 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 3, May 15, 2010 Short Communication EST sequencing and SSR marker development from cultivated peanut (Arachis hypogaea L.) Guo Qi Song1, Meng Jun Li2, Han Xiao3, Xing Jun Wang*4,Rong Hua Tang5, Han Xia6, Chuan Zhi Zhao7, Yu Ping Bi8
1, 2, 3, 6, 7 High-Tech Research Center,
Shandong Academy of Agricultural
Sciences,
Key Laboratory for Genetic
Improvement of Crop, Animal and Poultry of Shandong Province,
Key Laboratory of Crop Genetic
Improvement and Biotechnology, Huanghuaihai,
Ministry of Agriculture,
Ji′nan 250100, People’s Republic of China Code Number: ej10029 Making use of the gene resources of wild type peanuts is a way to increase the genetic diversity of the cultivars. Marker assisted selection (MAS) could shorten the process of inter-specific hybridization and provide a possible way to remove the undesirable traits. However, the limited number of molecular markers available in peanut retarded its MAS process. We started a peanut ESTs (Expressed Sequence Tags) project aiming at cloning genes with agronomic importance and developing molecular markers. In this study we found 610 ESTs that contained one or more SSRs from 12,000 peanut ESTs. The most abundant SSRs in peanut are trinucleotides (66.3%) SSRs and followed by dinucleotide (28.8%) SSRs. AG/TC (10.7%) repeat was the most abundant and followed by CT/GA (9.0%), CTT/GAA (7.4%), and AAG/TTC (7.3%) repeats. Ninety-four SSR containing ESTs were randomly selected for primer design and synthesis, of which 33 pairs could generate good amplification and were used for polymorphism assessment. Results showed that polymorphism was very low in cultivars, while high level of polymorphism was revealed in wild type peanuts. Keywords: Arachis hypogaea L., cultivars, EST-SSR, polymorphism, wild-type peanut.
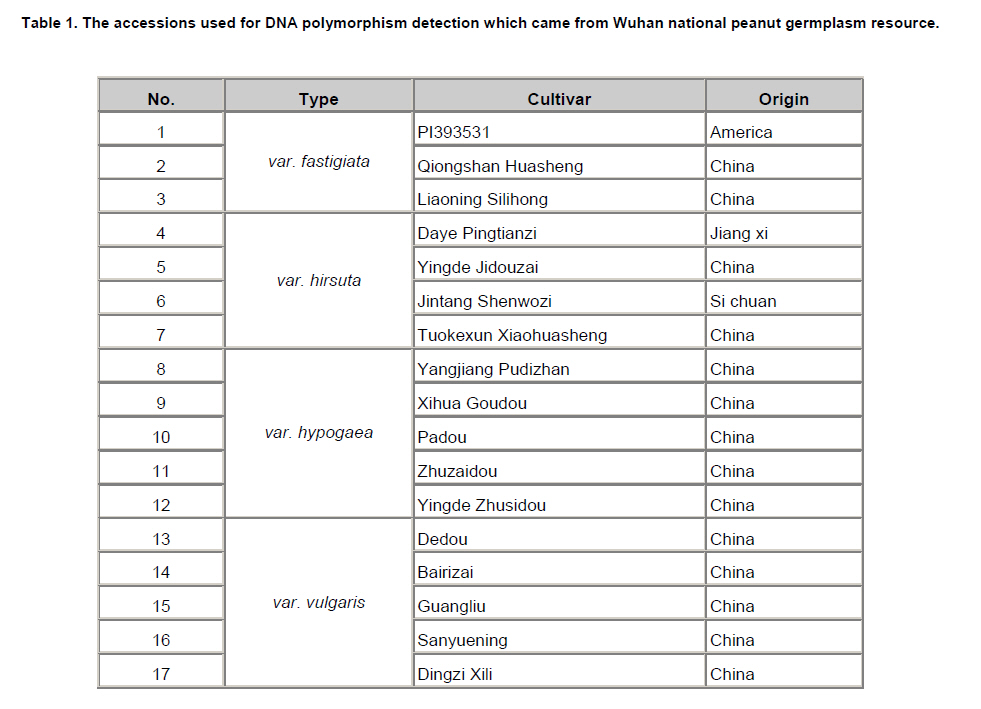
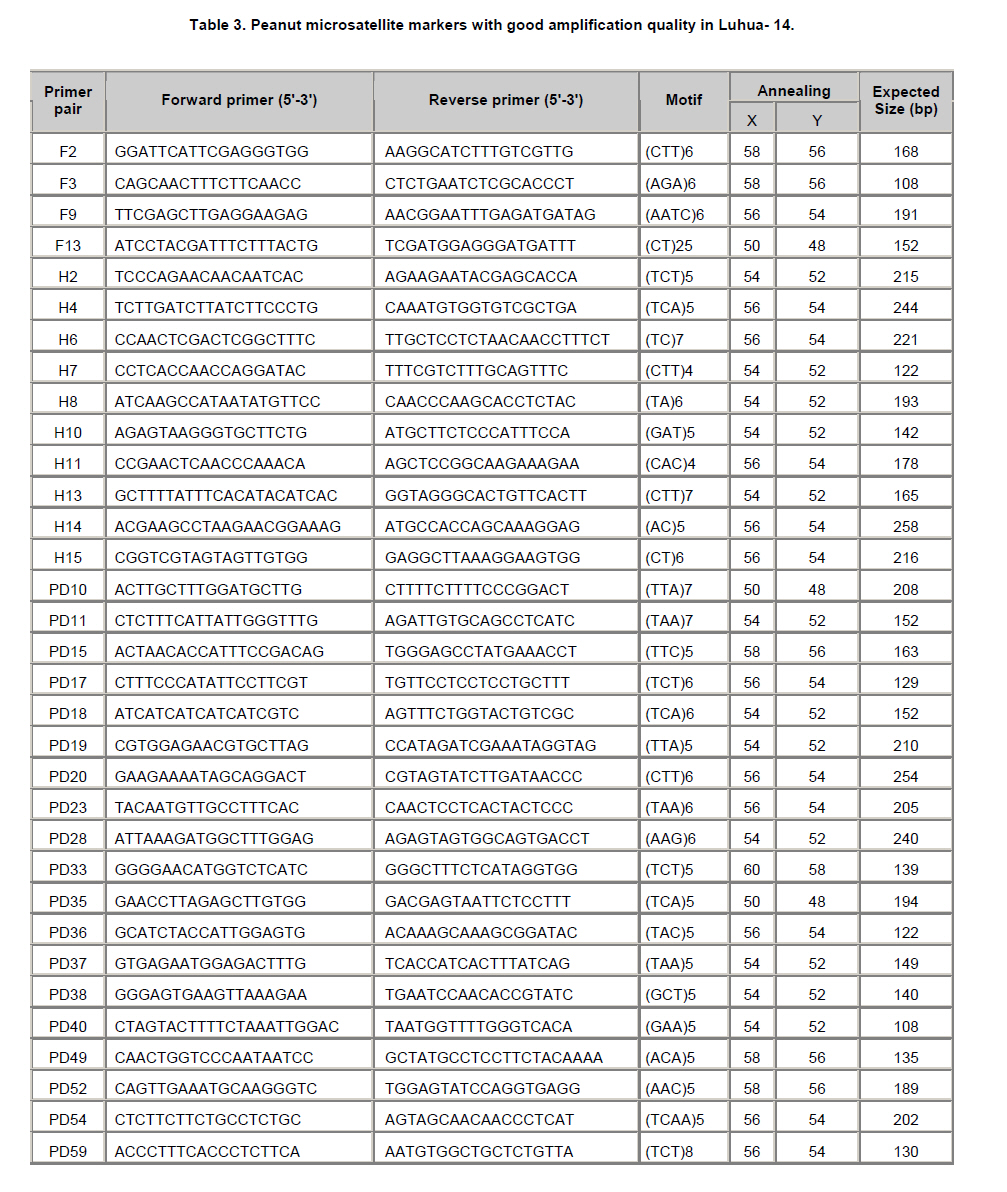
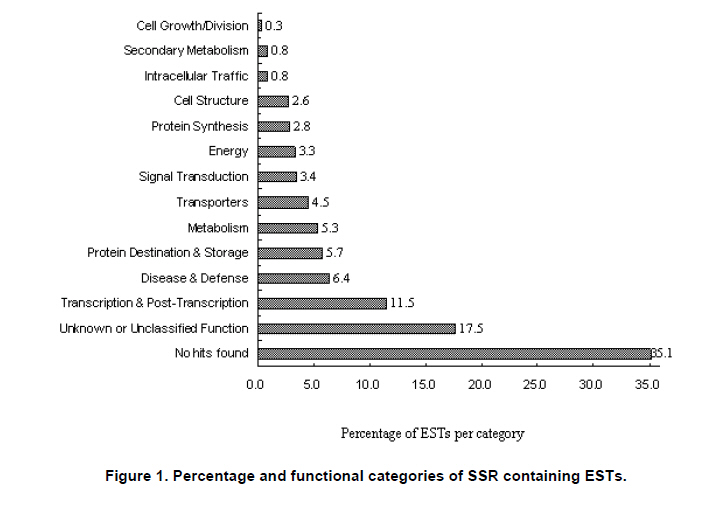
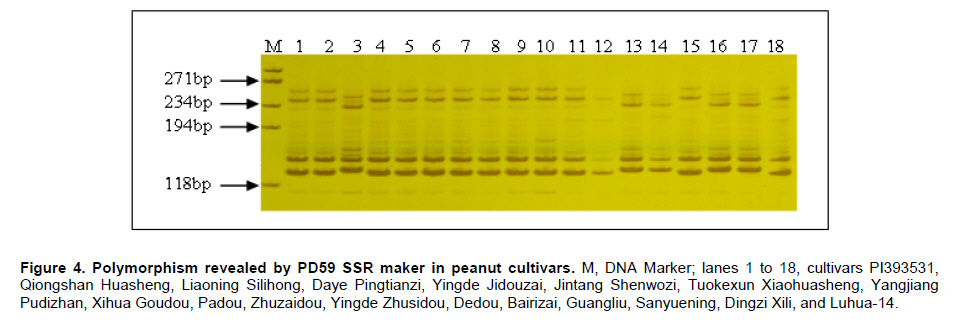
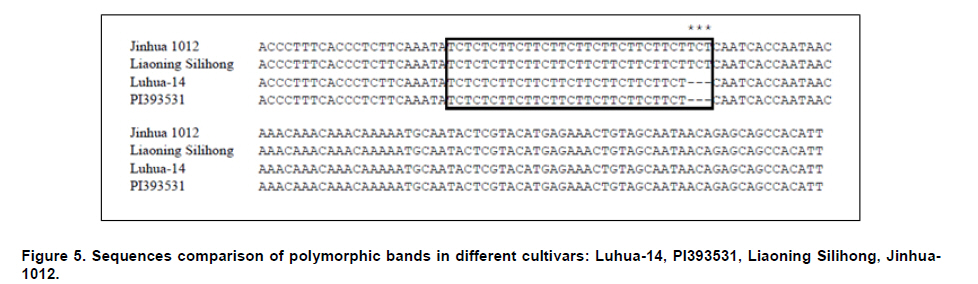
Peanut is one of the most important oil seed crops in the world, cultivated mainly in tropical, subtropical and warm temperate climates (Proite et al. 2007). The cultivated peanut is an allotetraploid (AABB, 2n = 4x = 40), originated from chromosome duplication of hybrid between AA and BB wild type species, which happened about 3500 years ago. Polyploidy and self-pollination nature of this species blocked the genetic exchange between the cultivated and the wild type species which lead to limited genetic diversity of the major cultivars. Using of only very few central germplasms in breeding programs further limited the addition of genes with