing, 1987). Transformants (white colonies) were pooled and grown at 37ºC overnight in LB broth containing 100 µg/ml ampicillin. The overnight bacterial culture was diluted to 1:100 and allowed to grow further to an OD600 of 0.2. The helper phage M13K07 was added to a multiplicity of infection (M.O.I) of 10- to 20-fold excess and incubated for 1 hr at 37ºC with gentle shaking (200 rpm). A final concentration of 70 µg/ml kanamycin was then added to select the infected bacterial cells, and the growth was continued overnight with vigorous shaking (300 rpm). After centrifugation (4,000 rpm, 10 min, Sorvall RC-5C PLUS), 20% PEG/2.5 M NaCl solution was added to the supernatant for precipitation of phage particles at 4ºC overnight. Phage pellet obtained from centrifugation (12,000 rpm, 20 min, Sorvall RC-5C PLUS) was dissolved in TE buffer (pH 8.0) and extracted with phenol: chloroform: isoamyl alcohol (25:24:1) for protein removal. The ssDNA phagemids were ethanol-precipitated (at least 1 hr, -20ºC) and the pellet was dissolved in TE buffer (pH 8.0). Digestion of the ssDNA with PvuII enzyme was performed at 37ºC for 4 hrs prior to purification using a Bio-Gel HTP HAP column (Bio-Rad) as described previously (Soares et al. 1994; Ali et al. 2000). PvuII digestion was used to eliminate double-stranded plasmid contaminants in the ssDNA preparation. The eluted ssDNA from HAP column was then concentrated by extracting twice with water-saturated butanol, once with dry-butanol and once with water-saturated ether. The sample was desalted by passing through a spin column from QIAquick PCR Purification Kit (Qiagen), followed by purification using a Microcon YM-100 column (Millipore Inc.). The driver DNA was prepared by PCR amplification of ssDNA using the SK primer (5’- CGCTCTAGAACTAGTGGATC -3’) and R primer (5’- ATAGGGCGAATTGGGTAC -3’). 10 ng of ssDNA was amplified for 35 cycles using a step cycle program (95ºC for 30 sec, 63ºC for 30 sec and 72ºC for 2 min) in 100 µl reaction mixture containing 1 µM of both primers, 200 µM dNTPs, 1.5 mM MgCl2, 1 x PCR buffer (Invitrogen) and 1 U Taq DNA polymerase (Invitrogen). The amplified PCR products (driver DNA) were purified using the QIAquick PCR Purification Kit (Qiagen). Normalization of standard embryoid cDNA library Reassociation method described by Bonaldo et al. (1996) was used to normalize the EO library. The reaction mixture containing driver (10 µg), tracer (1 µg of single-stranded plasmid DNA), 50% deionized formamide, 200 µg each of 5'-blocking oligonucleotide (5’-CGCTCTAGAACTAGTGGATCCCCCGGGCTGCAGGAATTCGGCACGAGG-3’) and 3’-blocking oligonucleotide (5’-A18 CTCGAGGGGGGGCCCGGTACCCAATTCGCCCTAT-3’) was heated to 80ºC for 3 min under mineral oil. This was followed by addition of 20 µl of 10 x hybridization buffer (1.2 M NaCl, 0.1 M Tris pH 8.0 and 50 mM EDTA) and 20 µl of 10% SDS. Hybridization was then carried out at 35ºC for 20 hrs (Calculated C0t ~5; C0t value = initial DNA concentration in moles nucleotides per liter X the reassociation time in second). The remaining ssDNA was purified by HAP chromatography and converted into dsDNA using Sequenase Version 2.0 (USB Corp.) prior to electroporation into competent cells of E. coli DH5α'. The entire normalized cDNA library was plated onto LB agar containing 100 µg/ml ampicillin. Sequencing and analysis of ESTs Randomly selected clones from the normalized library were sequenced from the 5’ end on an ABI PRISM 377 (Applied Biosystems). Sequence analysis was carried out using the PHRED (Ewing and Green, 1998; Ewing et al. 1998) and StackPACK programs (Miller et al. 1999). BLAST search was carried out against the GenBank non-redundant protein database. Functional classification of the non-redundant ESTs was performed using Blast2GO (Conesa et al. 2005). A different batch of cDNA clones (1,002) was randomly selected from the EON library for dot blot hybridization. For this purpose, 200 ng of plasmid from each of the 1,002 cDNA clones were transferred onto Hybond-N+ nylon membranes (GE Healthcare, formerly Amersham Biosciences). The radioactive probes used for screening were gel purified cDNA fragments of pectinesterase, ribosomal L23A (60S), ribosomal L2 (50S), cys-peroxiredoxin, lipid transfer protein homolog and metallothionein-like protein. The membranes were pre-hybridized in 5 x SSPE, 5 x Denhardt’s (1 x Denhardt’s solution is 0.02% each Ficoll 400, bovine serum albumin and polyvinylpyrrolidone), 0.5% SDS and 100 µg/ml herring sperm DNA at 65ºC for 4 hrs. Hybridization was carried out in a buffer containing 5 