 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
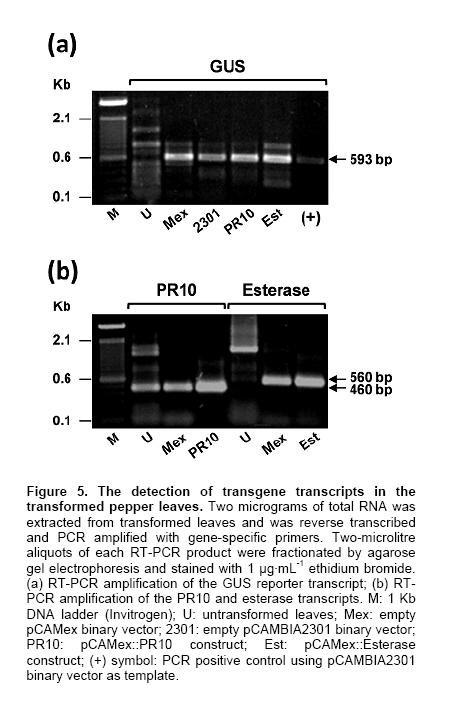
Electronic Journal of Biotechnology, Vol. 13, No. 4, July 15, 2010 Agrobacterium tumefaciens-transient genetic transformation of Habanero pepper (Capsicum chinense Jacq.) leaf explants Guadalupe Fabiola Arcos-Ortega1, Rafael Antonio Chan-Kuuk2, Wilma Aracely González-Kantún3, Ramón Souza-Perera4, Yumi Elena Nakazawa-Ueji5, Elidé Avilés-Berzunza6, Gregorio Godoy-Hernández7, 8Michael A. Lawton, José Juan Zúñiga Aguilar*9 1Centro de Investigaciones
Biológicas del Noroeste
Col. Playa Palo de Santa Rita
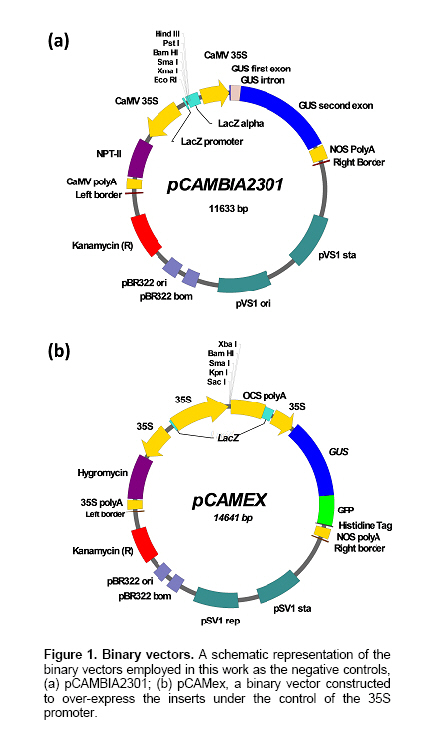
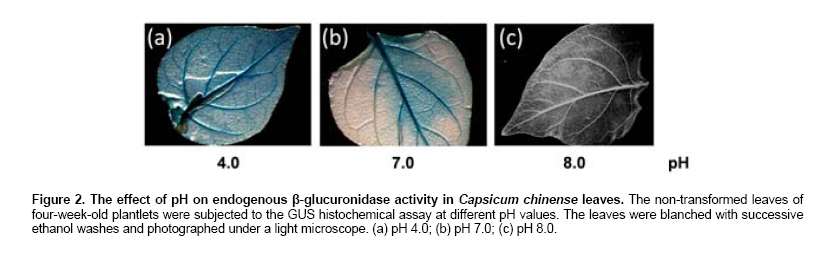
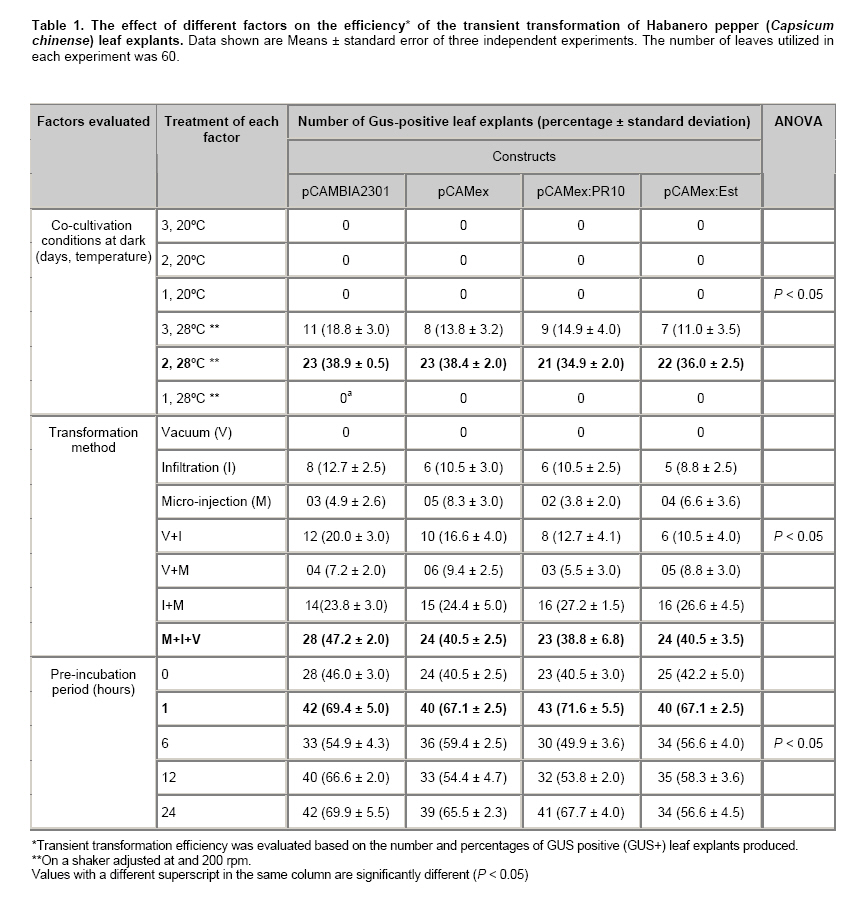
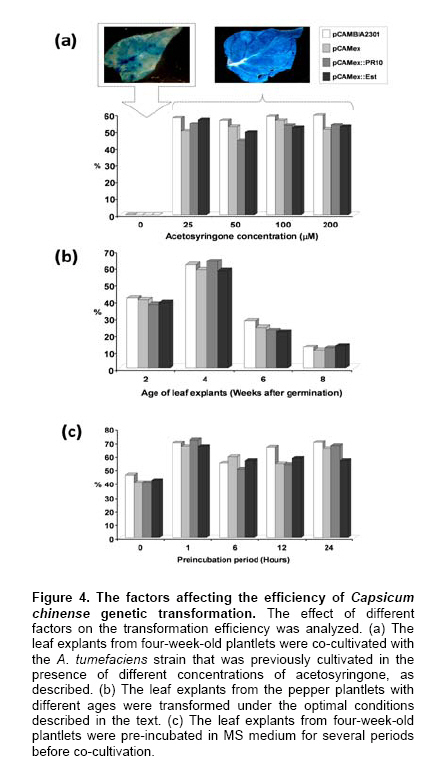
La Paz, B.C.S., México Financial support: This work was funded by the Consejo Nacional de Ciencia y Tecnología (Project P48831). Most of the pepper species of the genus Capsicum have been recalcitrant to efficient Agrobacterium tumefaciens-mediated stable or transient, genetic transformation. In the present work, we optimized a protocol for transient transformation of the Habanero pepper (Capsicum chinense Jacq.) through the standardization of several experimental factors. These included the age of the plants, the temperature, the length of co-cultivation, the application of a negative (vacuum) and/or a positive (infiltration) pressure, along with micro injection, the use of acetosyringone during the bacterial culturing, and modification of the pH during the GUS assay to eliminate the endogenous β-glucuronidase activity. The standardized protocol, which yielded nearly 55% fully transformed leaf explants, was used to successfully mobilize two empty binary vectors (pCAMBIA2301 and pCAMex), as well as the C. chinense cDNAs encoding the pathogenesis-related protein 10 and esterase, respectively. Keywords: cocultivation, GUS assay. In the last decade, Agrobacterium tumefaciens-mediated stable genetic transformation has been successfully performed with several members of the Solanaceae family (Ochoa-Alejo and Ramirez-Malagon, 2001; Nianiou et al. 2002). However, the genus Capsicum has remained largely recalcitrant to stable transformation and as a result only a low frequency and reproducibility (0.5-1% success rate) of transformation has been successfully achieved (Manoharan et al. 1998; Kim et al. 2001b; Li et al. 2003; Lee et al. 2004). The lack of a suitable pepper regeneration system represents a significant obstacle for its genetic manipulation and for the development of functional genomic studies (Ochoa-Alejo and Ramirez-Malagon, 2001; Shivegowda et al. 2002; Li et al. 2003; Lee et al. 2004). To overcome these difficulties, analyses of the gene functions of the pepper have been performed in heterologous systems; in the recent years the heterologous systems of Arabidopsis thaliana (Hong and Hwang, 2006), Solanum lycopersicum (Sarowar et al. 2006), Nicotiana tabacum (Park et al. 2006), and Allium cepa (Hong and Kim, 2005) have been used. However, a homologous transformation system to evaluate gene function is still needed for Capsicum. Transient genetic transformation could be a valuable tool and offer a rapid assessment of gene function prior to stable transformation. By using transient genetic transformation, which is a relatively fast process that would require only 5-8 days from infiltration to gene expression, transformation may occur at high frequencies (Fischer et al. 1999; Van der Hoorn et al. 2000) and as result a large number of constructs may be tested (Kapila et al. 