agronomic value. Due to its limited genetic variation peanuts are vulnerable to a wide variety of pathogens and abiotic stresses. For example, fungal foliar diseases of peanut such as rust, web blotch, and leaf spot cause severe loss of yield worldwide (Leal-Bertioli et al. 2009). Wild type peanuts are genetically very diverse and selectively accumulated biotic and abiotic stress resistant genes during their adaptation to different harsh environments (Holbrook and Stalker, 2003). However, hybrids between the diploid wild type and the allotetraploid cultivated peanut are sterile (Tallury et al. 2005). In order to obtain tetraploid fertile offspring of the diploid wild type and the allotetraploid cultivated peanut, complicated breeding programs have to be designed. It normally takes many years to get a hybrid with agronomical value (Simpson et al. 1993; Stalker and Beute, 1993; Simpson and Starr, 2001). Marker assisted selection could increase the efficiency and predictability during the process of wild gene pool utilization. However, it was retarded by the small number of peanut molecular markers and the unavailability of high density linkage map (Burow et al. 2001; Moretzsohn et al. 2005). The attempt to develop more peanut molecular markers for construction of high density linkage map is of great importance. Microsatellites (SSRs) as DNA markers are highly polymorphic, highly abundant, show co-dominant inheritance, analytically simple and readily transferable between different plant species (He et al. 2003). Previous studies showed that SSR markers could detect more polymorphism in peanut than other molecular markers like RFLPs (Gimenes et al. 2007), AFLPs (He and Prakash, 2001; Gimenes et al. 2002) and RAPDs (Dwivedi et al. 2001; Subramanian et al. 2000). Beside sequencing SSR enriched genomic library for SSR marker development (He et al. 2003; Han et al. 2006), searching SSRs using peanut EST information is an alternative way for marker discovery (Wang et al. 2006; Proite et al. 2007; Liang et al. 2009). We started an EST project using immature peanut seeds cDNA library of a Chinese cultivar to generate more sequence information for gene cloning (Li et al. 2009, Li et al. 2010) and marker development. The objectives of the present study were to develop more SSR markers from peanut using these EST sequences, and to detect the polymorphisms of the newly identified SSR markers in a collection of cultivated and wild type peanut accessions. The resulted SSR markers especially those showed polymorphism in different peanuts accessions would be valuable for peanut marker linkage map construction and marker assisted selection in the future. Materials and