 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 4, July 15, 2010 Isolation of purple nonsulfur bacteria for the removal of heavy metals and sodium from contaminated shrimp ponds Saijai Panwichian1, Duangporn Kantachote*2, Banjong Wittayaweerasak3, Megharaj Mallavarapu4 1Department of Microbiology,
Faculty of Science,
Prince of Songkla University,
Hat Yai 90112, Thailand Financial support: This work was granted by Office of the Higher Education Commission, Saijai Panwichian was supported by CHE PhD. Scholarship. This work was also supported by a project number SCI520001S, Prince of Songkla University. Code Number: ej10035 In order to determine whether waters used for the shrimp cultivation contained toxic levels of heavy metals (HMs) and sodium (Na), analysis was carried out on 31 shrimp ponds in areas of southern Thailand. Purple nonsulfur bacteria (PNB) were also isolated from the same ponds to investigate if they could be used for bioremediation of the above contaminants. The highest HMs concentrations of the sediment samples in mg/kg dry weight were found as follows: 0.75 cadmium (Cd), 62.63 lead (Pb), 34.60 copper (Cu) and 58.50 zinc (Zn). However, all sediment samples met Hong Kong standards for dredged sediment. In contrast, contamination of Cu (9-30 µg/L) and Zn (140-530 µg/L) exceeding the standard guidelines for marine aquatic animal set by the Pollution Control Department, Thailand, were found in 32 and 61% of water samples, respectively. Two metal resistant PNB isolates, NW16 and KMS24, were selected from the 120 PNB isolates obtained. Both isolates reduced the levels of HMs by up to 39% for Pb, 20% for Cu, 7% for Cd, 5% for Zn and 31% for Na from water that contained the highest levels of HMs found and 3% NaCl when cultured with either microaerobic-light or aerobic-dark conditions. The isolate NW16 removed a greater percentage of the HMs than the isolate KMS24, but the isolate KMS24 was able to survive better under a greater variety of environmental conditions. Both strains were therefore suitable to use for further investigating their abilities to remediate water contaminated with HMs and Na. Keywords: bioremediation, heavy metals, purple nonsulfur bacteria, salinity, shrimp farming.
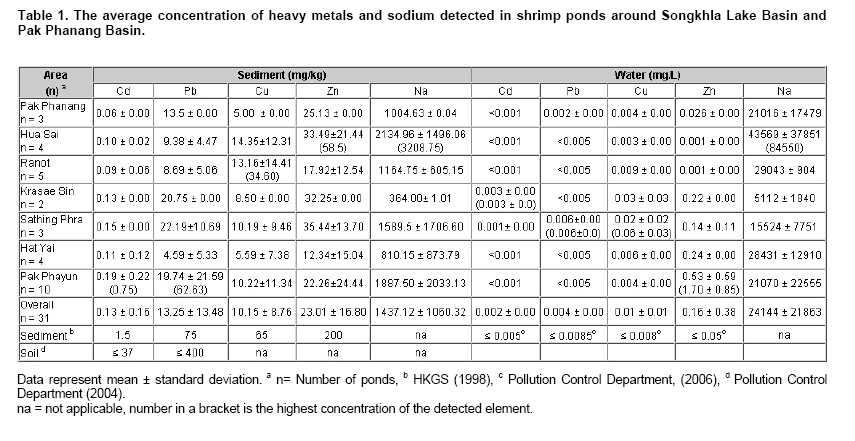
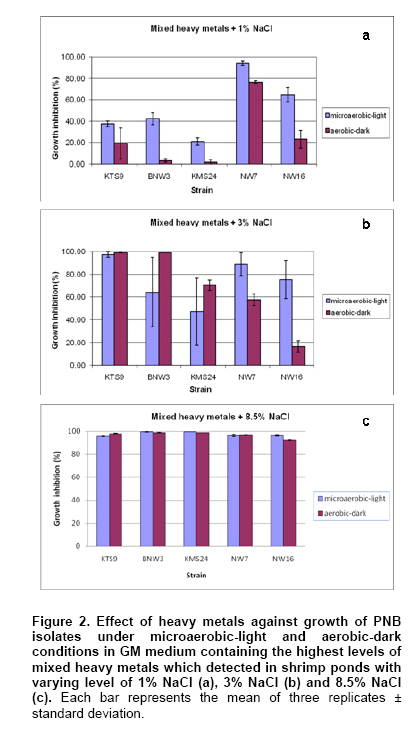
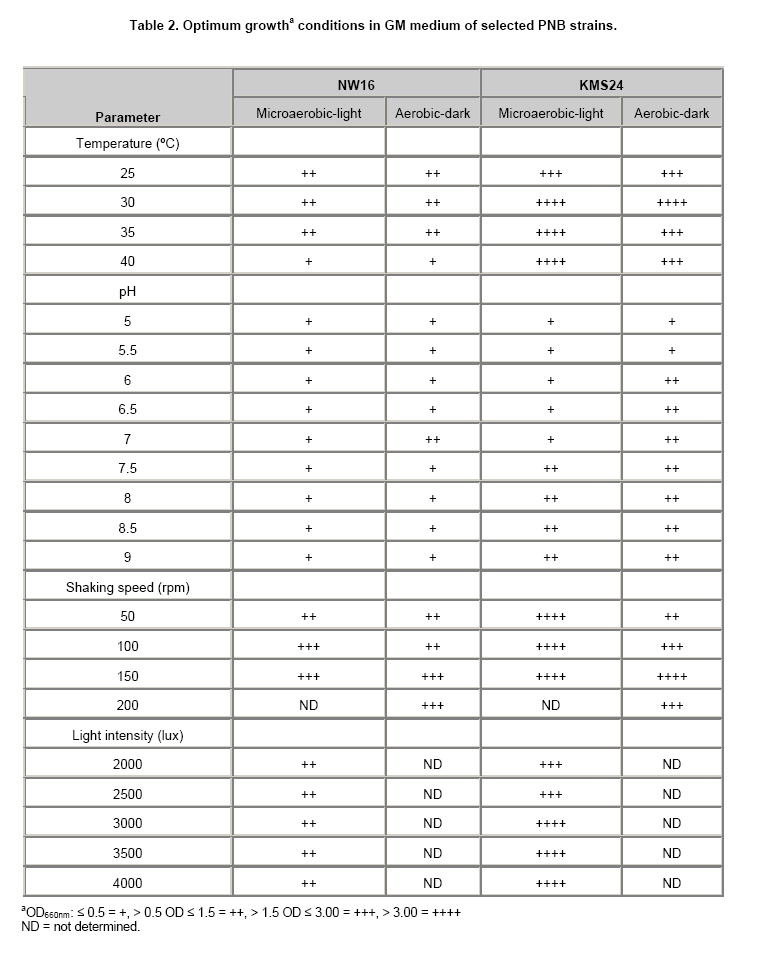
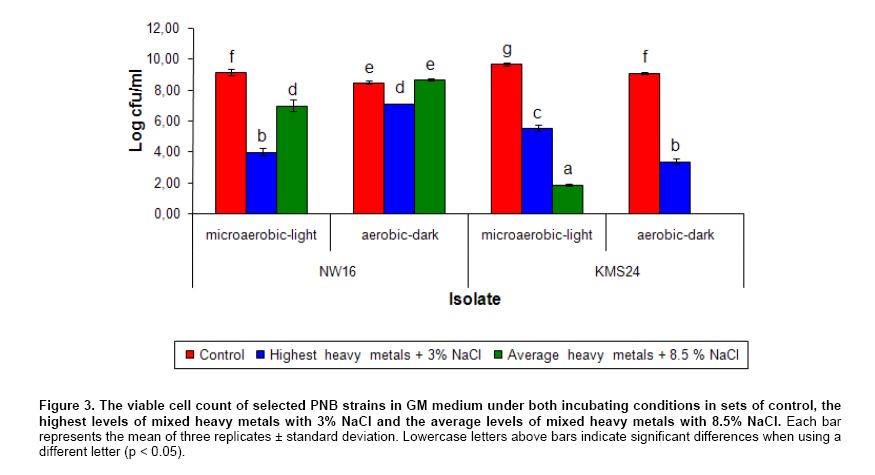
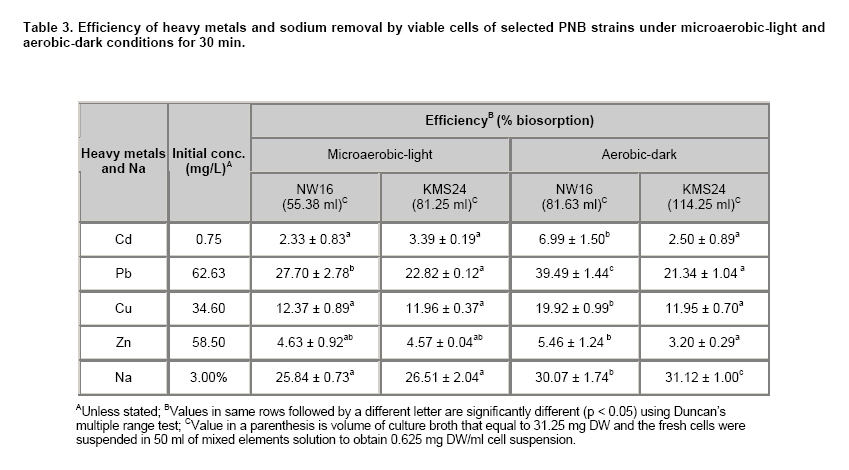
Currently, there have been a number of incidents involving food and food safety such as residuals of antibiotics and heavy metals (HMs) occurring in aquatic animals and their products. Fortunately, the contamination by HMs (i.e. mercury: Hg, Cd, Pb and Zn) in all of the aquatic animal products of Thailand did not exceed standard levels (Petroczi and Naughton, 2009). Among aquatic animals, shrimp is one of the prime exports for Thailand although at present there are many problems with shrimp cultivation and export. The rapid expansion in shrimp cultivation over the last 20 years in Thailand has caused several negative environmental and socio-economic impacts. These include destruction of mangrove forests, salination of the soil and the dispersion of toxic chemicals including HMs into the environment (Flaherty et al. 2000; Kautsky et al. 2000). The Songkhla Lake Basin (SLB) covers some parts of 3 provinces; Nakhon Si Thammarat, Patthalung and Songkhla and is located in southern Thailand along the coastal area of the Gulf of Thailand (Figure 1). SLB is one important area for shrimp farming because this area has many mangroves, so there are few beautiful beaches, and seawater for shrimp cultivation is readily available. Recently, the shrimp business has been on the downturn due to the collapse of the shrimp price and unsuccessful cultivation. There are now many abandoned shrimp ponds in the SLB area, and the soil has been left unfertile and cannot be used to grow other plants due to its increased salinity caused by shrimp farming. However, shrimp farming activities are still being conducted around the SLB. There have been no reports on HMs contamination in the shrimp ponds around the SLB, although some problems have been reported from trace metal contamination i.e. Cd, Cu, Pb, and Zn in the Gulf of Thailand (Cheevaporn and Menasveta, 2003). In addition, the Phawong and U Taphao Canals are major canals that transport wastewater from industries and municipalities to Songkhla Lake (Pradit et al. 2010). Thus, some shrimp farming areas around SLB may be contaminated by HMs from water sources used for shrimp cultivation. The other possible sources of HMs in shrimp ponds are from the use of chemical substances i.e. Cu2+, shrimp food, and perhaps some leaching from fertilizers used in agriculture (Visuthismajarn et al. 2005). Shrimp farming without good management practices can cause contamination by HMs and increased soil salinity in the vicinity of the shrimp cultivation area. Thus, it is important to remove HMs and treat the salinity in the shrimp pond before water is discharged into the environment. The conventional processes used to remove HMs are chemical and physical methods that are expensive and can accumulate toxic substances. Therefore, bioremediation could be an attractive process to remove HMs and to reduce salinity from contaminated areas because it uses organisms like plants or microbes (Vieira and Volesky, 2000; Pietrobelli et al. 2009). Microbes, particularly bacteria, play crucial roles in bioremediation and PNB are an interesting group to use for this purpose as they are normally found in shrimp ponds. PNB have been extensively used for wastewater treatment because they can grow as photoautotrophs/photoorganotrophs under anaerobic/microaerobic-light conditions or as chemoorganotrophs under aerobic-dark conditions (Imhoff and Triiper, 1989; Cheng et al. 2000) and also they are able to utilize a broad range of organic compounds as carbon and energy sources (Kim et al. 2004; Kantachote et al. 2005). Previous studies have shown that PNB have the potential to remove HMs (Watanabe et al. 2003; Giotta et al. 2006; Feng et al. 2007) but there have been no reports that PNB have been isolated from HMs contaminated shrimp ponds. Therefore, the aims of this study were to explore the concentrations of HMs (Cd, Pb, Cu, and Zn) and Na in shrimp ponds and to isolate and screen for PNB from shrimp ponds based on their resistance to HMs and Na, including an ability to remove metal contaminants. Materials and MethodsThe 31 selected shrimp pond sites are located around the SLB and coastal areas along the Gulf of Thailand in the district of Hua Sai, in Nakhon Si Thammarat Province; districts of Ranot, Krasae Sin, Sathing Phra, and Hat Yai, in Songkhla province and the Pak Payun district, in Patthalung province. Other sites, in the district of Pak Phanang, are not part of the SLB as it is part of the Pak Phanang Basin (Figure 1). Collecting and analyzing (HMs and Na) the samples Soil and water samples were collected from 31 shrimp ponds (Table 1). After shrimp harvesting, 13 soil or sediment samples each of about 100 gr were collected from the bottom of each pond at a depth of 5 cm in two diagonal and a half point of each bank. For water samples, 100 ml of water at about 50 cm from the surface water level was collected at or close to the time for shrimp harvesting. However, in some farms, only a few samples of water were collected per pond as there was no boat to support the collecting of samples following two diagonals of the pond. For collection of water samples in 4 shrimp ponds in the Hua Sai district, 2 ponds were storage ponds for retaining the water before discharge into the environment. All composite samples were kept in an ice box and used to isolate PNB as promptly as possible after collection (details will be provided later) and afterwards they were thoroughly combined into a single sample of water or sediment from each pond prior to analysis for HMs (Cd, Pb, Cu, and Zn) and Na using an ICP-OES (PerkinElmer, Germany). Water samples were analyzed directly. Each sediment sample was air dried and passed through a 2 mm sieve. Then 2 gr of sample was digested by nitric acid and hydrogen peroxide method (Radojevic and Bashkin, 1999) before analyzing by ICP-OES. The protocol used for the ICP-OES followed the instruction manual for the instrument. Isolating of PNB from the samples GM broth consisting of 3.8 gr sodium L-glutamic acid, 2.7 gr DL-malic acid, 2.0 gr yeast extract, 0.5 gr KH2PO4, 0.5 gr K2HPO4, 0.8 gr (NH4)2HPO4, 0.2 gr MgSO4.7H2O, 0.053 gr CaCl2.2H2O, 0.001 gr nicotinic acid, 0.001 gr thiamine hydrochloride, 0.01 gr biotin, 0.012 gr MnSO4.5H2O, 0.025 gr ferric citrate, 0.95 gr CoCl2.6H2O, and deionized water up to 1000 ml, pH 6.8 was used to isolate PNB from the sediment and water samples. One ml (1 gr) from each water and sediment sample respectively, was