1997). In recent studies, Santana-Buzzy et al. (2005) and Zapata-Castillo et al. (2007) were able to regenerate shoots from the leaf-derived callus of the Habanero pepper. Thus, the transient transformation of Habanero pepper explants might also be exploited to attempt the stable transformation of pepper via organogenesis. In the present study, we assessed several factors to optimize an A. tumefaciens-mediated transient transformation protocol for Habanero pepper leaf explants. The protocol was successfully used to mobilize the full-cDNAs of the genes encoding the pathogenesis-related protein 10 (EMBL AJ879115) and esterase (EMBL AJ879119), which were cloned from a C. chinense cDNA library constructed expressly from Phytophthora capsici-elicited cell suspensions. Materials and MethodsBiological materials and binary vectors Aseptically produced Habanero pepper plantlets of the “naranja”variety (Seminis®) were used as the source of explants. The pepper seeds were surface sterilized by immersion in 70% (v/v) ethanol for 5 min, rinsed twice in sterile distilled water and again in a 30% (v/v) solution of commercial chlorine (Cloralex®, 6% active chlorine) for 15 min with constant stirring. Then the seeds were thoroughly rinsed four times with sterile distilled water, and placed on sterile filter paper to eliminate an excess of water. The seeds were then preconditioned by spending five days at 25 ± 2ºC, in darkness, on Petri dishes of MS medium (Murashige and Skoog, 1962) supplemented with 3% sucrose, 0.2% (v/v) of Gelrite®, without growth regulators and at a pH of 5.6. Finally, the seeds were transferred to a 16 hrs photoperiod (photon flux density of 40-60 µmol·m-2·s-1, provided by 39W fluorescent lamps, Phillips, USA) at 25 ± 2ºC. Upon completion of germination, the plantlets were transferred to magenta containers to wait until they were ready for transformation. Agrobacterium tumefaciens strain LBA4404, either disarmed or transformed with empty or recombinant binary vectors (pCAMBIA2301/pCAMex or pCAMex::PR10/pCAMex::Esterase, respectively) were used for co-cultivation. The pCAMex binary vector was constructed by mobilizing the 35S promoter and the OCS terminator sequences, derived from the pART7 plasmid (Gleave, 1992), into the multiple cloning site of the pCAMBIA1303 binary vector (http://www.cambia.org.au) (Figure 1). pCAMex::PR10 and pCAMex::Esterase were constructed by sub-cloning the corresponding full complementary DNAs encoding the pathogenesis-related 10 (PR10) and esterase proteins, from pTriplEx2 (Clontech, Palo Alto, Cal.), into the pCAMex plasmid digested with Kpn I and Xba I. Wild-type and recombinant A. tumefaciens strains were cultured as described by Zaldivar-Cruz et al. (2003). In short, single colonies from each bacterial strain were inoculated into 50-mL Erlenmeyer flasks containing 25 mL liquid YEB medium with 100 mg·L-1 rifampicin, 100 mg·L-1 streptomycin, and either 50 mg·L-1 kanamycin (sulphate salt, Sigma, St. Louis, Mo.) or, when needed, 50 mg·L-1 hygromycin, all at a pH 5.6. The bacterial cultures were incubated in the presence or absence of acetosyringone (AS, 3’5’-Dimeth-oxy-4’-hydroxyacetophenone) for 24 hrs at 28ºC on a shaker that was adjusted to 200 rpm, until an O.D.600nm of 0.6 (late log phase) was reached. At different periods following germination (2, 4, 6 and 8 weeks), the plantlets were aseptically pre-incubated for different periods of time (0, 1, 2, 6, 12 and 24 hrs) in liquid MS media without growth regulators. Then, the A. tumefaciens strains were infiltrated into the leaves by different