MethodsLuhua-14 peanut used for cDNA library construction was obtained from Shandong Academy of Agricultural Sciences. A total of 98 accessions of wild type and cultivated peanuts were examined in this study. Twenty-five accessions belonging to section Arachis or Heteranthae which come from Nanning branch of the national wild peanut germplasm resource were used for polymorphism assessment. Seventy-three accessions belonging to cultivated species were used for genetic diversity analysis. Of these, fifty-six were collected from different provinces and the other 17 were from Wuhan national peanut germplasm resource belonging to var. fastigiata, var. hirsuta, var. hypogaea and var. vulgaris (Table 1 and Table 2). Peanut cDNA library construction The peanut seeds of different growing stages ranging from 20 to 60 DAP (days after pegging) were collected and placed into liquid N2 immediately and stored in -80ºC freezer. RNA was extracted using RNAgent kit (Promega) according to the manufacturer’s instructions. The quantity and quality of RNA was evaluated by spectrophotometry and agarose gel electrophoresis. cDNA synthesis and library construction followed the protocol of Stratagene's pBluescript II cDNA library construction kit. Plasmid DNA was isolated from the randomly selected colonies and preserved in 96-well plate. Plasmid DNA was used as template for PCR amplification using T3 and T7 universal primers. PCR products were purified using AxyPrepTM PCR Cleanup Kit (AXYGEN), and sequenced using BigDye Terminator Cycle Sequencing Kit. Sequencing was performed on 3730XL Sequencer (ABI) using T3 or T7 primers. Sequences were edited by the software SEQENCHER and vector sequences were trimmed manually. Analysis of microsatellites and primer design EST sequences were searched for SSRs using SSR tool software (http://www.gramene.org/db/searches/ssrtool) to identify di- and trinucleotide and tetranucleotide SSR motifs. SSR motifs repeated more than five times in dinucleotide, four times in trinucleotide and tetranucleotide were counted. Primers were designed using Primer 5 software. DNA extraction and PCR amplification Seventy-three cultivated peanuts and 25 wild type peanuts were used for polymorphism study. Genomic DNA was extracted from young leaf tissue using CTAB method (Murray and Thompson, 1980). DNA concentration was determined by spectrophotometer. PCR mixture consisted of 1X PCR buffer, 200 µM of each dNTP, 1U Taq polymerase (TaKaRa) and 0.5 µM of each primer and 40-60 ng of template DNA. The thermocycling conditions started with 94ºC for 5 min, 10 cycles of 94ºC for 45 sec, XºC for 45 sec, 72ºC for 45 sec, and then 30 cycles of 94ºC for 45 sec, YºC