transferred to 5 ml GM broth in a screw cap test tube and then sterile liquid paraffin was added to the top of the growth medium to achieve anaerobic conditions and finally cultures were incubated under continuous light with incandescent lamps (ca. 3000 lux) for 5-7 days. The anaerobic-light conditions were conducted to isolate PNB although aerobic anoxygenic phototrops (AAP) may be present in the collected samples. AAP are obligate aerobes and will not grow with anaerobic conditions in continuous light (Yurkov and Beatty, 1998) and thus the conditions were selective for PNB. Afterwards, the culture broths that were pink, red or brown were streaked onto GM agar and then incubated with the same conditions as previously described to purify the organisms. A pure culture of each isolate was maintained by stab in GM agar and stored at 4ºC until used. Screening for PNB strains resistant to HMs and Na The stock culture was subcultured twice to obtain an active inoculum then one loopful of each culture was transferred to a screw cap test tube containing GM broth leaving a little space to provide microaerobic conditions. All culture tubes were incubated under incandescent lamps (ca. 3,000 lux) for 48 hrs. The OD660 of the culture was adjusted to 0.5 using sterile GM broth as diluent and GM broth as a blank. Four steps were carried out to obtain strains that were resistant to HMs and Na. 1. Primary screening, a 10% inoculum of each culture was added to 10 ml or 18 ml of GM broth in a 15 x 150 mm tube and the cultures were incubated under two growth conditions; aerobic-dark and microaerobic-light for 48 hrs. For consideration of their possible use in shrimp ponds both growth conditions were investigated. All culture tubes were placed in a shaker at 100 rpm at 30ºC, in darkness for 48 hrs for the aerobic-dark conditions while for the microaerobic-light conditions all culture tubes were illuminated at about 3000 lux at the top of the shaker for 48 hrs. Bacterial growth in GM broth was measured at OD660 using a spectrophotometer and any isolates with growth exceeding 0.50 were selected for further screening. 2. In order to screen isolates that were resistant to Na each culture was grown in GM broth plus 1, 3, 5, and 8.5% (w/v) NaCl again with both incubating conditions as described above. 3. Screening for HMs resistant isolates was performed in GM broth containing the highest level of each heavy metal found in the shrimp ponds (Cd 0.75 mg/L, Pb 62.63 mg/L, Cu 34.60 mg/L and Zn 58.50 mg/L). The following inorganic salts; CdCl2, PbCl2, CuCl2 and ZnCl2 were used. Growth in the tubes was measured by the spectrophotometer at OD660 and the percent of growth inhibition calculated by comparing with a control set containing no HMs. 4. Selection and characterization of PNB strains that were resistant to mixed HMs and NaCl in shrimp ponds. Each PNB isolate was grown with GM broth containing the highest concentration of mixed HMs found in the ponds with varying concentrations of NaCl at 1, 3 and 8.5% under both incubating conditions. Determining optimum growth conditions for selected PNB resistant strains Selected strains were used to investigate optimal growth conditions in GM medium under both incubating conditions for 48 hrs. The growth parameters considered for investigations were as follows: pH, temperature and shaking speed while light intensity was only measured under microaerobic-light conditions. Variations of pH: 5, 5.5, 6, 6.5, 7, 7.5, 8, 8.5 and 9; temperature: 25, 30, 35, and 40ºC; and shaking speed: 50, 100, 150, and 200 rpm were tested. Light intensities were varied from 2,000, 3,000, and 4,000 lux and measured using a Denki light meter model DK-211 and the temperature was maintained to 30ºC. Counting of viable cells was conducted under each optimal growth condition based on the above results to obtain more information on the effects of various incubating conditions on the 2 selected strains. Conditions included mixed HMs and NaCl concentrations. Each isolate was grown with either GM broth that contained the highest concentration of each of the mixed HMs with 3% NaCl or in the GM broth containing the average determined concentration of the mixed HMs (Cd 0.15 mg/L, Pb 13.25 mg/L, Cu 10.15 mg/L and Zn 23.01 mg/L) and 8.5% NaCl (the highest level detected in shrimp ponds). The cultures were incubated at 30ºC for 48 hrs under microaerobic-light and aerobic-dark conditions and the