methods: (1) The Vacuum method [V]: the leaves were submerged into the bacterial suspensions and a negative pressure (-1 atm.) was applied for 15 min; (2) The Infiltration method [I]: the plantlets and the bacterial suspensions were poured into a 5-mL syringe, then a positive pressure (+1 atm.) was applied for 5 min; (3) The Micro-injection method [M]: the bacterial solutions were infiltrated directly, with a needleless syringe, into different points on the down-side of the leaves; (4) A combination of V and I; (5) A combination of V and M; (6) A combination of I and M; (7) A combination of M and I, then V. Following infiltration, each individual explant was rinsed with sterile MS medium and the co-cultivation was left in the dark for different periods of time (1, 2 and 3 days) at either 20 or 28ºC. After co-cultivation, the leaf explants were washed thoroughly with sterile liquid MS medium and subsequently rinsed four times with 250 mg·L-1 Cefotaxime (Claforan). Sixty leaf explants were used for each treatment. Analysis of transformed plants The efficiency of transient transformation was evaluated by the histochemical β-glucoronidase assay, and then by RT-PCR amplification of the transgene transcripts. The leaf and hypocotyl explants were immersed in X-Gluc solution, consisting of 2 mM X-Gluc (5-bromo-4-chloro-3-indolyl-b-D-glucoronide), 100 mM Tris-HCl pH 8.0, 50 mM NaCl, 2 mM potassium ferricyanide, and 1% (W/V) Triton X-100. After 12 hrs incubation at 37ºC in the dark, the explants were immersed in 70% ethanol until the chlorophyll had been completely bleached. Then, the explants were photographed under a light microscope. According to our requirements, only the explants displaying blue colour in the whole leaf area were considered to be GUS positive. In order to eliminate any possible endogenous GUS activity, the histochemical GUS assay was previously performed in non-transformed leaf explants with X-Gluc solutions and with different pH values (4.0, 7.0 and 8.0) (Alwen et al. 1992). All of the experiments were repeated three times. For the gene expression analysis, total RNA was isolated from the transformed leaves using the Trizol® reagent, according to the manufacturer’s instructions (Invitrogen). RT-PCR was carried out under the standard conditions in a My-Cycler® apparatus (BIO-RAD), with an initial denaturing step at 94ºC for 5 min; cycling conditions were 94ºC for 1 min, 45ºC for 1 min, 72ºC for 1 min; with a final extension of 10 min at 72ºC. The amplified fragments were fractionated by gel electrophoresis on a 1% agarose gel, stained with 1 µg·mL-1 ethidium bromide and visualized under UV light. Amplification of the different transcripts was completed with the following combination of primers. For the GUS transcript, forward (5’-AAAGTGTGGGTCAATAAT-3’) and reverse (5’-AAGCCAGTAAAGTAGAAG-3’) primers amplified a 593 bp fragment. For the PR10 transcript, forward (5’-TAATGAGTCCACAGTCCCAGTTGC-3’) and reverse (5’-CACCATTTAAGCATAGGCAGAAGG-3’) primers amplified a 460 bp fragment. For the esterase transcript, forward (5’-CTTCCTGCAGCTTACGACGATG -3’) and reverse (5’- AGCTGCTGCCTTTGATGGTTC -3’) primers amplified a 560 bp fragment. To estimate the difference between the different treatments, a one-way analysis of variance (ANOVA) was used and the means were compared using the Tukey’s post hoc test. All of the data are reported as the mean ± standard error. For all of the statistical analyses, the percentage results were transformed to arcsine before the ANOVA. The back-transformed means are presented and the standard errors are