 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
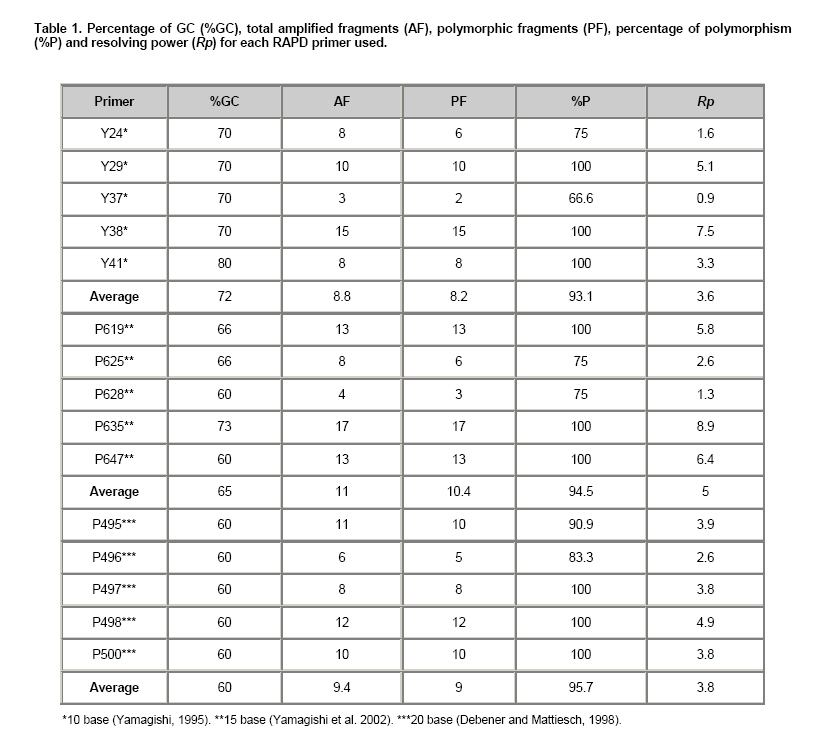
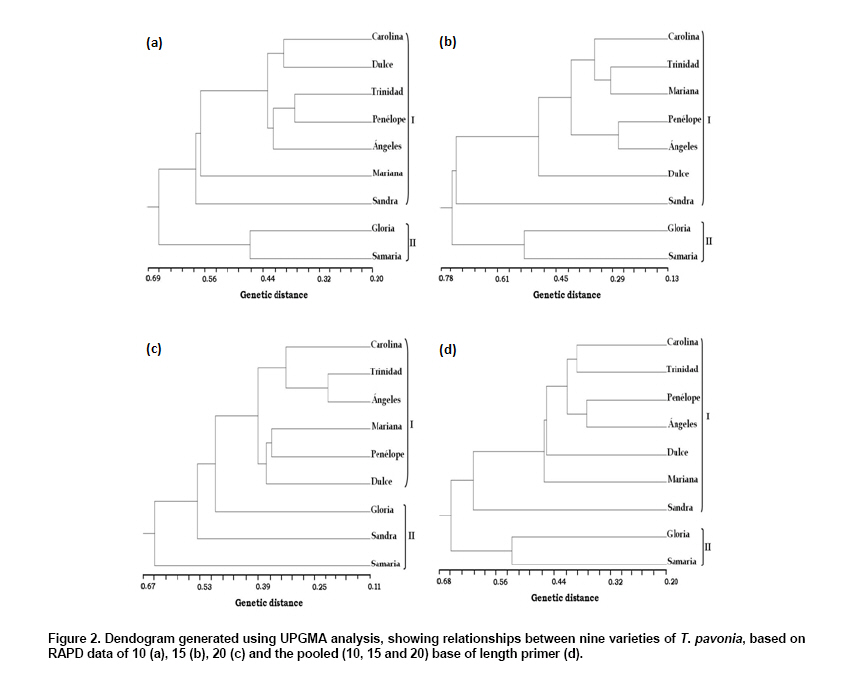
Electronic Journal of Biotechnology, Vol. 13, No. 4, July 15, 2010 Variety discrimination of Tigridia pavonia (L.f.) DC. assesed by different length RAPD primers José Luis Piña-Escutia1, Luis Miguel Vázquez-García2, Amaury Martín Arzate-Fernández*3 1, 2Centro de
Investigación y Estudios Avanzados en Fitomejoramiento
Facultad de Ciencias Agrícolas
Universidad Autónoma del Estado de México
Carretera Toluca-Ixtlahuaca km 11.5
Campus Universitario “El Cerrillo”50200
Toluca, Estado de México, México Financial support: We are grateful to Consejo Nacional de Ciencia y Tecnología (CONACYT) from México for the scholarship give to JLPE (2008-213641), and to the RED TIGRIDIA (SINAREFI, México) for covering the publishing payment. Code Number: ej10042 Tigridia pavonia (L.f.) DC. is one of the important phytogenetic resources of México. This species is used as ornamental, food and medicinal purposes. Despite its ornamental and economic potential, there is little information about the genetic variability. In this study, randomly amplified polymorphic DNA (RAPD) primers of 10, 15 and 20 bases were used to assess the level of genetic variation among nine botanical varieties of Tigridia pavonia collected in three localities within State of México. The total number fragments, polymorphic fragments, percentage of polymorphism and resolving power were greater for 15 base (55, 52, 94.5 and 5, respectively) and 20 base (47, 45, 95.7 and 3.8, respectively), in comparison with those obtained from 10 base primers (44, 41, 93.1 and 3.6, respectively). Results showed the major effectiveness of 15 and 20 bases RAPD primers in the genetic differentiation of varieties as compared to 10 bases RAPD primers. The dendrograms based on un-weighted pair group method arithmetic average (UPGMA) analysis of the 10, 15, 20 and the pooled (10, 15 and 20) bases RAPD data were consistent in the clustering varieties, grouping them in two main clusters. Keywords: genetic resources, RAPD, Tigridia pavonia, variety discrimination.
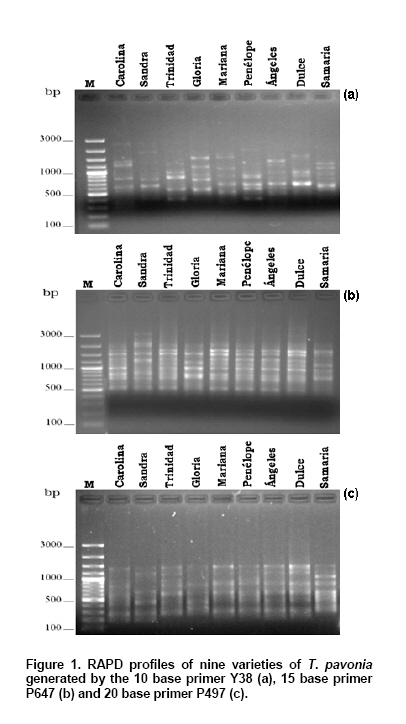
Tigridia pavonia (L.f.) DC., also known as “tiger flower or oceloxóchitl”, is a native bulbous plant of Mexico that grows as wild plant. Due to flower beauty, this species is been used as ornamental plant (Vázquez-García et al. 2001a). Furthermore, the bulb is used for food (cooked bulbs) (Martínez et al. 2007) and medicinal purposes (as antipyretic) (González et al. 2004). Although in México there is no record of its commercial production, nowadays it is been widespread in Europe, Asia and Australia where is commercialized as garden plant. Thus, the extraction of wild material and the lack of a rational management of this species could cause a genetic erosion of the same (Vázquez-García et al. 2001b). The recognition of genetic diversity of these Mexican native species is fundamental for the conservation, evaluation and suitable selection of each genotype, which are the base in the development of plant breeding programs. Genetic diversity has been traditionally performed by morphological markers. Thus, Vázquez-García et al. (2001a) described morphologically nine varieties of T. pavonia. However, these markers have been shown unreliable due to the influence of environmental