growth was then measured using viable plate counts. Plates were incubated under microaerobic-light and aerobic-dark conditions depending on the conditions used for their growth in the broth cultures. For counting growth of PNB under microaerobic-light conditions the plates were placed in a plastic anaerobic jar with Gas-Pak under illumination at 3,000 lux. Estimation of the efficiency of selected PNB strains to remove HMs and Na The two selected PNB isolates were grown in GM broth under optimized conditions according to the aerobic-dark and microaerobic-light conditions for 48 hrs and bacterial cells were separately harvested by centrifugation at 8,000 rpm for 15 min. The cell pellets were washed twice with 0.1% peptone water and fresh cells equivalent to 31.25 mg DW were suspended in 50 ml of mixed HMs at their highest concentrations detected in shrimp ponds and 3% NaCl to obtain 0.625 mg DW/ml. Dry weight was measured following the method in AOAC (2002). The cell suspensions were incubated at 30ºC on a shaking incubator with a speed of 100 rpm for 30 min under aerobic-dark conditions and the same condition was set for the microaerobic-light conditions with the light intensity adjusted to 3,000 lux. After that the bacterial cells were centrifuged at 8,000 rpm for 15 min and each supernatant was analyzed for the remaining HMs concentrations and Na to calculate their disappearance. Data presentation and statistical analysis All experiments in this study were conducted in triplicate unless otherwise stated. Means and standard deviations are presented. Statistical analysis using one way ANOVA to analyze statistical differences at a p-value < 0.05 and mean comparisons were performed by the Duncan’s multiple range test. ResultsContamination of HMs and Na in the shrimp ponds Table 1 shows the levels of HMs and Na in shrimp ponds around Songkhla Lake and the Pak Phanang Estuary. Overall, the average concentrations of HMs; Cd, Pb, Cu, and Zn in the sediment samples (mg/kg dry weight: DW) were 0.13 ± 0.16, 13.25 ± 13.48, 10.15 ± 8.76, and 23.01 ± 16.80, respectively, were much higher than those found in the water samples (mg/L) (Cd: 0.002 ± 0.001, Pb: 0.004 ± 0.003, Cu: 0.011 ± 0.017 and Zn: 0.155 ± 0.378). In contrast, the average Na content in the water samples (24.14 ± 21.86 g/L) was higher than that found in the sediment samples (1.44 ± 1.06 gr/kg DW). In order to identify areas that might have a serious problem with either HMs and/or Na, the highest concentration of each element was considered. Among the sediments, Pak Phayun was the area that had the highest contaminations in mg/kg DW of Cd (0.75), and Pb (62.63) while Hua Sai was highest for Zn (58.5) and Ranot for Cu (34.60) and Na (3.21 gr/kg DW). In contrast, the highest levels (mg/L) of each heavy metal and Na in the water samples were found in different areas such as Pb (0.006) and Cu (0.06) in Sathing Phra, Cd (0.003) in Krasae Sin, Zn (1.70) in Pak Phayun and Na in Hua Sai (84.55 gr/L). Isolation and selection of PNB resistant strains A total of 120 PNB strains were isolated from 31 shrimp ponds, 71 strains (59.2%) from water and 49 strains (40.8%) from sediment samples, but only 100 strains were able to grow well (OD660 > 0.5) over 48 hrs in GM broth under both aerobic-dark and microaerobic-light conditions. Twenty one of these strains (21%) were resistant to 5% NaCl i.e. inhibition < 50%. Most of the strains, that were resistant to 5% salt were inhibited by more than 95% by 8.5% NaCl, only 5 strains survived and some grew well (data not shown). Based on their resistance to Na, 5 strains were classified into being either halotolerant (KTS9, KMS24 and BNW3) or halophilic (NW7 and NW16). Isolates were then further selected for their resistance to the highest levels of each heavy metal (Cd, Pb, Cu, Zn) found in the shrimp ponds. Each strain had a different tolerance to each heavy metal i.e. NW16, BNW3 and KTS9 were the most tolerant to Zn and Cu under aerobic-dark conditions, and Cd and Pb under both incubating conditions. However, strain KMS24 grown under aerobic-dark conditions was resistant to all HMs tested, particularly with Cd and Pb, whereas NW7 was resistant only to Cd and Cu under aerobic-dark conditions (data not shown). As none of the 5 strains stood out as the best possible candidate for use in the shrimp ponds they were all retested for their resistance against a mixture of 4 HMs at the highest levels detected in