 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 4, July 15, 2010 Endogenous GUS-like activity in Capsicum chinense Jacq. Laura Yesenia Solís-Ramos*#1, Tomas González-Estrada¥2, Antonio Andrade-Torres3, Gregorio Godoy-Hernández4, Enrique Castaño de la Serna5 1Unidad de
Bioquímica y Biología Molecular de Plantas,
Centro de Investigación Científica de Yucatán,
Calle 43, No.130, Chuburná de Hidalgo,
C.P. 97200, Mérida, Yucatán, México,
E-mail: laura.solisramos@ucr.ac.cr Financial support: Consejo Nacional de Ciencia y Tecnología (CONACYT), México and Dirección de Intercambio Académico de la Secretaría de Relaciones Exteriores from México. Code Number: ej10034 The gene uidA, codes for β-glucuronidase, which is one of the reporters more frequently utilized in transgenic plants. However, this can only be use if the selected organism does not present endogenous GUS-like activity. In tissues of C. chinense we found a GUS-like activity showing different levels of intensity. Histochemical screening showed that endogenous GUS-like activity decreased, or reduced significantly, in almost all tissues with exception of stament, when phosphate buffer was adjusted to pH 8. Subsequently, C. chinense zygotic embryo explants were transient transformed with Agrobacterium tumefaciens LBA4404 (pCAMBIA2301) and plantlets regenerated were histochemically stained in phosphate buffer pH 8. Observations of incubated tissues of C. chinense regenerants showed blue staining, suggesting expression of uidA. Incubated tissues of non-transformed regenerants did not show blue staining in phosphate buffer pH 8. The results show that for transformation experiments of C. chinense with uidA gene, pH 8 is recommended for histochemical staining. Keywords: Agrobacterium tumefaciens, β-glucuronidase, Capsicum chinense, gene uidA.
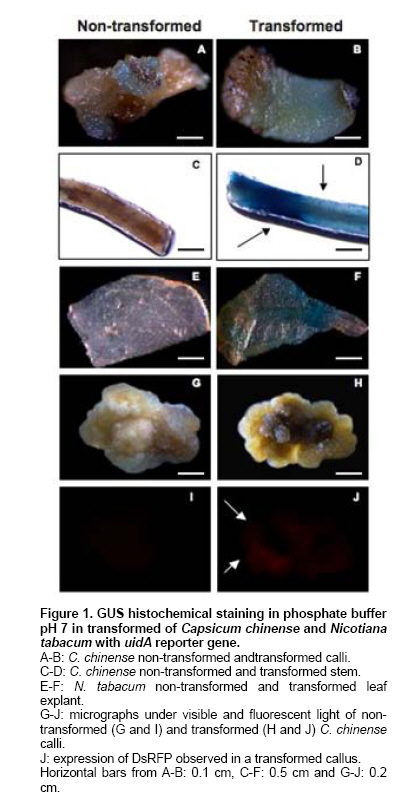
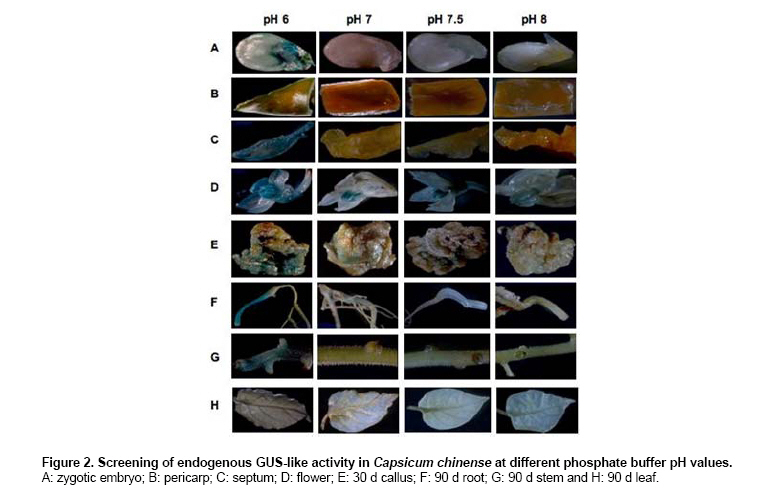
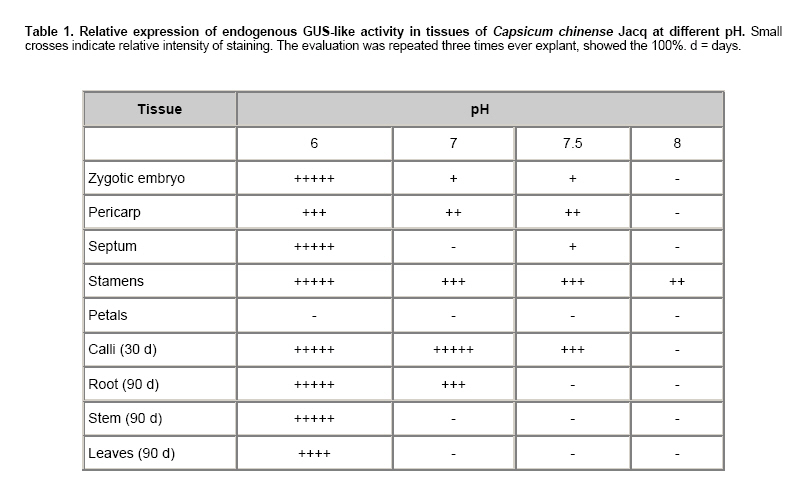
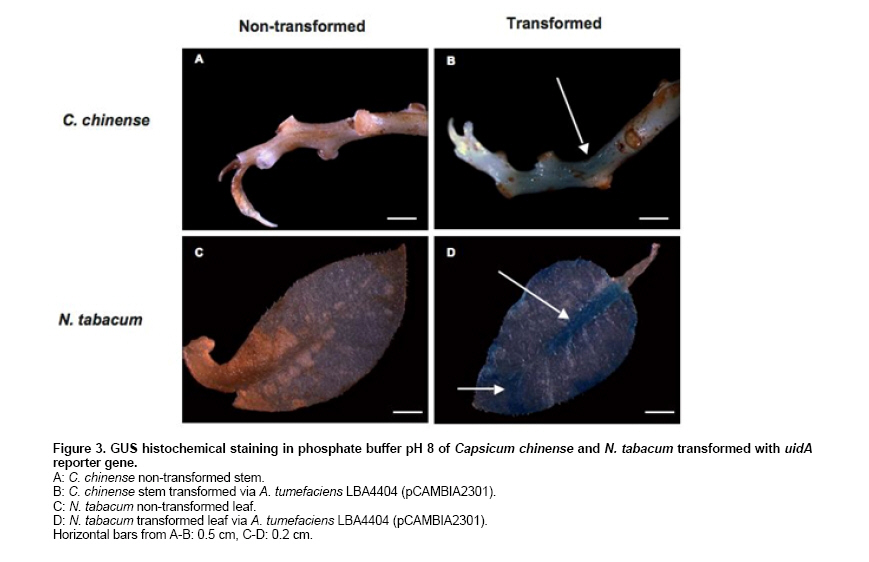
Habanero chili (Capsicum chinense Jacq.) is a species of economic importance to Mexico. The Capsicum genus is recalcitrant in in vitro morphogenesis protocols, complicating regeneration and genetic transformation (reviewed by Ochoa-Alejo and Ramírez-Malagón, 2001). C. chinense is no exception (Santana-Buzzy et al. 2005; López-Puc et al. 2006), and no efficient, reproducible regeneration system has yet been developed for this species. A dependable regeneration system is indispensable for genetic improvement, and is also important to conduct studies to establish a reliable genetic transformation protocol, as it can be an alternative to abate recalcitrance of tissues to in vitro regeneration, besides being a useful tool for genetic improvement (Zuo et al. 2002; Solís-Ramos et al. 2009). The gene uidA codes for β-glucuronidase which is utilized as a reporter in plant genetic transformation because it is generally believed that higher plants do not show GUS-like endogenous activity (Jefferson, 1987; Martin et al. 1991; Sudan et al. 2006). However, several studies have demonstrated that some plant species show endogenous GUS-like activity in vegetative tissues as well as reproductive organs (Cervera, 2005; Sudan et al. 2006). The enzyme stability and high sensitivity of the GUS assay to qualitative (histochemical assay) and to quantitative (fluorometric or spectrophotometric assay) detection are some of the reasons that explain the extensive use of uidA gene in plant genetic transformation (Cervera, 2005). Therefore, in order to avoid undesirable effects in interpreting genetic transformation results, it is recommended to evaluate potential endogenous GUS-like activity in tissues that will be targeted to genetic transformation by using uidA as a reporter. The pH of the assay buffer is very critical for detection of the GUS