conditions. Besides these, morphological characters are possible to evaluate only at level of the whole plant or in mature stage (Vicente and Fulton, 2003). Isozymes analysis showed low variation in banding patterns among nine botanical varieties of T. pavonia (Arzate-Fernández et al. 2008), which indicate the limited utility of such biochemical markers for variety identification purposes, because they could be also limited by the environment influence and for the changes that happen in different stages of plant development (Vicente and Fulton, 2003). Molecular markers have been used recently in the varieties identification and genotype characterization because they offer a fast screening, a more precise discriminatory power, and are independent of the environmental factors (Vicente and Fulton, 2003). RAPD markers have been most extensively used due to the simplicity, short time requirements, and no information of DNA sequences. These markers have been also applied successfully in the genetic variation analysis of closely genotypes, in a wide range of species such Chrysanthemum mutants (Kumar et al. 2006), Opuntia spp. (Luna-Paez et al. 2007), Phaseolus vulgaris (Marotti et al. 2007), Vigna umbellate (Muthusamy et al. 2008), Zinnia elegans (Ye et al. 2008), and others. Generally decamers are being used as RAPD markers for the genetic diversity studies of plant species. Nevertheless, the use of longer primers (15-20 bases) has been also evaluated for the same purpose (Debener and Mattiesch, 1998; Yamagishi et al. 2002; Solouki et al. 2007). Thus, it has been observed that depending of the primer length, the DNA fingerprint profiles can be affected. While in some species the use of 10 base primers produce more polymorphic bands and a best genetic differentiation (Debener and Mattiesch, 1998; Kwak et al. 2009), in others, the major efficiency to produce polymorphic markers can be obtained with the use of 15 or 20 base primers (Yamagishi et al. 2002; Solouki et al. 2007; Solouki et al. 2008). Considering the mentioned above, the utility of different length base primers could be also applied to assessment of genetic relationships in species for which there is no sequence information available. For T. pavonia, reports about molecular techniques are very limited. Thus, the objectives of this study were evaluate the efficiency of 10, 15 and 20 base primers in the generation of RAPD polymorphisms, and to determine the genetic differentiation of nine varieties of T. pavonia. Materials and MethodsNine botanical varieties of Tigridia pavonia (L.f.) DC were collected in three localities within of the State of Mexico: Tenancingo (Sandra), Temascaltepec (Carolina, Trinidad, Mariana, Penelope, Ángeles and Dulce), and Temoaya (Gloria and Samaria). These varieties were grown at the same time, in a greenhouse at the Facultad de Ciencias Agrícolas (FCA) of the Universidad Autónoma del Estado de México (UAEM). Young leaves were collected of two individuals (in the same development stage), of each T. pavonia variety and stored at -20ºC prior to DNA extraction. All of the samples were processed in the laboratory of plant molecular biology at the FCA of the UAEM. The genomic DNA was extracted of approximately 100 mg of leaf tissue of each variety of T. pavonia. The extraction procedure was the CTAB as reported by Zhou et al. (1999). The DNA samples