x SSPE, 0.5% SDS, 100 µg/ml herring sperm DNA and 32P-labelled probe (1-2 x 106 cpm/ml) at 65ºC overnight. The membranes were washed at 65ºC in 2 x SSC/0.1% SDS (30 min), 1 x SSC/0.1% SDS (30 min) and 0.5 x SSC/0.1% SDS (30 min). Positive clones were manually counted after exposure of the membranes to X-ray film (Kodak) for 4-8 hrs. Total RNA was isolated from tissue culture materials and various tissues of oil palm according to McCarty (1986). The total RNA was purified using RNeasy Mini Kit (Qiagen), and on-column digestion with RNase-free DNase I (Qiagen) was performed in the RNeasy Mini Spin column according to the manufacturer’s instructions. The quality of the total RNA was investigated by electrophoretic fractionation on an Agilent 2100 Bioanalyzer (Agilent Technologies) and a RNA 6000 Nano LabChip® (Caliper Technologies Corp.). Reverse transcription of total RNA to first-strand cDNA was carried out using High-capacity cDNA Reverse Transcription Kits (Applied Biosystems). A total of 2 µg of total RNA was reverse transcribed in the presence of 50 units of MultiScribeTM Reverse Transcriptase in a total volume of 20 µl. This step was performed under the following temperature scheme: 10 min at 25ºC, 120 min at 37ºC, 5 sec at 85ºC. The single-stranded cDNA was used as template for quantitative real-time PCR using ABI PRISM® 7000 Sequence Detection System (Applied Biosystems) equipped with ABI Prism 7000 SDS Software (Applied Biosystems). Specific primers and TaqMan® probes for all the genes listed in Table 1 were designed using the Primer Express Software (Applied Biosystems). For each PCR reaction, 40 or 80 ng of cDNA was added to 12.5 µl TaqMan® Universal PCR Master Mix (Applied Biosystems) and 1.25 µl 20 x TaqMan® Gene Expression Assays (Applied Biosystems) in a total volume of 25 µl. PCR was carried out at 50ºC for 2 min, 95ºC for 10 min, followed by 40 cycles at 95ºC for 15 sec and 60ºC for 1 min. Following amplification, threshold cycle (Ct) values were determined by adjusting the threshold setting within the exponential amplification region. The Ct values were accepted if the standard deviation between two replications was smaller than 0.5. Delta-Ct method was then used to transform the Ct values to raw expression values. Three housekeeping genes with the most stable expression in oil palm tissues were used in normalization of raw expression values of the genes of interest (GOI). Geometric mean of the three housekeeping genes in each tissue, which is known as normalization factor, was calculated according to Vandesompele et al. (2002). Normalized expression levels of GOI in each tissue were obtained by dividing the raw expression values of GOI in each tissue with the appropriate normalization factor. The three housekeeping genes encode for superoxide dismutase, DNA polymerase III and unknown protein (unpublished data). Analysis of variance (ANOVA) was performed across normalized expression values obtained from the two different sets of tissue culture clones. This was carried out for the ESTs that were used to screen for the embryogenic potential of callus. Values of p < 0.15 were considered as significant differential gene expression between NEC and EC/EMB. The statistical analysis was performed using TIGR Multiexperiment Viewer version 4.1 (Saeed et al. 2003). Results The standard embryoid cDNA library (EO) and its normalized library (EON) The EO library used for normalization was described by Low et al. (2008). The EO library had a titer of 109 pfu/ml and the average insert size of 1,400 bp as determined by PCR amplification of 3,463 randomly selected clones from the EO library using vector-specific primers. Sequences from this library have been deposited in the dbEST division of GenBank (Accession numbers: EY408451-EY413718). A normalized library was constructed from the standard cDNA library in order to reduce the frequency of high abundance cDNAs and increase the chances of identifying the low abundance genes. The normalization involved hybridization of a large excess of cDNA inserts with the single-stranded cDNA of the EO library until a C0t value of 5 was reached. The conditions used allowed the high and moderately abundance cDNAs to anneal to form dsDNA. The remaining ssDNA was purified by HAP chromatography, yielding the normalized EON library with at least 10,000 colonies. PCR amplification of 1,007 randomly selected cDNA clones from this library revealed that the average insert size is 1,500 bp (Table 2). Cluster analysis of ESTs from the normalized library The cDNA