activity in plants. The E. coli-derived GUS has optimum activity at pH 7.0 and hence plant tissues are assayed at neutral pH after transformation (Sudan et al. 2006). The histochemical staining of plant tissues for GUS activity has some disadvantages, such as a high degree of difficulty to detect activity when gene expression is low or in cases when optical density of plant tissues is high. In order to allow a better visualization of blue staining in transformed plants, it is recommended that green tissues must be treated with ethanol to eliminate chlorophylls (Rech et al. 2003). The aim of this study was to genetically transform Capsicum chinense Jacq. via Agrobacterium tumefaciens (1) to characterize GUS activity in transformed and non-transformed tissues, (2) to evaluate the effect of pH level of phosphate buffer on the endogenous GUS-like activity of non-transformed tissues, and provide means of distinguishing between the two activities in different tissues of C. chinense. Materials and MethodsTransient genetic transformation of zygotic embryo explants via Agrobacterium tumefaciens Segments of mature zygotic embryos of C. chinense were used as explants for transient transformation with A. tumefaciens LBA4404 (pCAMBIA2301) and C58C1 (pER10W-35S Red). T-DNA in pCAMBIA2301 (Center for the Application of Molecular Biology to International Agriculture, Canberra, Australia) includes a copy of Escherichia coli uidA gene under the control of CaMV35S promoter and the NOS terminator. In addition, T-DNA contains the neomycin phosphotranspherase II gene (npt) flanked by the CaMV35S promoter and the CaMV35S terminator. In this binary vector, uidA gene coding sequence is interrupted by a Castor Bean catalase intron, which has to be removed for eukaryotic expression and prevents bacterial transcriptions of the gene coding sequence. The disarmed A. tumefaciens C58C1 carrying the binary vector pER10W-35S Red (Canche-Moo et al. 2006) was used to transform C. chinense (Solís-Ramos et al. 2009) as control for transformation efficiency. The binary vector pER10W-35S Red, contains the DsRFP reporter gene under the 35S constitutive promoter, and the gene WUSCHEL under an 17 β-estradiol inducible promoter (Zuo et al. 2002). Transient transformation of C. chinense explants and plant regeneration were carried out following the protocol previously described by Solís-Ramos et al. (2009). In addition, as a positive control leaves explants of Nicotiana tabacum were transient transformed via A. tumefaciens LBA4404 (pCAMBIA2301), to verify that the protocol used for GUS activity was done properly. Histochemical staining for β-glucuronidase activity Histochemical staining of C. chinense explants was carried out following a protocol reported by Jefferson (1987) with modifications. Explants were vacuum infiltrated for 5 min in a phosphate buffer solution consisting of 100 mM NaH2P04, 10 mM EDTA, 0.5 mM potassium ferricyanide K3Fe (CN)6, 0.5 mM potassium ferrocyanide K4Fe(CN)6 3H2O, and 0.2% triton X-100. X-Gluc (5-bromo-4-cloro-3 indolyl-β-D-glucuronide) previously dissolved in DMSO was added to a final concentration of 1 mg/L. Explants were dark incubated at 37ºC for 24 hrs. After incubation, staining solution was eliminated and explants washed with 70% ethanol for 24 hrs to eliminate chlorophylls and other pigments present in treated tissues. Explants were stored in fresh 70% ethanol. Presence of blue spots was recorded and interpreted as transient GUS expression. A stereoscope (Leica MZLIII, Germany) was utilized for better visualization. Color photographs were taken with a digital camera attached to the stereoscope and connected to a PC. Analysis of variance at α = 0.05 was conducted with Sigma Stat for Windows (version 3.11) to identify statistical differences. Detection of transient red fluorescent