were stored to -20°C prior to RAPD analysis. All DNA amplifications were performed in a total reaction volume of 10 µl containing 10 ng DNA template, 1X PCR buffer, 1.25 mM MgCl2, 250 µM of each dNTPs, 0.2 µM primer and 0.1 units Taq DNA polymerase (Mercury, California, USA). For RAPD reactions, five 10 base primers (Yamagishi, 1995), five 15 base primers (Yamagishi et al. 2002), and five 20 base primers (Debener and Mattiesch, 1998) were used (Table 1). Amplifications were performed in a thermocycler (Mastercycler gradient, Eppendorf, Germany). The RAPD program for 10 base primers was the reported by Yamagishi (1995). The program for 15 base primers used in this study was followed according to Yamagishi et al. (2002) with minor modifications (40 cycles of 94ºC for 1 min, 53ºC for 3 min and 72ºC for 2 min). The PCR cycle conditions for 20 base primers was performed according to Debener and Mattiesch (1998). The PCR amplified products were subject to horizontal electrophoresis using 1% agarose in 1X TAE buffer at 100 V and 120 mA for 80 min. 100 base pair DNA ladder (MBI Fermentas) was used as molecular weight marker, and the gels stained with ethidium bromide were visualized using a Transilluminator (UVP, USA). The RAPD bands were scored either absent (0) or present (1). For considering a marker as polymorphic, the absence of an amplified product in at least one variety was used as criteria. The total number fragments (FT), polymorphic fragments (FP), percentage of polymorphism (%P) and the Nei ̓s genetic distance (GD) were calculated using POPGENE version 1.32 software (Yeh and Boyle, 1999). The cluster analysis was implemented on the basis of the genetic distances using the UPGMA (un-weighted pair group method arithmetic average) method. The capacity of each RAPD primer to distinguish the varieties was studied by calculating resolving power (Rp), which is based on the distribution of alleles within the sampled genotypes, and is defined as: Rp = Σ Ib where Ib is the informative band, that takes the values of: 1-(2 x [0.5-p]), being p the proportion of the nine varieties of T. pavonia containing the band (Prevost and Wilkinson, 1999). A simple correlation analysis was performed to investigate the correlation between the length primer and the polymorphism generated by each primer group. Results and DiscussionThe varieties discrimination through DNA profiles is very useful for the protection and conservation of the genetic resources. In this respect, RAPD markers have been most extensively used due to the simplicity, short time requirements, and low technical requirements of DNA purity. These markers also play an important role in assessing the genetic diversity where no sequence information is available. In the present study, PCR amplification with 10, 15 and 20 length base RAPD primers led to reproducible fragment patterns for all varieties of T. pavonia evaluated. The majority of those RAPD fragments ranged from 250 to 2800 bp (Figure 1). For 10 base primers, the average of total and the polymorphic fragments generated per primer were 8.8 and 8.2, respectively, whereas for 15 base primers, those values were 11 and 10.4, respectively, and for 20 base primers, 9.4 and 9, respectively (Table 1). Generally, in RAPD analysis 10 base primers are preferred, nevertheless, because of the high annealing temperature applied in PCR reaction