clones from the EON library were randomly selected and sequenced from the 5' end. Similarity search of 1,007 high quality ESTs with at least 400 bp insert size was performed against the GenBank protein database using BLASTX. With an E-value cut off at 10-6, approximately 68% of the ESTs showed significant similarity to known sequences from plants. The other 32% of the ESTs had no matches with any known sequences. These sequences are either unique to oil palm (Tu et al. 2007) or could represent sequences containing non-coding regions such as the 5’ untranslated region (5’ UTR) which are less conserved across species (Jouannic et al. 2005). The 1,007 nucleotide sequences reported have been deposited in GenBank (Accession numbers: DW247764-DW248770). StackPACK clustering analysis of the 1,007 ESTs resulted in the identification of 316 singletons (groups that contain only one EST) and 237 clusters, which were assembled from two or more ESTs in the EON library (Figure 1). A total of 553 non-redundant ESTs (singletons + a representative of each consensus) were thus identified. Of these 553 ESTs, 399 showed significantly similarity to known sequences in the non-redundant protein database at a cut off E-value of 10-6 (397 exhibited significant similarity to plant sequences). The remaining 154 (28%) did not show significant similarity to any known sequences in the public databases. Some of these ESTs may be specific to oil palm. The list of EST transcripts predominant in the normalized library is shown in Table 3. Ribosomal protein L23A with a cluster of 14 sequences was predominant. The other abundant transcripts were ribosomal protein S3, putative polyprotein, putative pectin methylesterase, putative formamidase, extensin-like protein, etc. Among these transcripts, some were not identified in the standard EO library. This group of genes is also indicated in Table 3. Evaluation of normalization by sequencing analysis Frequency of redundant clones in EO and EON library were examined to verify the degree of normalization. The results showed that normalization had reduced the frequency of highly expressed genes, such as ribosomal protein, metallothionein-like protein, lipid transfer protein homolog, cys-peroxiredoxin and PVR3-like protein. Conversely, it increased the frequency of low abundance genes like pectinesterase, early nodulin, PBS lyase HEAT-like repeat-containing protein and crumpled leaf. Table 4 summarizes the frequency of the cDNA clones observed in the sequencing of the two libraries (EO and EON). The results suggest that normalization process minimized the differential abundance of the various cDNA clones in the EO library. Evaluation of normalization by dot blot hybridization A total of 1,002 cDNA clones from the EON library were screened by dot blot hybridization. These clones were different from those used in the sequencing analysis. To evaluate the extent of the normalization process, six probes representing low, medium and high abundance genes in the EO library were chosen for the hybridization experiments (Table 4). Positive signals generated from hybridization between the selected probes and cDNA clones were recorded. Comparing the dot blot data of the EON library with the sequencing data of the EO library revealed that normalization reduced the frequency of highly expressed genes and simultaneously increased the frequency of the genes not captured in conventional cDNA libraries. The comparison is summarized in Table 4. As expected, the frequencies of highly abundant clones such as lipid transfer protein homolog and metallothionein-like protein were reduced 8-fold and 2-fold, respectively in the EON library. For the medium abundance clones, the frequency of cys-peroxiredoxin was significantly decreased while a 2-fold increase in ribosomal L2 (RPL2) was observed. Promising results were also obtained for the lowly abundant clones, pectinesterase and ribosomal L23A (60S). The frequency of these genes increased significantly in the EON library as compared to the EO library. New ESTs identified by normalization Normalization increased the frequency of identifying new ESTs and this in return increased the size of the EST collection. Clustering of the ESTs from the EO and EON libraries revealed that 76% (762 out of 1,007; 423 non-redundant ESTs) of the ESTs in the EON library were not previously sequenced. However, when the clustering was performed with all the EST collection at MPOB (17,599 ESTs, GenBank accession numbers: EY396120 to EY413718), the percentage of ESTs specific to the EON