protein activity The transient expression of red fluorescent protein was detected using a Leica MZFLIII stereoscopic microscope equipped with appropriate filtres (546/10 nm, 600/40 nm). pH effect on endogenous GUS-like activity in C. chinense We analyzed the pH effect on endogenous GUS-like activity in C. chinense, tissue explants from reproductive organs of plants grown in a greenhouse were utilized as experimental biological material including zygotic embryo, pericarp, septum, petals and stamen. In addition, vegetative tissues including calli of 30 days, roots, stems and leaves of 90 days in vitro cultured plantlets. To determinate in C. chinense tissues a condition to inhibit endogenous GUS-like activity identified when using the phosphate buffer at pH 7, a set of experiments were carried out by modifying phosphate buffer pH at 6, 7, 7.5 and 8, there were 3 replicates per treatment. Incubation conditions were followed as previously described and the number of stained explants was recorded. The relative expression of endogenous GUS-like activity in tissues of C. chinense, at different pH levels was evaluated. The endogenous GUS-like activity was defined by visual observation of the blue color provided by the staining protocol, to the higher intensity the greater number was assigned. Histochemical staining of regenerants in phosphate buffer pH 8 Zygotic embryo explants of C. chinense were transient transformed in vitro with A. tumefaciens LBA4404 (pCAMBIA2301) following a procedure reported by Solís-Ramos et al. (2009). In vitro regenerated plantlets were stained in phosphate buffer pH 8. Leaf explants of N. tabacum transformed plants via A. tumefaciens LBA4404 (pCAMBIA 2301) were included as a control for GUS staining at pH 8. ResultsGUS-activity and GUS-like activity in transient transformed and non-transformed zygotic embryo explants Successful transient transformed C. chinense zygotic embryo explants were achieved with A. tumefaciens LBA4404 (pCAMBIA2301) and the bacteria were eliminated with 1 g/L cefotaxime and 500 mg/L timentin (Solís-Ramos et al. 2009). The calli of C. chinense transient transformed with pER10W-35S Red (used as control for transformation efficiency) expressed the red fluorescent protein (DsRFP) (Figure 1J), but not the non-transformed calli (Figure 1I). Developing calli from transient transformed explants (LBA4404 pCAMBIA2301) showing no bacteria incidence after 30 days in vitro culture, showed positive GUS-staining at pH 7 (Figure 1B). Similarly, the non-transformed leaf tissue and the non-transformed developing calli, showed endogenous GUS-like activity in phosphate buffer pH 7 (Figure 1A). Stem segments of seedlings of C. chinense grown in vitro and transient transformed with the uidA gene, showed GUS activity (Figure 1D), while the non-transformed stem segments showed no GUS activity (Figure 1C). We show results with N. tabacum as a control to verify that the protocol used for GUS activity was done properly (Figure 1E-F). Analysis of variance did not report significant differences among transformed (p = 0.622) and non-transformed explants (p = 0.242) at pH 7. Therefore, GUS-staining at pH 7 of C. chinense calli developed from zygotic embryo explants did not allow to differentia between putative A. tumefaciens transformed and non- transformed tissues. pH effect on endogenous GUS-like activity in C. chinense The endogenous GUS-like activity in C. chinense tissues was defined by visual observation of the blue color provided by the staining protocol the higher the intensity. A screening for endogenous GUS-like activity in C. chinense tissues was performed in phosphate buffer adjusted to pH 6, 7, 7.5 and 8 (Figure 2 and Table 1). At pH 6 the 100% of all samples (vegetative and reproductive tissues) presented endogenous GUS-like activity. We should