 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 4, July 15, 2010 Mapping aluminum tolerance loci in cereals: A tool available for crop breeding Claudio Inostroza-Blancheteau1, Braulio Soto2 , Cristian Ibáñez#3, Pilar Ulloa4, Felipe Aquea5, Patricio Arce-Johnson6, Marjorie Reyes-Díaz*7
1Programa de Doctorado
en Ciencias de Recursos Naturales,
Universidad de La Frontera,
Temuco, Chile Financial support: Fondecyt Project Nº11080231, C. Inostroza-Blancheteau was supported by a PhD fellowship from CONICYT-Chile and F. Aquea is supported by a Postdoctoral Project “Programa Bicentenario de Ciencia y Tecnología CONICYT Banco Mundial”PSD74 2006 and the Millennium Nucleus for Plant Functional Genomics (P06-009-F). Code Number: ej10043 Aluminum (Al) toxicity is the main factor limiting crop productivity in acidic soils around the world. In cereals, this problem reduces crop yields by 30-40%. The use of DNA-based markers linked to phenotypic traits is an interesting alternative approach. Strategies such as molecular marker-assisted selection (MAS) in conjunction with bioinformatics-based tools such as graphical genotypes (GGT) have been important for confirming introgression of genes or genomic regions in cereals but also to reduce the time and cost of identifying them through genetic selection. These biotechnologies also make it possible to identify target genes or quantitative trait loci (QTL) that can be potentially used in similar crops to increase their productivity. This review presents the main advances in the genetic improvement of cereals for Al-tolerance. Keywords: ALMT1, aluminum tolerance, cereals, marker-assisted selection, organic acid.
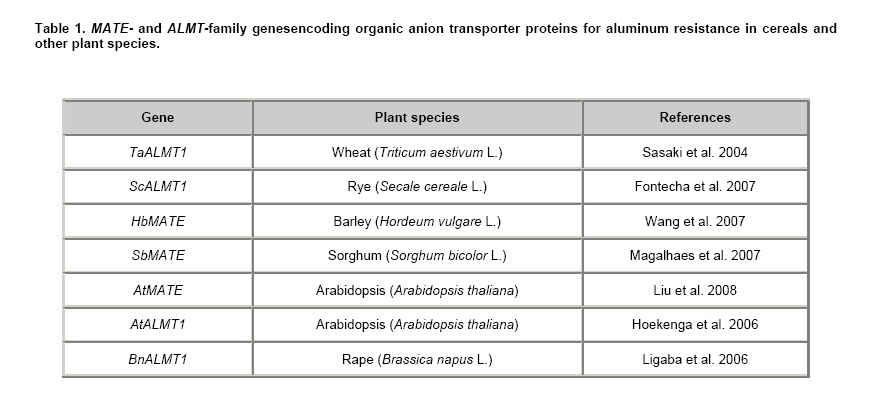
Aluminum (Al) is a light metal that makes up 7% of the earth’s crust, and is the third most abundant element after oxygen and silicon (Ma et al. 2001). Al is also one of the major factors that limits the productivity of acidic soils (Kochian, 1995) affecting almost 40% of soils used for agriculture around the world (Foy et al. 1978; Zheng et al. 1998; Delhaize et al. 2004). Most of the Al can be found in alumino silicates (Al2O3, SiO2) with only small amounts present in soluble forms in the rhizosphere. Acid soils (pH < 5) increase the phytotoxic levels of trivalent Al species (Al3+) whereas at higher pH other non-toxic forms such as Al(OH)2+ and Al(OH)2+ are more prevalent (Delhaize and Ryan, 1995). Al toxicity primarily affects the division and elongation of the root apex. When Al3+ penetrates into the roots, it binds to the negative charges of phospholipids in the plasma membrane leading to rigidification and disruption of membrane function and also enhancement of oxidative stress (Jones et al. 2006). These physiological changes in the root cell result in the poor uptake of nutrients and water that ultimately affect crop yields (Kochian, 1995; Ciamporová, 2002; Inostroza-Blancheteau et al. 2008). In response to Al stress, higher plants have evolved two main mechanisms to resist the effect of Al toxicity. The first is an exclusion mechanism in which Al is prevented from moving through the plasma membrane to the cytoplasm in the root cells (Kochian, 1995; Giaveno and Miranda, 2000). This is achieved by the secretion of organic acids from the radical apex to the rhizosphere which, in turn, modifies the pH and chelates the toxic Al3+ (Marschner, 1986; Miyasaka et al. 1989; Taylor, 1995; Degenhardt et al. 1998; Kinraide et al. 2005). The second mechanism involves chelation of Al by specific proteins, short-chain organic acids, phenolic compounds and tannins that can bind and form complexes with Al3+and subsequently compartmentalize it in the vacuole thus reducing Al-toxicity in the cell (Basu et al. 1994b; Jones, 1998; Jones and Ryan, 2004). In a species-specific manner, several studies have shown that there are differences in the type of organic acid involved in this Al3+ detoxification. For example, in Triticum aestivum (wheat), the organic acid involved in the Al exclusion mechanism is malate (Delhaize et al. 1993; Basu et al. 1994a; Ryan et al. 1995), in Fagopyrum esculentum (buckwheat) and Colocasia esculenta (taro) the organic acid uses is oxalate (Ma and Miyasaka, 1998, Zheng et al. 1998) whereas