 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 2, March 15, 2010 Simultaneous saccharification and fermentation process of different cellulosic substrates using a recombinant Saccharomyces cerevisiae harbouring the β-glucosidase gene Verônica Ferreira1 , Mariana de Oliveira Faber2 , Sabrina da Silva Mesquita3 , Nei Pereira Jr.*4, 1 Laboratorios de Desenvolvimento de
Bioprocessos,
Departamento de Engenharia Bioquímica,
Universidade Federal do Rio de
Janeiro,
Rio de Janeiro, Brasil Financial support: Brazilian Petroleum Company (PETROBRAS); The Brazilian Council for Research (CNPq) and the Rio de Janeiro Foundation for Science and Technology (FAPERJ). Received May 11, 2009 / Accepted October 14, 2009 Code Number: ej10019 Abstract In Brazil, the production of ethanol from sugarcane produces large amounts of lignocellulosic residues (bagasse and straw), which have been driving research and development for the production of second generation ethanol. In the present work, a recombinant Saccharomyces cerevisiae strain expressing the β-glucosidase gene from Humicola grisea was used for ethanol production from three different cellulosic sources by simultaneous saccharification and fermentation. Initially, a enzymatic pre-hydrolysis step was done with a solid:liquid ratio of 1:4, and an enzymatic load of 25 filter paper activity (FPU).g-1 of cellulosic substrate. Using sugarcane bagasse pretreated cellulignin, crystalline cellulose and carboxymethyl cellulose, 51.7 g L-1, 41.7 g L-1 and 13.8 g L-1 of ethanol was obtained, respectively, at the end of 55 hrs of fermentation. The highest ethanol productivity (0.94 g L-1 hrs-1) was achieved using sugarcane bagasse pretreated cellulignin. The use of a recombinant S. cerevisiae led to extremely low glucose concentrations when compared to other works reported in literature. Keywords: bioethanol, lignocellulosic biomass, sugarcane bagasse derived cellulignin.
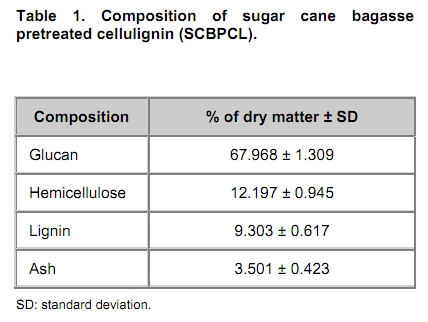
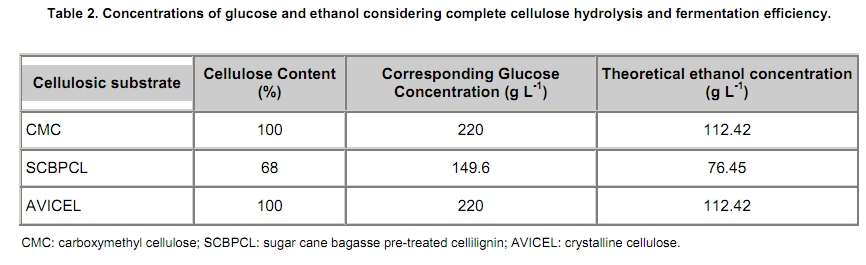
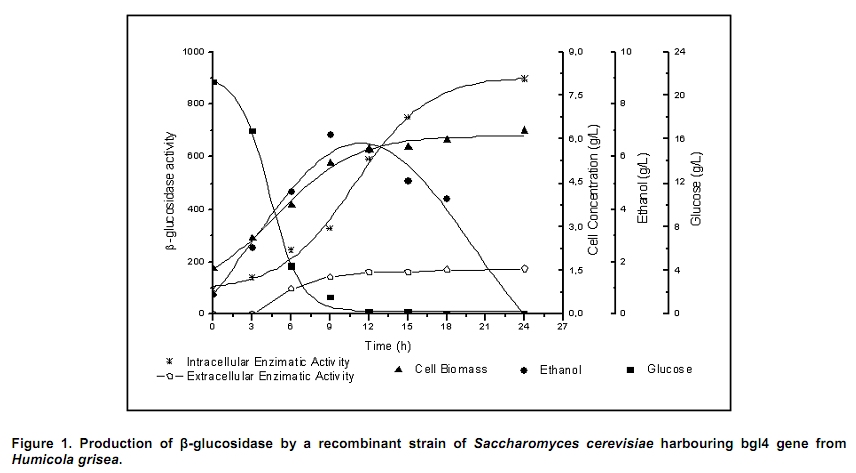
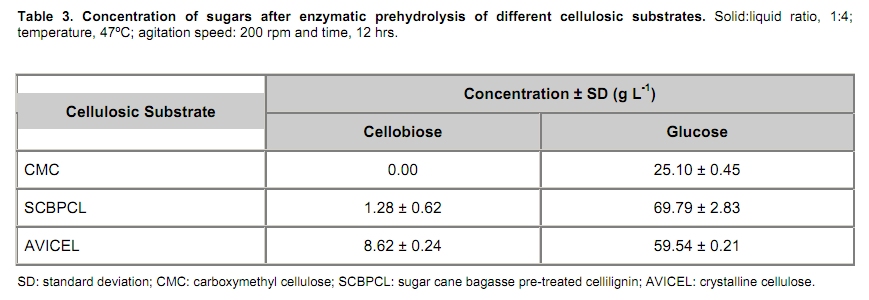
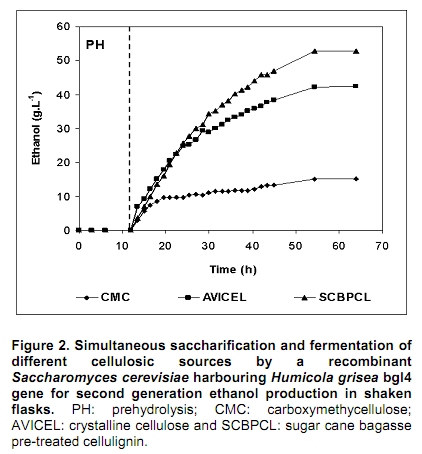
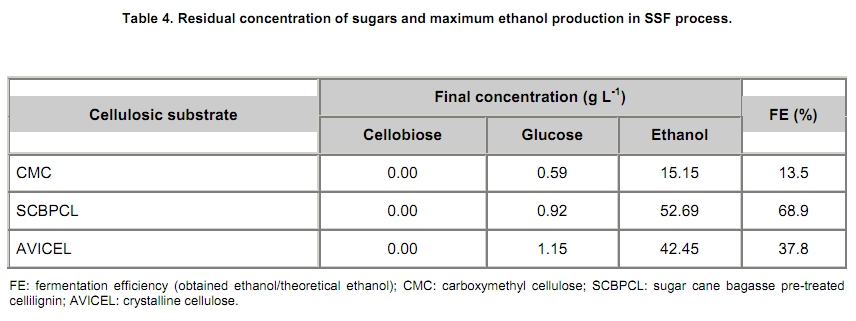
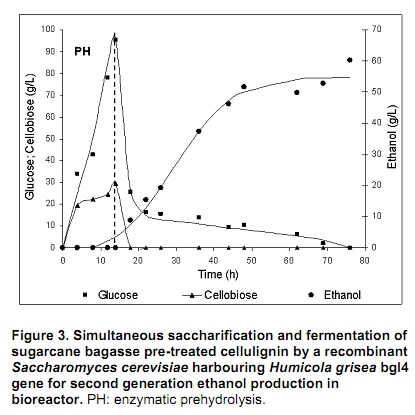
The search for new technologies for converting plant biomass into alternative biofuels is leveraged by many social and environmental problems associated with the use of fossil fuels and their exploration. Bioethanol is one the most promising biofuels from renewable resources. Fermentation-derived ethanol can be produced from sugar, starch or lignocellulosic biomass. Sugar and starch based feedstocks are currently predominant at the industrial level and they are so far economically favorable (Krishna et al. 2001; Shen et al. 2008; Sukumaran et al. 2009). In Brazil, bioethanol is produced by the sucrose fermentation from sugarcane. Great amounts of sugarcane bagasse are produced in this process. In the 2007/2008 harvest season, 496 Millions of metric tons of sugarcane were processed, producing 22 Million cubic meters of bioethanol (MAPA - www.agricultura.gov.br). The sugarcane bagasse has a complex structure, composed of 40-50% cellulose, 25% hemicellulose and 25% of lignin. Its conversion into fermentable sugars is possible using thermal pre-treatment, followed by chemical or enzymatic hydrolysis (Hernández-Salas et al. 2009). One of the routes for second generation ethanol production from sugar cane bagasse starts with a thermal pre-treatment, which aims at disorganizing and fractioning the lignocellulosic complex. This pre-treatment can be integrated with an acid catalyzed step in mild conditions as for hydrolyzing the hemicellulose component, generating a pentose-rich liquid phase, which can be fermented by an adequate