those obtained from the untransformed variables (Zar, 1999). The statistical significance was preset at P < 0.05, although the P values obtained are indicated. The statistical analyses were done with the General Linear Model (GLM) module with the Statistica software version 7.0 (StatSoft Inc., Tulsa OK, USA). Results and DiscussionIn our lab, we analyze the interaction between Phytophthora capsici and the Habanero pepper at physiological, biochemical and molecular levels. Since one of our research goals is to study the dispersion of the oomycete mycelia through the leaf explants that are transformed with defence genes, we wanted to generate a transformation system that allows homogenous transgene expression through the whole leaf area, instead of having discrete transformation spots. In this study, two types of binary vectors were used, pCAMBIA2301 and pCAMex (Figure 1). The first vector was used as a control to discriminate the false positives produced by the bacterial expression of transgenic GUS activity because this vector contains an intron in the reporter gene that cannot be processed by bacteria. Consequentially, the GUS staining is attributed to be exclusively from transformed plant tissues. pCAMex was constructed from pCAMBIA1303 (http://www.cambia.org) to constitutively express the genes of interest. pCAMex is a versatile binary vector that, in addition to the classic GUS histochemical detection assay, allows monitoring of the transformed tissues in vivo through detection of GFP in a light microscope. Our first efforts to transform the Habanero pepper explants transiently, by vacuum infiltrating the bacterial solutions containing the binary vector controls, yielded no results at all; instead, the leaf explants showed a low level of GUS background activity, even in the negative controls. Since members of the Solanaceae family express an intrinsic β-glucuronidase activity (Plegt and Bino, 1989) that is dependent on the pH (Sudan et al. 2006), we assessed whether the endogenous activity in the Habanero pepper could be eliminated by modifying the pH of the histochemical assay (Alwen et al. 1992). We incubated the non-treated leaf explants with the GUS staining solution, buffered at different pH values. We found that incubation of the leaf explants at pH 4.0 gave a widespread and weak staining that was especially prominent in the vascular system. The staining was reduced at pH 7.0, and was completely eliminated at pH 8.0 (Figure 2). These results confirmed that the Habanero pepper, like other Solanaceae members, possesses an endogenous pH-dependent β-glucuronidase activity. Therefore, the histochemical determinations for all of the transformation assays were done at pH 8.0. Since using a direct application of the vacuum to infiltrate the bacterial solution gave no positive GUS results, we applied either a negative or positive pressure, or a combination of both pressures, in conjunction with a leaf puncture by a syringe needle in order to infiltrate the bacterial solutions. As can be observed in Table 1, a combination of the sequential application of a negative and positive pressure in the punched leaves gave the best results. It has been demonstrated that the age of the plant can be a determinant for a tolerance against pathogenic microorganisms; the older the plant, the more tolerance they exhibit during an interaction with pathogens (Kim et al. 1989). In addition, several factors are critical for a successful transformation and these include the use of young seedling explants (Binns and Thomashow, 1988). Thus, we