library was reduced to approximately 57% (570 out of 1,007). From these numbers, 325 non-redundant ESTs (188 singletons and 137 consensus sequences) were obtained (Electronic supplementary materials, Table S1a, b, c, d, e) with 205 exhibiting significant homology to known sequences in the public databases. When the 325 non-redundant ESTs were further compared to the published sets of oil palm ESTs by Jouannic et al. (2005) and Ho et al. (2007), 237 ESTs were found unique to the EON library (Table 5). This revealed that the normalization process greatly increased the discovery of new ESTs. Examples of the ESTs identified from the normalization process together with their biological functions are given in Table 6. Functional classification of ESTs from the EON library Blast2GO was used to assign 325 non-redundant ESTs from the EON library to three main functional categories: Cellular Component, Biological Process and Molecular Function. It was found that only 152 ESTs (47%) were successfully mapped to the Gene Ontology Consortium structure under one or more ontologies. The summary of Gene Ontology classification at Level 2 is shown in Table 7. The ESTs were mostly assigned to the Cellular Component (349) category, with 65% involved in functions related to cell and cell part. The second highest category was assigned to the Biological Process, where about 28% were mapped to metabolic process, followed by cellular process (27%) and response to stimulus (10%). With respect to Molecular Function, only 162 sequences were classified under this category. The predominant functional classes in this category are assigned under binding (43%) and catalytic activity (42%). Expression studies of genes associated with oil palm tissue culture using quantitative real-time PCR Many of the unique ESTs identified encoded for genes which, based on their putative functions, may be involved in the tissue culture process. Examples of these genes are shown in Table 1. This group of genes may provide further insight into the molecular mechanism associated with oil palm tissue culture. With this in mind, quantitative real-time PCR was used to profile the expression of these genes across two sets of tissue culture clones (FC1825 and FC2185, each comprising the three main developmental stages in oil palm tissue culture: NEC, EC, EMB). The main purpose of this study was to determine if the genes are differentially expressed in NEC, EC or EMB. To ensure that the transcript levels observed were not due to differences in their genetic backgrounds, each set (NEC, EC and EMB) was sampled from a single genotype. The use of two genotypes (FC1825 and FC2185) can help determine if there is potential for the differences in transcript level to be reproducible across different genetic backgrounds. The inclusion of tissues such as spear leaves, kernel at 15 WAA, mesocarp at 15WAA, roots from seedling palms and young inflorescences (with unopened flowers) was also to determine expression of the selected genes in the various tissues of whole plant. Based on the analysed real-time PCR data, only five of the ESTs demonstrated informative expression profiles in the tissue culture samples. The remaining five gene clones (data not shown) were expressed through out the different tissue culture developmental stages, however no significant expression profiles were observed. The three ESTs in Figure 2-DW247830 (putative transcription factor Myb1), DW247987 (F-box family protein) and DW248696 (granule-bound starch synthase)- were up-regulated in EC and EMB as compared to NEC. However, at p-value of 0.15 generated by the ANOVA, only DW247830 and DW248696 showed significant differential gene expression between EC/EMB and NEC. The expression patterns of these two transcripts were reproducible in both sets of the tissue culture clones, where they were consistently down-regulated in NEC. Up-regulation of these genes is an indicator that the callus has potential to produce embryoids. Another group of two ESTs (Figure 3) -DW247791 (fertilization-independent endosperm) and DW248206 (putative late embryogenesis abundant protein)- exhibited higher expression in EMB of the genotype FC2185. The expression patterns of these transcripts are considered genotype-specific. It may be interesting to further explore their expression profiles across a wider range of tissue culture materials. To determine the expression of these genes in the whole plant, the 10 selected TaqMan® probes were also screened across various tissues of oil palm. The results are presented