the sediment samples together with varying concentrations of NaCl under both incubating conditions. Strains; KTS9, BNW3, and KMS24 were resistant to the highest levels of mixed HMs at 1% of NaCl with both incubating conditions (< 50% growth inhibition) (Figure 2a-2c). In contrast, at 1% NaCl strain NW16 was resistant only with aerobic-dark conditions and the growth of strain NW7 was inhibited (> 50% growth inhibition) with both conditions (Figure 2a). When the NaCl content was adjusted to 3%, only two strains (NW16 and KMS24) were resistant to the highest level of mixed HMs under aerobic-dark and microaerobic-light conditions (Figure 2b). Neither of them was resistant with both incubating conditions at the highest levels of mixed HMs at 8.5% NaCl (Figure 2c). Based on the above results, strains NW16 and KMS24 were selected for further investigating their ability to remove HMs and Na. Optimum growth conditions of selected PNB resistant strains The results of changing the following factors on growth, temperature, pH, shaking speed and light intensity affecting the growth of selected PNB strains, NW16 and KMS24, are shown in Table 2. Strain NW16 grew over a range of temperature (25-40ºC) with an optimum temperature between 25-35ºC under both incubating conditions. With microaerobic-light conditions the optimal temperature for strain KMS24 was between 30-40ºC while with aerobic conditions it was 30ºC. Both strains could grow in the full range of pH values tested (5-9). Strain NW16 had a narrow optimum pH of 7 with aerobic-dark conditions whereas strain KMS24 had a wide range of optimum pH from 7.5-9 and 6-9 with both microaerobic-light and aerobic-dark conditions. The shaking speed of 150 rpm was an optimum speed for both PNB strains. For the light factor, an intensity of 3,000-4,000 lux produced optimum growth for both PNB strains under microaerobic-light conditions; however, the strain NW16 grew equally in the range of 2,000-4,000 lux. The effects of the incubating conditions, HMs and Na on bacterial growth (optimal growth conditions) are presented in Figure 3. In the control sets without addition of HMs and Na, the growth of both strains reacted in a similar way with the aerobic-dark conditions being less than the growth with microaerobic-light conditions. In contrast, each strain had a different response to the incubating conditions when the medium contained mixed HMs and Na. Strain NW16 grew less with the GM medium containing both the highest and average levels of HMs plus 3% NaCl or 8.5% NaCl with microaerobic-light conditions than when grown with aerobic-dark conditions. In contrast, strain KMS24 grew more with the microaerobic-light conditions than with aerobic-dark conditions. In addition, the strain NW16 that was identified as a halophilic Rhodobium marinum (data not shown) was more resistant to the highest level of NaCl than with the highest levels of HMs. On the other hand, strain KMS24 that was identified as Rhodobacter sphaeroides (data not shown) was more resistant to the highest levels of HMs than to the highest level of NaCl. Overall, the isolate NW16 showed more tolerance to HMs and NaCl than did the strain KMS24. Efficiency to remove heavy metals and Na of selected PNB strains With optimal growth conditions in GM medium, both strains grew better with microaerobic-light than that with aerobic-dark conditions; strain KMS24 produced a biomass of 0.385 gr DW/L and 0.270 gr DW/L while strain NW16 produced 0.564 gr DW/L and 0.383 gr DW/L, respectively. Hence, results (this experiment and Table 2) indicated that comparisons of growth measurements based on turbidity using a spectrophotometer at the same wavelength (OD660 nm) may not be accurate, especially if the colors of the cells are different like NW16 is pink while KMS24 is brown and the colors also differ with different growth conditions for the same organism. This is why the removal of the elements was tested with the same weight of bacteria rather than the same OD660 nm. The efficiency of the same dry weight of fresh cells (0.625 mg DW/ml) of each strain to remove the highest levels of a mixed HMs and Na in solution is shown in Table 3. Both strains were capable of removing HMs with the relative efficiencies of Pb > Cu > Zn > Cd with both microaerobic-light and aerobic-dark conditions. The highest removal efficiency was found for Pb by the strain NW16 with microaerobic-light and aerobic-dark conditions at 27.70% and 