than their original, some 10 base primers could not hybridize with template DNA, generating only a few minor bands or none (Yamagishi et al. 2002), while the advantage of long primers is the smaller number of fragments containing repetitive DNA, thereby increasing the specificity and reproducibility of DNA fragments (Debener and Mattiesch, 1998). In the present study, the positive correlation (r = 0.99; p ≤ 0.05) between primer length and the percentage of polymorphism generated per each primer was observed, indicating that the efficiency of primers to generate polymorphic fragments it increased by primer length. Thus, 15 or 20 base primers generated more DNA fragments (55), and a greater number of polymorphic fragments (52), in comparison with those obtained by the 10 base primers (44, and 41, respectively) (Table 1). The high efficiency of long primers to generate a large number of RAPD markers has been also reported in other species as Lilium spp. (Yamagishi et al. 2002), Vitis vinifera (Solouki et al. 2007) and Matricaria chamomilla (Solouki et al. 2008). Although it is no clear exactly why the long primers produced more polymorphic bands, it has been reported that the GC content may be a factor that determine the efficiency of a primer (Solouki et al. 2007), since GC content is associated with annealing temperature and related to generation of more DNA fragments. Thus, it has been observed that with long primers with a lower GC content, the efficiency for amplifying polymorphic bands is higher, in comparison with short primers with a higher GC content (Ye et al. 1996; Solouki et al. 2007). According to this, it is possible that the GC content has favored the major efficiency of the 15 (65% GC) and 20 (60% GC) base primers used in the present study, because more DNA fragments were amplified and also a greater percentage of polymorphism, in comparison with those amplified with 10 base primers with 72% GC content (Table 1). Our results are closely similar with those reported by Solouki et al. (2008) where it were obtained more DNA fragments and 100% of polymorphism, using a long primer with low GC content. The high efficiency of long primers in the genetic differentiation of T. pavonia varieties was also confirmed with the measurement of resolving power. The higher values of Rp belonged to the 15 and 20 base primers (5 and 3.8, respectively) (Table 1), indicating a better distinction of the varieties. So, these results also confirm the utility of the Rp as measure of capacity of a primer to discriminate among closely related individuals as was pointed out by Prevost and Wilkinson (1999) and Escandón et al. (2007). The dendrograms based on UPGMA analysis of the 10, 15, 20 and the pooled (10, 15 and 20) RAPD data showed the genetic differentiation of the nine varieties of T. pavonia (Figure 2). In the dendrogram generated with decamers, the GD among the varieties ranged from 0.20 to 0.69, with an average of 0.24. For the 15 base dendrogram, the GD ranged from 0.13 to 0.78, with an average of 0.32, while the dendrogram of the 20 base showed a range of GD of 0.11 to 0.67, with an average of 0.28, and finally the dendrogram with pooled data showed a range of GD of 0.20 to 0.68, with an average of 0.24. All four dendrograms clearly grouped the varieties in two major clusters. In the dendrograms of 10 and 15 bases, and in the