in Figure 4. Interestingly, DW247837 (putative germin A) was exclusively expressed at high levels in the kernel at 15WAA. A few other genes also showed tissue-specific expression with at least 0.8-fold up-regulation in specific tissues compared to other tissues tested. Clones DW247830 and DW248206 were abundantly expressed in the spear leaf and root tissues, respectively. The genes associated with tissue culture also appear to play a vital role in the development and maintenance of highly differentiated plant organs. Discussion Experimental strategy of the normalized process The main objective of this study was to construct a normalized library with approximately equal representation of all the mRNA sequences in order to increase the chances of identifying rare genes in embryoid tissues. The normalized cDNA library denoted EON was constructed based on the reassociation kinetics reaction. Effectiveness of the normalization process was confirmed through direct sequencing and dot blot analysis. Direct sequencing of 1,007 randomly picked ESTs demonstrated changes in the redundancy level of certain genes in EON library compared to the standard EO library. Frequency of genes, such as ribosomal protein, metallothionein-like protein, lipid transfer protein homolog and cys-peroxiredoxin, which were reported to be highly abundant in the standard EO library were reduced in the EON library. The frequency of low abundance genes like pectinesterase, early nodulin, PBS lyase HEAT-like repeat-containing protein and crumpled leaf were increased. The same pattern was also observed in the dot blot hybridization experiment using a different set of EST clones (1,002). It was found that the number of lowly abundant clones such as pectinesterase and ribosomal L23A (60S) were significantly increased in the EON library. The dot blot hybridization method demonstrated here has not been used as a screening method in previous studies on normalized libraries. The method is time-consuming, since plasmids of each cDNA clone have to be prepared for spotting on the membranes. Nevertheless, it is a simple and effective method to complement the sequencing results in validating the normalization process. Differences in frequency of ESTs between ribosomal protein L2 and L23A were observed in EON library. While ribosomal protein L2 and total ribosomal protein declined with normalization, frequency of ribosomal protein L23A increased. The results are unusual for ribosomal protein genes, as it is generally assumed that most of the ribosomal protein genes share a common promoter which leads to a unified control mechanism of gene expression. However, in the work carried out by Barakat et al. (2001) using Arabidopsis ESTs, it was found that the frequency of ESTs among different families of ribosomal protein genes varies greatly and the expression was also regulated differently. Furthermore, it was also observed that although RPL23A-1 and RPL23A-2 genes were 94% identical at the amino acid level, differences in the arrangement and number of predicted motifs at the 5’ regulatory regions had probably contributed to the differential transcriptional regulation of these genes (McIntosh and Bonham-Smith, 2005). The analysis of ribosomal protein genes in human also concluded that small numbers of ribosomal protein genes were regulated in a tissue-specific manner and each of them is controlled by different regulators (Ishii et al. 2006). These findings will shed light on the study of ribosomal protein genes across plant species. The efficiency of the normalization method was further supported by the clustering results of the ESTs. Comprehensive analysis revealed that 325 non-redundant ESTs from the EON library had not been previously identified in the EST collection at MPOB. About 47% of these ESTs were mapped to the GO terms, where majority of the genes appeared to be related to component of cells and organelles. This is expected as the cells at embryoid stage are actively dividing to form differentiated cells containing high storage lipid content and starch reserve (Kanchanapoom and Domyoas, 1999). When the comparison was extended to other published sets of oil palm ESTs (Jouannic et al. 2005; Ho et al. 2007), 237 ESTs were still specific to the EON library. The normalization process, as expected, increased the chances of identifying the low abundance transcripts, not isolated using conventional EST sequencing or any other routine molecular biology techniques. These genes are also listed in the Electronic supplementary material (Table