39.49%, respectively. In contrast, the lowest removal efficiency was found for Cd by strains NW16 and KMS24 in a range of 2.33-6.99% with both incubating conditions. The efficiency to remove HMs and Na by strain NW16 in aerobic-dark conditions was more significant than that with microaerobic-light conditions, whereas there was no significant difference found for the strain KMS24. However, the removal percentage of Na by the strain KMS24 was significantly higher with aerobic-dark conditions compared to microaerobic-light conditions. In addition, the strain KMS24 gave a higher efficacy to remove Na (31.12-30.07 = 1.05%) than the strain NW16 with aerobic-dark conditions. The overall results clearly showed that strain NW16 had a higher efficiency to remove HMs than the KMS24 with both incubating conditions but there was no significant difference in ability to remove Na found for either strain with microaerobic-light conditions. As the loss of elements from the solution by fresh cell suspensions with no added substrates is reported as a removal percentage from the solution and the biosorption capacity over the 30 min period could be determined from (biosorption capacity = mg heavy metal removed/gr DW biomass). Then, using data in Table 3 the biosorption capacity for each element can be calculated e.g. the biosorption of Pb by the strain NW16 with aerobic and microaerobic conditions was 39.68 and 27.78 mg/gr DW and equivalent to the removal of 15.20 and 15.67 mg/L culture broth. In the same way, strain KMS24 grown with aerobic and microaerobic conditions remove Pb with a biosorption capacity of 21.38 and 22.88 mg/gr DW equivalent to a loss of 5.77 and 8.81 mg/L culture broth. In case of Na, the biosorption capacity of strain NW16 with light and dark conditions was 12.42 and 14.43 gr/gr DW equivalent to the loss of 7.00 and 5.53 gr/L culture broth. The Na biosorption capacity of strain KMS24 with light and dark conditions was 12.74 and 14.94 gr/gr DW equivalent to a loss of 4.90 and 4.03 gr/L culture broth. This means that the removal of HMs and Na depended on both the biomass capacity and yield of biomass. Based on the above information, it can be concluded that strain NW16 had a higher ability to remove HMs than strain KMS24 with all conditions tested. DiscussionContamination of HMs and Na in the shrimp ponds This study provides information on the distribution of HMs and Na in some shrimp ponds around the SLB and Pak Phanang Estuary in southern Thailand (Table 1). Analysis for HMs showed that sediment samples had significantly higher levels than the water samples. However, based on the standard guidelines issued by the national environmental committee for agricultural soil (Pollution Control Department, 2004) and Hong Kong standards for dredged sediment planning (Hong Kong Government Secretariat, 1998) (Table 1) results of sediment samples showed that the HMs concentrations in shrimp ponds of the study areas were lower than the limitation of HMs allowed in agricultural soil and sediment. In contrast, the distribution of Cu and Zn in water samples from most sampling sites was higher than the standard guidelines for marine aquatic animal cultivation (≤ 8 and ≤ 50 µg/L for Cu and Zn, respectively) (Pollution Control Department, 2006). The Cu contents of 9 and 30 µg/L that exceeded the guideline were detected in 32% of water samples collected from shrimp ponds in Sathing Phra, Krasae Sin and Ranot. Whilst contamination by Zn in water samples (140-530 µg/L) that also exceeded the guidelines was found in 61% of water samples collected from shrimp ponds in Pak Phayun, Hat Yai, Krasae Sin and Sathing Phra. As there was no correlation between the amounts of each heavy metal and Na in the sediment and water samples from each pond (Table 1) this meant that the HMs concentrations in the waters were not caused by the sediments in the shrimp ponds. The results reflect the improper discharge of wastewater from various human activities, e.g. domestic, industry and agriculture in which Cu and Zn were transported into the Songkhla Lake and the Gulf of Thailand as previously mentioned (Cheevaporn and Menasveta, 2003; Pradit et al. 2010) and then this water containing HMs was transported to shrimp ponds for shrimp cultivation. This finding was in agreement with findings of Cheung and Wong (2006) who reported that contamination of the shrimp ponds in the Mai Po Nature Reserve, Hong Kong, was caused