for 45 sec, 72ºC for 45 sec followed by a terminal extension step of 72ºC for 10 min. The annealing temperatures (X and Y) were optimized for each primer to allow the best amplification (Table 3). Amplification products were visualized on 6.5% w/v denaturing polyacrylamide gels (19:1 acrylamide/bisacrylamide, 7 M urea) that were silver stained (Gao et al. 2009). Product size was estimated based on Φ×174-HaeIII digest DNA Marker (TaKaRa) or Bio DL100 (Bio Flux). Results and DiscussionSequence analysis and SSR identification A cDNA library was constructed using mRNA from immature seeds at different developmental stages of cultivated peanut Luhua-14. The insert length of the library ranged from 500-1500 bp with an average length of 900 bp based on PCR results for >1000 randomly picked colonies (data not shown). Sequences were edited by the software SEQENCHER and vector sequences were removed manually. Short sequences (<200 bp) and sequences with many unreadable nucleotides were discarded. 12,000 ESTs with good quality including the 7,456 ESTs deposited to GenBank (EE123340-EE127745; EG372473-EG374270; EG529454-EG530705) were subjected for SSR discovery. From these ESTs 610 SSR containing sequences were identified. A homology search against NCBI protein database was conducted by BLASTX with an E value of 0.02 as the cut-off threshold. Based on the BLAST results 610 SSR containing ESTs were grouped into 14 categories (Figure 1). More than half of these ESTs have no hits (35.1%) and corresponding unknown proteins (17.5%) in the database. The third largest functional category (11.5%) was transcription and post-transcription related factors, followed by disease and defense (6.4%), protein destination and storage (5.7%) and metabolism (5.3%) related genes. Among 610 SSR-containing ESTs, 436 (71.5%) contained single SSR, 132 (21.6%) contained two SSRs, and the rest (6.9%) contained more than two SSRs. Among 841 SSRs identified, 66.3% were trinucleotides, followed by dinucleotides (28.8%), tetranucleotides (3.2%), pentanucleotides (0.8%), and hexanucleotides (0.8%). AG/TC was found to be the most common repeat motif (10.7%), followed by CT/GA (9.0%), CTT/GAA (7.4%), AAG/TTC (7.3%), AGA/TCT (6.2%), and AT/TA repeat motifs (5.4%). Previous surveys carried out on microsatellite abundance analysis in plant genomes have shown ATT and CTT were the most frequent trinucleotide repeat motifs (Lagercrantz et al. 1993; Ferguson et al. 2004), and AT was the most abundant dinucleotide repeat motif followed by AG/CT and GT/CA (Cuc et al. 2008; Han et al. 2006). He et al. (2003) found that GA/CT repeat was the most frequently dispersed microsatellite in peanut. Cuc and