pooled, the cluster I consisted of the varieties Carolina, Dulce, Trinidad, Penélope, Ángeles, Mariana, and Sandra, whereas the cluster II grouped the varieties Gloria and Samaria. The close relationship observed in these dendrograms, among the varieties collected in Tenancingo and Temascaltepec, might be the result of having some common morphological characters such as: number of shoots by bulb, number of nodes by shoot, length of leaf, and number flowers by shoot. The separation of Gloria and Samaria collected in Temoaya, could be due to the highest values for length of shoot, length of internode, and length of floral escape, besides that the flowering of these varieties is delayed in comparison with the rest of them, as it was reported by Piña-Escutia et al. (2010). On the other hand, the dendrogram developed with 20 base primers also grouped the cluster I similar to the dendrograms generated with other primers sets, except for the variety Sandra, which was grouped with the varieties Gloria and Samaria in the cluster II. Thus, this dendrogram showed a best differentiation of the varieties evaluated, confirming the utility of long primers in generation high polymorphism and genetic discrimination of plant species, which was also reported by Solouki et al. (2007), and Solouki et al. (2008). In a previous study, a morphological identification was applied to nine varieties of T. pavonia (Vázquez-García et al. 2001a), but this method could not be reliable enough due to the environment effect on phenotype. Likewise, a biochemical characterization of the same varieties using isozymes was performed (Arzate-Fernández et al. 2008). However, that study revealed that the varieties Ángeles and Sandra as well as Carolina and Dulce were the same. In contrast, the present study differentiated all varieties with the use of 10, 15 or 20 base length RAPD markers. The low discrimination power of isozymes can be due to inadequate genome coverage. Besides that, those markers can be influenced by environmental factors, and the level of expression differences cause confusion in the interpretation of the results, whereas the RAPD markers offer a fast screening, a more precise discriminatory power, and are independent of the environmental factors (Vicente and Fulton, 2003). The assesment of genetic variability of Mexican native species is fundamental for the conservation, and plant variety protection. In this sense, the results presented in this paper are very significant because it first time revealed the use of RAPD markers in the genetic differentiation of varieties of T. pavonia which eventually can be utilized in the development of plant breeding programs of this specie. Concluding RemarksThe RAPD analysis showed the effectiveness of 10, 15 and 20 base RAPD primers in the detection of polymorphism and genetic differentiation of nine varieties of T. pavonia.However, the efficiency of primers to generate polymorphic fragments increased with increasing primer length. The study revealed that 15 and 20 base random primers are more efficient to detect polymorphism and genetic differentiation of the T. pavonia varieties. AcknowledgmentsWe also thank to Mrs. Ana Isabel Quintana Carapia, for her logistic support. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10042t1.jpg] [ej10042f2.jpg] [ej10042f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}