S1a, b, c, d, e). Identification of potential somatic embryogenesis-related genes Regeneration of plants through the tissue culture process is important for mass propagation of elite planting materials (Sharry et al. 2006) and production of transgenic plants through genetic engineering (Masani and Parveez, 2008). In oil palm, the estimated time taken for the tissue culture process (from sampling of immature leaf explant to field planting) is around 58 months (Rohani et al. 2000). Major limitation of this process is the low embryogenesis rate. Therefore, identification of genes associated with somatic embryogenesis can help improve the current embryogenesis rate, which remains at 6%. Initially, researchers used the candidate gene approach to identify potential genes. This led to the association of auxin-inducible genes and stress response genes with the somatic embryogenesis process in oil palm (Meilina and Ooi, 2006). Another interesting gene, serine kinase, which is a classical embryogenesis marker, is also postulated to be involved in the signal transduction pathway of oil palm somatic embryogenesis (Ooi et al. 2008). This gene is one of the non-redundant EST (325) found in the EON library and was not observed in the previously sequenced standard cDNA libraries reported by Low et al. (2008). Studies by Ooi et al. (2008) also showed that the transcript levels of these kinases were low in embryogenic calli and suspension cultures. This could be the main reason why serine kinase was not captured in the standard library and not predominantly found in EON library. The advancement of technology has now made it possible to explore the expression profiles of large number of genes simultaneously, instead of relying on the candidate gene approach. Such technology, like microarray, has been implemented to profile the gene expression patterns during somatic embryogenesis in maize (Che et al. 2006) and soybean (Thibaud-Nissen et al. 2003). Using an alternative strategy, Low et al. (2008) successfully carried out digital northern analysis of 17,599 tissue culture ESTs to identify genes associated with oil palm embryogenesis. The genes identified by Low et al. (2008) were the lipid-transfer protein family, catalase 2, defensin EGAD1 and dehydrin-like protein. However, none of the 10 ESTs short listed for quantitative real-time PCR analysis in this study were coding for these genes. The 10 ESTs were specific to the EON library and were short listed based on their similarity to genes with potential involvement in tissue culture process. These ESTs were not identified previously probably due to the limitation of standard libraries in capturing low abundance transcripts. The presence of these unique ESTs will increase the chances of identifying additional somatic embryogenesis markers, which are required for accurate prediction of embryogenic potential across tissue culture materials. The response of a leaf explant on a tissue culture medium is highly genotype-dependent (Thibaud-Nissen et al. 2003). Therefore, the search for molecular markers with reproducible expression patterns across tissue culture materials derived from different genetic backgrounds is a challenging task. It is for this reason that two genotypes were used to test the expression profiles of selected genes. Based on the quantitative real-time PCR analysis, this study identified two ESTs that can be used to screen for the embryogenic potential of callus (Figure 2a and Figure 2c). The first EST encodes for the granule-bound starch synthase, GBSS (DW248696), involved in the synthesis of starch in the amyloplast organelle (Denyer et al. 1999; Miyazawa et al. 1999). Histological studies by Kanchanapoom and Domyoas (1999) showed the accumulation of starch grains during induction of oil palm somatic embryogenesis. This may explain the up-regulation of the GBSS transcript in EC/EMB of both the genotypes tested. Another EST, putative transcription factor Myb1 (DW247830), is a DNA-binding protein. Involvement of this protein in regulation of auxin-inducible genes has been reported previously in Arabidopsis (Ruegger et al. 1998; Shin et al. 2007). As auxin is used in the induction of oil palm embryogenic cell types, it was proposed by Meilina and Ooi (2006) that this protein is likely to trigger the same pathway during oil palm somatic embryogenesis. Genotype-specific gene expression patterns Two ESTs demonstrated genotype-specific regulation of gene expression where they were expressed at higher levels in the EMB tissues from FC2185 (Figure 3). These