coworkers indicated that GT/CA repeat motif was the most common, accounting for 37.6% of all repeat types, followed by GA/CT repeat at 25.9% (Cuc et al. 2008). From this it can be concluded that SSR motifs are not equally abundant in eukaryotic genomes and the relative abundance of different motifs varies among species (Wang et al. 1994; Han et al. 2006). Cultivated peanut Luhua-14was used to analyze the amplification quality of the identified SSRs. All 33 (35.1%) primer pairs with good amplification were selected for further analysis of polymorphism in different cultivars and wild type peanuts (Table 3, Figure 2). Polymorphism study of wild type and cultivated peanuts Twenty-five wild type peanuts were analyzed using the selected 33 SSR primer pairs. Most primer pairs could amplify distinct bands. However, some wild type peanuts failed to amplify any bands with some primer pairs. The possible reason for this result might due to the notable difference between wild type and cultivated peanuts. Initially 55 cultivated peanut genotypes were analyzed using the 33 primer pairs. Results showed that the polymorphism was very limited. Only one primer pair (PD59) could amplify polymorphic bands. In order to further evaluate the polymorphism of these 33 SSRs, seventeen cultivated peanut (from Wuhan national cultivated peanut germplasm resources) including var. fastigiata, var. hirsuta, var. hypogaea and var. vulgaris were analyzed (Table 1). Similarly, low polymorphism was revealed by these SSR markers and only 4 (12.1%) primer pairs, F9, F13, PD20 and PD59, could amplify polymorphic bands (Figure 4). For the 73 cultivated peanuts, primer pair PD59 could always amplify either one or both of two bands between 120 and 150 bp (Figure 4). The upper band showed same size within cultivated peanut accessions tested. However, the lower band displayed varied size in different cultivars (Figure 4). The lower band of four cultivated peanuts, PI393531, Liaoning Silihong, Jinhua-1012 and Luhua-14 were recovered from the agarose gel for sequence analysis. The sequencing results were shown in Figure 5. Cultivars PI393531 from America and Luhua-14 had one TCT less than Liaoning Silihong which belonging to var. fastigiata and Jinhua-1012 which belongs to var. hypogaea. AcknowledgmentsThis work was supported by grants from Shandong Province (200701004, 2006BS06008, Z2002D06) and Shandong Academy of Agricultural Sciences (2006YBS001, 2007YCX001). References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10029t2.jpg] [ej10029f1.jpg] [ej10029t1.jpg] [ej10029f3.jpg] [ej10029f5.jpg] [ej10029f4.jpg] [ej10029f2.jpg] [ej10029t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}