evaluated whether or not the age of the explants have had any influence over the transformation efficiency. As can be seen in Figure 3a-b and Figure 4a, the maximum efficiency of transient transformation with any of the constructs was reached when 4-week-old plantlets were used as source of explants; with 8-week-old plantlets and older, the transformation efficiency was compromised and serious tissue necrosis occurred in the explants, although non-homogenous transient transformation was still observed (Figure 3a). Acetosyringone (AS), a molecule derived from wounded plants, is used by A. tumefaciens as a chemo-attractant to initiate cell adhesion (Binns and Thomashow, 1988). AS is added during the culturing of bacterial cells to increase virulence and, consequently, the efficiency of gene transfer can be improved (Sheikholeslam and Weeks, 1987; Sunilkumar et al. 1999). In our work, there were no significant differences observed between the different AS concentrations that were evaluated (Figure 4b). However, there was no homogenous transient transformation through the whole leaf area observed when AS was not supplied. This absolute AS-dependency has been also observed in Arabidopsis thaliana (Sheikholeslam and Weeks, 1987) and in C. annuum (Kim et al. 2001b), where AS was one of the essential components for pepper transformation. The co-cultivation was developed for one, two or three days at two temperatures, 20 or 28ºC, and the best results were obtained when the co-cultivation proceeded for two days at 28ºC (Table 1). The transformation efficiency was reduced when the co-cultivation occurred for three days, and no results were obtained when the co-cultivation was executed for one-day at 20ºC. Similar results were reported with C. annuum by Li et al. (2003) who observed that a co-cultivation period of two days was optimal for the transformation of pepper cotyledons and hypocotyls, but a longer (4, 5, 7 days) co-cultivation period resulted in the necrosis of explants, which impeded shoot regeneration. In addition, in Citrus paradisi the highest number of GUS+ shoots and segments with GUS+ shoots were obtained with a two-day co-cultivation period (Costa et al. 2002). In order to control the hypersensitive response of the Habanero pepper leaf explants, induced by co-cultivation with A. tumefaciens, the effect of explant pre-cultivation with MS medium, without growth regulators was tested, using the best conditions from the co-cultivation experiments (2 days in the dark, on a shaker at 200 rpm and at 28ºC). The one-hour pre-cultivation produced an almost 50% increment of change in the frequency of the GUS positive leaf explants for all of the constructs (Table 1). However, an extension of the pre-cultivation period (6, 12 and 24 hrs) did not result in a significant change of the transient transformation (Figure 4c). In the groundnut (Arachis hypogaea), the transformation efficiency of explants was higher when compared to those transformants that were obtained without pre-cultivation (Venkatachalam et al. 1998). Li et al. (2003) believed that the pepper explants, when preconditioned for a relatively long time, could not be infected well by A. tumefaciens, but that the preconditioning promoted the transformation of pepper explants. Over two decades, several studies have presented evidence for the existence of a "window of competence"; a time during which the cells are susceptible to Agrobacterium transformation, although the basis for this promotion of transformation by a pre-culturing of the explants has not yet been completely