by municipal and industrial activities in the surrounding areas (northwest Hong Kong and northern part of Deep Bay). Na levels in the water samples collected from shrimp ponds (Table 1) could be classified into 3 groups as follows: high (4.44%), medium (2.11 - 2.90%) and low (0.51 - 1.55%). In the area of Hua Sai, farmers used sea water from the Gulf of Thailand for shrimp cultivation and in this case a high Na content was found because two storage ponds for waste shrimp water were included in the water samples. In Hat Yai farmers used brackish water from a lower part of the Songkhla Lake which is very close to the Gulf of Thailand (Figure 1) resulting in a medium level of Na. As expected the shrimp ponds in Krasae Sin and Sathing Phra had a low level of Na in water as these areas are located in the upper and middle parts of Songkhla Lake, respectively where the sea water should have little or no effect on the water in this area. Therefore, the amount of Na detected in shrimp ponds of the study areas depended on the salinity of the water sources. From the above information, it seems that contamination by HMs and Na in the water column of shrimp ponds is caused by the sources of water used. In storage ponds there is a high accumulation of Na. Therefore, water after shrimp harvest should be treated before discharge to any environment to prevent HMs and Na into natural areas. If there are no strict regulations by governmental agencies for shrimp farmers to follow the situation might worsen. Properties of PNB isolates resistant to HMs and Na Results indicate that the PNB in shrimp ponds are more frequently isolated from the water (59.2%) rather than from the sediment (40.8%). The dominance of PNB in the water column of the shrimp ponds reflects the situation in that the water is directly exposed to sunlight, while the sediment is rather far from the sunlight or the turbidity in water decreases the light penetration. Among the salt resistant isolates, a halophile group (NW7 and NW16) was isolated from water samples while a halotolerant group (KMS24 and KTS9) was isolated from sediment samples, except BNW3 (Figure 2). It can be explained that isolates were familiar with the amounts of Na in their habitats (Table 1). However, Xu et al. (1998) have reported that PNB isolated from freshwater could adapt themselves to tolerate NaCl in the range of 0%-4%. In contrast, the halophilic PNB are able to grow well in a medium containing NaCl in the range of 1% to 4% while growth is retarded from 5% to 10% (Xu et al. 2001). Each heavy metal and Na had differential toxic effects on the growth of the PNB isolates tested. Additionally, their susceptibility to toxicity of HMs and Na with an aerobic-dark or microaerobic-light conditions differed. It seems that a high amount of Na had a more negative effect on the growth of the 5 PNB isolates than the highest levels of the mixed HMs used (Figure 2). However, with a combination of the highest levels of HMs and Na isolates NW7 and NW16 had retarded growth (> 90% inhibition) even though they are halophilic bacteria (Figure 2c). Based on the results in Figure 2 it is likely that only isolates NW16 and KMS24 could survive in all shrimp ponds after the shrimp were harvested because the pond waters had Na concentrations in a range of 0.51-4.36% (Table 1) and thus their abilities of the 2 isolates to remove HMs and Na were worth investigating. Each isolate had a different response to the incubating conditions when the GM medium was supplement with different concentrations of mixed HMs and Na. The HMs contaminants had a greater negative effect on the isolate NW16 with microaerobic-light conditions while isolate KMS24 was more sensitive with the aerobic-dark conditions. In the control sets with normal media both isolates had the same response to the different growth incubating conditions as both strains grew better with microaerobic-light than that with aerobic-dark conditions. This indicates that each isolate might have different mechanisms to respond to the contaminants. Some can bind the metal ions present in the medium at the cell surface or transport them into the cells and detoxify them in various ways (Ariskina et al. 2004; Gavrilescu, 2004; Mallick, 2004). In these experiments, it is possible that the inhibition observed may be due to the combination of HMs used together with the high Na concentrations. However, both isolates did survive the conditions