ESTs are fertilization-independent endosperm (DW247791) and putative late embryogenesis abundant protein (DW248206), which encode for the proteins that have been implicated in the regulation of embryo development (Luo et al. 2000; Che et al. 2006). Spatial expression of these genes may be dependent on the developmental stage of the embryo. A recent microarray analysis by Che et al. (2006) showed a number of genes to be differentially expressed during the transition from mid to late embryo development in maize. In oil palm, embryoids at different developmental stages have been observed in a single solid culture medium (Rohani et al. 2000). Therefore, to observe a similar expression profile for the genes identified in this study, the embryos tested may have to be sampled at exactly the same developmental stage. Identification of differentially expressed genes across various tissues of oil palm The genes listed in Table 1 were also profiled across various tissues of oil palm. Differential expression of these genes were observed in the spear leaves (putative transcription factor Myb1), kernel at 15WAA (putative germin A) and the roots of seedling palms (putative late embryogenesis abundant protein). Among these genes, germin A demonstrated the most interesting expression pattern. This gene was detected exclusively in 15WAA kernel with minimal expression in other tissues tested (Figure 4b). The expression profile of this gene is similar to what has been reported in conifers, where high accumulation of a germin-like protein was observed in the zygotic embryos of Pinus caribaea (Neutelings et al. 1998). The putative transcription factor Myb1 was found to be predominantly expressed in the spear leaves (Figure 4a). This gene has been shown to be expressed in barley leaves containing undifferentiated plastids (Churin et al. 2003). Another gene, putative late embryogenesis abundant protein exhibited a tissue-specific expression in the root tissues sampled at the actively dividing stage (Figure 4c). The localized expression of a late embryogenesis abundant protein in the root meristemic tissues has previously been demonstrated in Phaseolus vulgaris (Colmenero-Florez et al. 1999). Importance of normalized embryoid library Using the constructed normalized embryoid cDNA library, we were able to study genes related to oil palm tissue culture not previously identified in the standard library. The usefulness of the normalization process in discovering rare mRNA species is thus obvious. In other crops, such as cassava (Lokko et al. 2007), Brassica napus (Malik et al. 2007), cotton (Tu et al. 2007), citrus (Terol et al. 2007) and watermelon (Levi et al. 2006), genes related to economically important traits have been isolated from normalized cDNA libraries and characterized. Furthermore, the high number of unique genes available through the normalized library also enhances the assembly of an EST database. This database would be useful for high throughput assay systems, such as DNA microarray, for the analysis of gene expression profiles. Efforts are on-going to further characterize additional genes associated with oil palm tissue culture. The collection of ESTs from the normalized embryoid library will serve as a suitable pool for mining candidate genes related to somatic embryogenesis. These candidate genes can also be used as probes for genetic mapping to determine the genomic loci associated with embryogenesis. Combining data from genome and transcriptome analysis will help enhance our understanding on the molecular mechanism associated with the tissue culture process. This will assist in the development of diagnostic tools for predicting tissue culture amenity in oil palm. Acknowledgments The authors would like to thank the Director-General of MPOB for permission to publish this paper. We are indebted to FELDA Agricultural Services Sdn. Bhd. for providing the tissue culture materials. Special thanks to Mr. Andy Chang Kwong Choong for his valuable comments on the manuscript. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Supported by UNESCO / MIRCEN network Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10010f2.jpg] [ej10010ts1d.jpg] [ej10010f4.jpg] [ej10010t3.jpg] [ej10010tS1e.jpg] [ej10010tS1c.jpg] [ej10010t2.jpg] [ej10010ts1c.jpg] [ej10010t7.jpg] [ej10010ts1a.jpg] [ej10010f1.jpg] [ej10010t6.jpg] [ej10010tS1b.jpg] [ej10010f3.jpg] [ej10010t1.jpg] [ej10010t4.jpg] [ej10010tS1d.jpg] [ej10010ts1e.jpg] [ej10010tS1a.jpg] [ej10010t5.jpg] [ej10010ts1b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}