established. It has been proposed that the plant cells undergo a physiological adaptation to become competent for transformation during the pre-conditioning step (Sunilkumar et al. 1999; Costa et al. 2002). In addition, several authors have suggested that the pre-cultivation step has served to reduce plant-stress (Venkatachalam et al. 1998; Li et al. 2003). The best results were obtained when the different factors evaluated were combined, and this produced an efficient protocol for the transient transformation of the Habanero pepper leaf explants. The most efficient protocol includes a 1 hr pre-incubation in MS medium without growth regulators, followed by a combination of the three transformation methods previously evaluated, addition of 100 µM AS to the YEB medium during bacterial culturing, and co-cultivation for 2 days in MS sterile liquid medium without growth regulators, in the dark and on a shaker adjusted to 200 rpm at 28ºC. We evaluated the expression of transgenes in the leaf explants that were transformed under these conditions. By means of RT-PCR assays, high levels of GUS transcripts were detected in the leaves transformed either with empty vectors or with PR10 and esterase genes (Figure 5a); as expected, GUS transcripts were not detected in the untransformed leaves (Figure 5a, lane 1). The transcripts for the PR10 and esterase genes were also detected in the transformed leaves (Figure 5b); however, both of the genes were also expressed, at lower levels, in the leaves transformed with the empty vectors (Figure 5b, lanes 3 and 5). Furthermore, lower levels of PR10 were present in the untransformed controls (Figure 5b, lane 2). As a member of the pathogenesis-related proteins, PR10 may be expressed as a result of both biotic and abiotic stresses. Esterase is not classified as a PR-family member, but it is a hydrolase that functions in the defence against pathogens (Kim et al. 2001a). Thus, it is possible that these particular genes are expressed as a result of the transformation process itself, and that they are not the best examples to evaluate when determining the efficiency of genetic transformation. Nevertheless, as the results of GUS gene expression are clear, it is perfectly possible that the transformation procedure resulted in the over-expression of PR10 and esterase genes, in addition to their basal expression. Over the past five years, remarkable progress has been made in the development and understanding of the A. tumefaciens infection and transformation processes. The literature reviewed has provided strong evidence that there is a high recalcitrance of the peppers to the Agrobacterium-mediated transformation, and to cell culture in vitro. As a consequence, there are no standard protocols that can be used between the different species, even within the Genus Capsicum. Researchers around the world use heterologous systems to express the pepper genes in order to study their biological functions (Kim et al. 2004). In the present study, a good system for the transient transformation of Capsicum chinense, the hottest habanero pepper worldwide (Canto-Flick et al. 2008), has been established. This optimized protocol can thus be used to study pepper gene functions in a homologous system. AcknowledgmentsGFAO and YENU gratefully acknowledge Consejo Nacional de Ciencia y Tecnología for the scholarships 56153 and 172927, respectively.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10039f4.jpg] [ej10039f2.jpg] [ej10039f1.jpg] [ej10039f5.jpg] [ej10039t1.jpg] [ej10039f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}