tested. Therefore, it was of interest to know if these isolates could remove HMs with the conditions that exist in the shrimp ponds i.e. the presence of HMs and 3% NaCl. Efficiency to remove HMs and Na of selected PNB strains In order to investigate the potential of the 2 isolates to remove HMs and Na, their optimal growth conditions were first investigated. Strain KMS24 grew better at a wide range of pH values (6-9) and temperature (30-40ºC) when compared with the strain NW16 (Table 2) but strain NW16 was more resistant to all HMs tested and Na (Figure 3). Based on their optimal growth conditions it was possible to provide one set of conditions for the experiment to determine if suspensions of either strain could remove HMs and Na as follows: pH 7, 30ºC and 150 rpm with a light intensity of 3,000 lux for microaerobic-light conditions. Both selected strains, NW16 and KMS24, with both incubating conditions did remove HMs but with different efficiencies in the order of Pb > Cu > Zn > Cd that were not directly related to their initial concentrations (Table 3). The removal of HMs like Cd, Zn, Cu by halophilic bacteria, PNB and microalgae have been studied and it has been found that the removal efficiency is higher at higher initial concentrations (Al-Momani et al. 2007; Bai et al. 2008; Monteiro et al. 2009). This could be one reason why the removal of Cd was so low as its initial concentration was very low when compared with other HMs (Table 3). However, in case of Zn the removal efficiency was lower than that found in Cu although its initial concentration was higher (Table 3). It may be that Zn gave an adverse effect on live cells of organisms tested and this is supported by Balsalobre et al. (1993) who reported that Zn (10 mg/L) had a negative effect on the growth of R. sphaeroides but Cu at 10 mg/L was not toxic to cells. Hence, it could be explained that less toxicity of Cu compared to Zn caused a higher removal of Cu than Zn by both strains tested. It is possible that the selected isolates may use more than one mechanism to remove metal ions like biosorption (adsorption of metal ions onto the cell surface without requirement of energy) or bioaccumulation (absorption of metal ions into the cells with an energy requirement). However, in order to select for an isolate to use for bioremediation and as the removal efficiency was investigated using living cells suspended in only a mixed HMs solution without addition of nutrients and with a contact time of only 30 min. It is likely that biosorption would be the main mechanism that the biomass could use in the absence of an energy demanding accumulation inside the cells. The big differences between the removal efficiencies of Pb and Cu and Zn might indicate that their mechanisms of removal are different. The results in this study have shown that the removal percentage of each heavy metal is not high. However, the biosorption experiment was performed for only 30 min with a viable biomass equivalent to 0.625 mg DW/ml and it might be possible to obtain a higher percentage of HMs removal by both strains by increasing the exposure time and biomass dose including the use of optimal conditions for pH and temperature. In addition, the organisms may use other mechanisms in addition to biosorption to remove HMs and Na as previously mentioned. It was stated earlier that removal of HMs including Na depended on both the biosorption capacity and yield of biomass and it has been proved that the isolate NW16 gave the best result (Table 3). However, isolate KMS24 seemed to be a good candidate for use in shrimp ponds (Table 2). Therefore, further investigations to achieve the goal of using inoculants to remove contaminants like Cu and Zn (Table 1) from contaminated water will focus on both selected isolates. Findings from this study indicate that the resistant PNB strains, NW16 and KMS24, have the potential to remove HMs and Na in amounts that were found in shrimp ponds with both aerobic-dark and microaerobic-light conditions. Therefore, it will be possible to use both strains as inoculants for bioremediation of water from shrimp ponds contaminated with toxic HMs and Na. AcknowledgmentsWe would like to thank Dr. Brian Hodgson for critical reading the manuscript. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10035t1.jpg] [ej10035f2.jpg] [ej10035f3.jpg] [ej10035t3.jpg] [ej10035t2.jpg] [ej10035f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}