point that the triplicate foliar tissue tested only presented activity in vascular vessels (Figure 2H). In phosphate buffer pH 7 an intense GUS-like activity was observed in the 100% of stamen, calli and roots developing from in vitro cultivated zygotic embryo explants (Figure 2D, E, and F). In segments of zygotic embryo (non-in vitro cultivated), and pericarp (external layers) a less intense endogenous GUS-like activity was observed (Figure 2 A and B). In contrast, the 100% of septum, petals, stems and mature leaves of 90 days culture did not present endogenous GUS-like activity (Figure 2 C, D, G and H). At pH 7.5 no GUS-like activity was observed in all of the petals, root, stem or leaves (Figure 2 D, F, G and H). However, in septum, stamen and calli some GUS-like activity was observed (Figure 2 C, D and E). A substantial decrease, or even a total absence, of GUS-like activity was observed in phosphate buffer pH 8 in almost all tissue analyzed with an exception for a slight activity in stamens (Figure 2-D). Histochemical screening at pH 8 of transient transformed regenerants From C. chinense zygotic embryos transient transformed plants were regenerated in vitro (Solís-Ramos et al. 2009), and were evaluated by histochemical staining using the phosphate buffer at pH 8 (Figure 3). As a control test was also assessed leaf tissue of N. tabacum transformed and non-transformed (Figure 3 C-D). C. chinense stem segments of in vitro regenerated plantlets from transient transformed explants showed positive blue staining (Figure 3 B). Leaves of transformed N. tabacum utilized as a control, showed GUS activity which verified that the protocol used for GUS activity was done properly (Figure 3 D). In contrast, tissues of C. chinense and N. tabacum regenerated from non-transformed explants did not show GUS-like activity at pH 8 (Figure 3 A and C). Figure 3 shows that use of phosphate buffer at pH 8 in tests, does not affect the activity of the uidA gene in transient transformed tissues of C. chinense. These results suggest that uidA gene is transcriptional active in C. chinense and N. tabacum tissues of plantlets regenerated from Agrobacterium-mediated transient transformed explants. DiscussionPrevious reports of plant genetic transformation utilized reporter genes to determine whether the transformation method allowed incorporation of the transgen into the genome of interest. The uidA gene that codes for β-glucuronidase is one of de most frequently reporter gene utilized in plant transformation experiments (Cervera, 2005). In this study zygotic embryo segments of C. chinense were transient transformed with A. tumefaciens LBA4404 (pCAMBIA2301) and then histochemically stained in phosphate buffer pH 7 for GUS activity after 30 days of in vitro culture. Blue staining was observed in both transient transformed and non-transformed explants (Figure 1 A, B). This result suggested an endogenous GUS-like activity in C. chinense calli. In addition, we observed endogenous GUS-like activity in pericarp, stamen, and 30 days calli, and 90 days roots of plantlets cultivated in vitro (Figure 2 B, D, E and F). These findings coincide with results reported for other species such as Ricinus communis L. that showed GUS-like activity in reproductive tissues (Rezmer et al. 1999, Cervera, 2005). However in pollen obtained from the in vitro flowers of C. frutescens inoculated with A. tumefaciens was observed transient GUS expression (Sharma et al. 2008). In contrast, other reported species such as Bixa orellana (Zaldívar-Cruz et al. 2003), Cucurbita maxima L., Vicia faba L., Kalanchoe daigremontiana (Rezmer et al. 1999), Lycopersicon esculentum (Sun et al. 2006), Musa acuminata cv. “Grand Nain”scalps (Acereto-Escoffié et al. 2005) and Argemone mexicana L. (Godoy-Hernández et al. 2008), did not show