microorganism (e.g., Pichia stipitis). The remaining solid material is composed mainly of cellulose and lignin, named cellulignin. This solid residue undergoes an alkaline delignification process to ensure the increase of the cellulose saccharification efficiency. Cellulose saccharification can be carried out either separately (SHF process) or simultaneously (SSF process) to the hexose fermentation step (Chen et al. 2007; Olofsson et al. 2008). Cellulose, the major component of plant biomass, is a polysaccharide enzymatically hydrolysable by three different cellulolytic enzymes: endoglucanases, exoglucanases and β-glucosidases (Vásquez et al. 2007). It is well documented that this enzymatic complex system is inhibited by its final hydrolysis products, particularly by glucose (Sukumaran et al. 2009). The Simultaneous Saccharification and Fermentation processes, firstly described by Takagi et al. (1977), combines enzymatic hydrolysis of cellulose with simultaneous fermentation of its main derived sugar (glucose) to ethanol. In this process, the stages are virtually the same as in the separate hydrolysis and fermentation system, except that both are performed in the same bioreactor. Thus, the presence of yeast together with the cellulolytic enzyme complex reduces the accumulation of the inhibiting sugars within the reactor, thereby increasing the yield and the saccharification rates. Another advantage of this approach is that a single bioreactor is used for the entire process, therefore reducing the investment costs. In addition, the presence of ethanol causes the medium to be less vulnerable to invasion by undesired microorganisms (Ballesteros et al. 2004; Olofsson et al. 2008). The present study aimed at evaluating the simultaneous saccharification and fermentation of three different cellulosic sources (carboxymethyl cellulose, crystalline cellulose and sugar cane bagasse pre-treated cellulignin) using a recombinant Saccharomyces cerevisiae harbouring the β-glucosidase gene from Humicola grisea. Materials and Methods A strain of Saccharomyces cerevisiae containing the β-glucosidase gene from Humicola grisea was obtained from the Molecular Biology Group of Brasilia University (Brazil) and was kindly provided to be use in this work. The bgl4 gene from Humicola grisea was cloned in the pGEM-T Easy (Promega) plasmid and the pYBG4 plasmid was used to transform the Saccharomyces cerevisiae yeast. Active cultures for inoculation were prepared by growing the organism on a rotary shaker at 200 rpm for 18 hrs at 30ºC, in a growth medium containing: yeast nitrogen base (Difco), 0.34 g L-1; aspartic/glutamic (Vetec), buffer 0.4 g L-1 and glucose (Vetec), 20 g L-1. The enzymatic production was performed in an instrumented 2 L bioreactor model Biostat® (B. Braun Biotech International), with control of temperature, pH, dissolved oxygen level and foam. After the yeast growth, the cell suspension was centrifuged and resuspended in 1 L of medium for enzymatic production (yeast extract (Difco), 5.64 g L-1; aspartic/glutamic (Vetec) buffer, 0.4 g L-1 and glucose (Vetec), 20 g L-1). The medium temperature was maintained at 37ºC and pH adjusted to 6.0 with automatic addition of a 2M NaOH solution. The dissolved oxygen concentration in the medium was fixed at 30% saturation, through agitation in the range of 200-550 rpm and air flow rate in the range of 0.5-1.0 L min-1. The cellulosic sources evaluated were: carboxymethyl cellulose (Sigma), crystalline cellulose (Reagen) and sugar cane bagasse pre-treated cellulignin, which was obtained after diluted acid pre-treatment of sugar cane bagasse (H2SO4, 1%v/v; solid:liquid ratio, 1 g:2 mL; temperature, 121ºC; and time of 40 min), followed by alkaline delignification (NaOH, 4% w/v; solid:liquid