endogenous GUS-like activity in evaluated tissues. An important prerequisite in order to utilize a reporter gene, is the absence of endogenous activity in the target organism for transformation (Martin et al. 1992). In Arabidopsis thaliana it was reported that pH of the staining solution had a strong influence in endogenous GUS-like activity (Martin et al. 1992). Furthermore, endogenous GUS-like activity at pH lower than 7 can be inhibited with 20% methanol in the incubation buffer (Rezmer et al. 1999). Based on the previous results and reports, we decided to determine the effect of pH on the endogenous GUS-like activity identified in C. chinense by modifying pH of phosphate buffer. In our experiments all evaluated tissues of C. chinense at pH 6 showed endogenous GUS-like activity (Figure 2 A-G), which is in agreement with previous reports for a wide variety of plant species that presented endogenous GUS-like activity at acidic pH in the staining buffer (Martin et al. 1992; Rezmer et al. 1999). The pH of the assay buffer was found to be critical with pH 4.0 being optimum for detection endogenous GUS-like activity, recently were demonstrated ubiquitous in the tissues (vegetative as well as reproductive) root, stem, leaves and flowers of model plant species Arabidopsis thaliana, Oryza sativa, Nicotiana tabacum and Zea mays (Sudan et al. 2006). However, by increasing pH to alkaline values in the staining solution the endogenous GUS-like activity was suppressed in most of the C. chinense tissues with the exception of stamen (Figure 2D). By culturing in vitro we regenerated plantlets from zygotic embryo segments transient transformed by A. tumefaciens, which were histochemically stained in phosphate buffer pH 8 (Figure 3), the transient transformed regenerants showed blue staining, in contrast, non-transformed regenerants did not show any blue staining under the same staining conditions. This observed GUS activity in C. chinense tissues transformed by A. tumefaciens could be due to uidA expression, successfully inserted in the plant genome (Figure 3B). However, the genetic transformation of the C. chinense tissues is considered as transient, therefore only parts of the tissues are transformed which is shown in Figure 3B and it is typical of transient transfections. A false positive bacterial gene expression is discarded since pCAMBIA2301 includes an intron that interrupts the GUS coding sequence and prevents bacterial transcriptions of the gene coding sequence. GUS activity was also observed in N. tabacum foliar tissue excised of regenerated explants that were transformed by A. tumefaciens (Figure 3D). Non-transformed negative controls regenerated plants did not show GUS activity in staining buffer pH 8 (Figure 3C). Our results of histochemical staining in phosphate buffer pH 8, suggest that uidA gene was introduced in regenerants of C. chinense and N. tabacum and the gene was transcriptional active as it can be inferred from the blue stain observed in tissues of regenerated plantlets. Additional characterization of regenerants will be conducted to verify by molecular methods these staining results. The main problem during initial steps of transformation is just to get an assay conditions which can provide an initial screening. This problem has been solved by adjusting the pH to 8 for C. chinense. AcknowledgmentsThe first author thanks to Dirección de Intercambio Académico of Secretaría de Relaciones Exteriores from México for scholarship and Centro de Investigación Científica de Yucatán for facilities and laboratory support. We wish to thank Q.F.B. Elidé Avilés Berzunza and I.I.A. Wilma González Kantún for technical help. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10034t1.jpg] [ej10034f2.jpg] [ej10034f3.jpg] [ej10034f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}