ratio, 1 g:20 mL; temperature, 121ºC; and time, 30 min) of the remaining solids, as reported by Vásquez et al. (2007). The contents of sugars, lignin and ash were determined according to the National Renewable Energy Laboratory (Sluiter et al. 2004) and Ververis et al. (2007). Cellulase complex derived from Trichoderma reesei (MULTIFECT®) was provided by Genencor (Danisco Division). Simultaneous saccharification and fermentation assays The SSF experiments were carried out batchwise in 250 mL conical flasks and in a 1 L bioreactor (Bioflo III, New Brunswick Scientific Co.). The conical flasks and the bioreactor were added with 10 g and 100 g dry cellulosic source, and 40 mL and 400 mL of sodium citrate buffer pH 5.0, respectively, and the media sterilized at 121ºC/20 min. A cellulase load, corresponding to 25 FPU.g-1 dry cellulosic source,was transferred to the systems, prior to the SSF process as for an enzymatic hydrolysis pre-treatment, which was performed at 47ºC for 12 hrs. Thereafter, the temperature was reduced to 37ºC and a yeast cell suspension was added into the fermentation systems in a concentration of 2 g L-1. Recombinant cells were subjected to successive washes with sodium acetate buffer pH 6.0 followed by freezing and subsequent shearing with glass beads by vortex. The intracellular extract was stored at 4ºC for subsequent quantification of the intracellular β-glucosidase activity. Filter paper activity (FPU) was measured as described by Ghose (1987). One International Filter Paper Unit of enzyme activity was defined as the amount of enzyme required for producing one micromol of glucose per minute. For the determination of β-glucosidase activity in cellobiose, 50 µL of enzymatic extract (intra or extracellular), previously diluted, were incubated with 50 µL in a 15 mM cellobiose solution, pH 6.0 for 30 min at 40ºC in a thermostatized bath (Dubnoff). After the reaction time, the enzyme deactivation (100ºC for 10 min) was done followed by the quantification of glucose concentration using the enzymatic-colorimetric assay (GOD-POD). The cell concentration was determined using the correlation between dry cell concentration and absorbance at 600 nm. Cellobiose, glucose and ethanol were analyzed with HPLC Waters® chromatograph equipped with a refractive index detector using a Bio-Rad aminex HPX-87H column at 60ºC and water as mobile phase at a flow rate of 0.4 mL min-1. Results and Discussion Glucan contents in the cellulosic sources The composition of sugar cane bagasse pre-treated cellulignin is present in Table 1. These values were used in the calculation of the sugar recovery and ethanol yield. Considering complete cellulose hydrolysis and fermentation efficiencies, the theoretical glucose and ethanol concentrations obtained in the spent media are displayed in Table 2. These values are of importance as for estimating the practical fermentation efficiency. The recombinant β-glucosidase production in bioreactor was carried out batchwise in aerobic conditions, with the objective to analyze the cell growth and the enzymatic production rates (Figure 1). The specific cell growth rate between 3 and 9 hrs was 0.116 hrs-1. With an average consumption rate of 2.27 g L-1 hrs-1, after 9 hrs of cultivation, glucose was nearly depleted, and the specific growth rate was reduced significantly. The recombinant strain expresses β-glucosidase constitutively, producing mainly intracellular activity, and in a less extent extracellular activity. The total enzymatic production (intracellular and extracellular) was roughly 1100 U L-1 at the end of 24 hrs of cultivation, corresponding to an enzymatic yield on grown cells (YP/X) of 170 U g-1. First, the recombinant strain expressed the enzyme synthesis (intra and extracellular) concurrently with cell growth and ethanol production (fermentative phase). The intracellular activity continued to be expressed even after total glucose depletion, when the cell consumed ethanol as carbon source (aerobic phase). On the other hand, extracellular activity was observed up to 10 hrs of cultivation, remaining constant thereafter. The results point out this strain of Saccharomyces cerevisiae, expressing the β-glucosidase gene, with great potential for ethanol production by the simultaneous saccharification and fermentation process (SSF), since this particular strain is capable of supplying the medium with β-glucosidase, enriching the commercial enzymatic blends used in the SSF process. Chen et al. (2007) and Hodge et al. (2008) report the need to supplement commercial cellulases used in SSF process with β-glucosidase. Vásquez et al. (2007) also observed the accumulation of cellobiose nearly to 10 g L-1 during the 10 first hours of fermentation, when using the bakery S. cerevisiae strain for bioethanol production in SSF process with sugarcane bagasse pre-treated cellulign, and had to add additional β-glucosidase in their experiments to reduce the cellobiose concentration. Simultaneous saccharification and fermentation assays with the recombinant yeast Experiments in conical flasks. The SSF process was preceded by 12 hrs-enzymatic prehydrolysis, after which the concentrations of glucose and cellobiose generated from different cellulosic substrates were determined (Table 3). While carboxymethyl cellulose (CMC) prehydrolysis produces exclusively glucose, sugar cane bagasse pre-treated cellulignin (SCBPCL) and crystalline cellulose (AVICEL) prehydrolyses generate both glucose and cellobiose. It is likely that the amount of glucose produced using SCBPCL and AVICEL led to the inhibition of β-glucosidase, leaving a residual cellobiose concentration. According to Öhgren et al. (2007), β-glucosidases are strongly inhibited by glucose, while endo- and exo-glucanase are inhibited by cellobiose. The difference between the produced sugar concentrations with SCBPCL and AVICEL in comparison with CMC in the prehydrolysis step is due to the type of cellulose and the enzymatic activities present in the commercial pool, being also strongly dependent on the sort of lignocellulose pre-treatment (Hodge et al. 2008). Sukumaran et al. (2009) also obtained different fermentable sugar concentrations at the end of the prehydrolysis process using water hyacinth biomass, rice straw and sugar cane bagasse, resulting in 14.2 g L-1, 26.3 g L-1 and 17.8 g L-1, respectively, indicating the close relationship between enzymatic pools and cellulosic sources. After the prehydrolysis each cellulosic substrate was inoculated with the recombinant Saccharomyces cerevisiae and the ethanol production was monitored for 65 hrs of SSF process (Figure 2). As it can be expected SCBPCL and AVICEL were the best substrates for SSF process, resulting in higher values of ethanol concentration when compared with CMC, since the former cellulosic sources resulted in higher available fermentable sugars than the latter source in the prehydrolysis step (Table 3). Cellobiose, glucose, ethanol concentrations and fermentation efficiencies, at the end of the SSF process, for the three investigated cellulosic sources are shown in Table 4. In all of them, celobiose was totally converted in glucose, and the values of volumetric productivity were 1.18 g L-1 hrs-1, 0.95 g L-1 hrs-1 and 0.33 g L-1 hrs-1 for SCBPCL, AVICEL and CMC, respectively, at the 43th hrs after cell inoculation. The presence of glucose in the medium at the end of the SSF process indicates the continuation of the catalytic activity of the celullase complex (Ballesteros et al. 2004), as it was noticed in the experiments herein done (Table 4). Bench bioreactor experiments. Since SCBPCL provided the best results in shaken flasks for both enzymatic prehydrolysis and SSF process, this cellulosic source was selected for experiments with the recombinant yeast to evaluate its performance in bioreactor, and quantify more accurately the process variables. At the end of the enzymatic prehydrolysis, the medium contained 95.5 g L-1 of glucose and 29.5 g L-1 of cellobiose. The bioreactor was then inoculated with 2 g L-1 of the recombinant S. cerevisiae (Figure 3), starting the SSF process. After 78 hrs, an ethanol concentration of 60 g L-1 was achieved and no accumulation of cellobiose was observed, demonstrating the effective action of the β-glucosidase expressed by the recombinant strain. The ethanol production in bioreactor also validated the results obtained using shaken flasks, resulting in similar performance (average volumetric productivity of 1 g L-1 hrs-1 for both systems). The results obtained in the present work are of interest, since they bring the solution for reducing the residual cellobiose concentrations normally found in SSF processes of lignocellulosic biomass. Besides this, the ethanol concentrations and the ethanol efficiency achieved are far greater than others reported in recent literature. Hernández-Salas et al. (2009) achieved 12.5 g L-1 of ethanol using the SSF process with sugarcane bagasse previously acid-pre-treated and NaOH-delignified, after 48 hrs of process, corresponding an ethanol efficiency of 32.6%. Similar results were obtained by Sukumaran et al. (2009), achieving at the end of SSF process an ethanol concentration of 12.3 g L-1 using rice straw pre-treated with alkali and enzymatically hydrolyzed, after 24 hrs of process, which corresponded to an ethanol efficiency of 40.3%. Concluding Remarks The major bottleneck in the second-generation bioethanol production is the high cost of the cellulolytic enzymes. Most commercially available enzymatic pools do not show sufficient β-glucosidase activity for an efficient simultaneous saccharification and fermentation process. In this work, a recombinant S. cerevisiae strain harbouring the blg4 gene from Humicola grisea was used. Apart from producing ethanol in high levels, the recombinant strain can complement the used commercial enzymatic pools with β-glucosidase, and thus guaranteeing the total conversion of cellobiose to glucose during SSF process. The β-glucosidase production by submerged fermentation carried out batchwise resulted in a global enzyme activity (intra and extracellular) of 1100 U L-1. The intracellular enzyme continued to be produced even after glucose depletion, while the extracellular activity was detected only when there was glucose in the medium. This fact resulted in a boost in the use of this strain not only for enzymatic production but also for the ethanol fermentation by the SSF process, which glucose is continuously generated in the production medium, assuring the extracellular expression of β-glucosidase. The best results of ethanol production using SSF were obtained with the most complex cellulosic source (sugar cane bagasse pre-treated cellulignin), achieving 51.7 g L-1 and 60 g L-1, in shaken flasks and in bioreactor, respectively. These results show the biotechnological potential of the recombinant strain of S. cerevisiae for the production of second generation ethanol. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication Supported by UNESCO / MIRCEN network Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10019f3.jpg] [ej10019t4.jpg] [ej10019t2.jpg] [ej10019t1.jpg] [ej10019f2.jpg] [ej10019t3.jpg] [ej10019f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}