in Phaseolus vulgaris (snapbeans), Cassia tora (tora) and Zea mays (maize), citrate is employed (Miyasaka et al. 1991; Pellet et al. 1995; Ma et al. 1997; Piñeros et al. 2002). In Secale cereale (rye), exudation of both malate and citrate have been reported (Li et al. 2000). Citrate, malate and oxalate are organic acids with high affinity for Al3+ (Jones, 1998). Transport of these organic acids occurs via anionic channels, the opening of which may be activated by Al. Differences regarding the activation of these channels have been observed between tolerant and sensitive genotypes (Ryan et al. 1997; Zhang et al. 2001). The identification of a possible protein responsible for the transport of organic acids in wheat may indicate the existence of a new type of membrane transporter (Sasaki et al. 2004). This gene is called ALMT1 (Aluminum Activated Malate Transporter 1) and when expressed in transgenic rice seedlings and sensitive barley plants, ALMT1 activated malate exudation in roots (Delhaize et al. 2004). Recent studies in barley have identified another gene, HvAACT1 (Hordeum vulgare Aluminum Activated Citrate Transporter 1), which belongs to the multidrug and toxic compound extrusion family (MATE) and is responsible for citrate exudation in response to Al (Furukawa et al. 2007). Additionally, in Sorghum bicolor (sorghum) a MATE gene (SbMATE) was identified as an aluminum-activated citrate transporter (Magalhaes et al. 2007). The monogenic inheritance of genes encoding proteins responsible for transporting organic acids in cereals such as T. aestivum and Hordeum vulgare (barley) facilitates the prospects of improving these species for tolerance to Al in acidic soil. Knowledge of the molecular physiology of Al-tolerance and the genetics that control this trait, may allow significant advances in the development of tolerant varieties in sensitive cereals. Nevertheless, the complexity of this genetic control seems to vary among species. For instance, control of Al tolerance in Oryza sativa (rice) is polygenetic, thus making genetic improvement difficult for this trait (Nguyen et al. 2003). Although conventional breeding methods have been useful in identifying tolerant varieties of various crops (Riede and Anderson, 1996; Gallego and Benito, 1997; Tang et al. 2000), they do not guarantee per se an efficient transfer of these genes to other elite materials. Fortunately, it is possible to increase the efficiency of conventional breeding by combining it with marker-assisted selection (MAS) and gene mapping strategies, which reduces the costs and the selection time of developing Al-resistant varieties. Genetics of Al3+ tolerance in cerealsThe genetic control of Al tolerance has only been studied for a limited number of species of agronomic interest. In cereals such as T. aestivum, H. vulgare, S. cereale, S. bicolor and Avena sativa (oat), different members of the ALMT and MATE families have been found to control Al tolerance traits (Gallego and Benito, 1998; Raman et al. 2002; Miftahudin et al. 2002; Tang et al. 2002; Magalhaes et al. 2004; Nava et al. 2006). In T. aestivum, the major locus that defines Al tolerance has been mapped on the long arm of chromosome 4D (4DL), and has been mapped in many different segregating populations and is linked to known molecular markers (Luo and Derorak, 1996; Riede and Anderson, 1996; Rodríguez-Milla and Gustafson, 2001; Raman et al. 2005; Raman et al. 2008). Recently Ryan et al. (2009) identified a major locus on chromosome 4BL that accounts for 50% of the phenotypic variation in citrate efflux. This suggests that citrate could be acting as a second mechanism for Al resistance in T. aestivum (in conjunction with malate). This locus, Xcec, was mapped within 6.3 cM of the Simple Sequence Repeats (SSR) marker Xgwm495. Nevertheless, studies in the Atlas 66 cultivar of T. aestivum determined that not all genes were located on the 4DL chromosome suggesting that Al-tolerance is inherited polygenically in this cultivar (Berzonsky, 1992). Zhou et al. (2007) identified a minor quantitative trait loci (QTL) for Al tolerance in Atlas 66 on chromosome 3BL in a population of recombinant inbred lines derived from the Chisholm cultivar of T. aestivum. Raman et al. (2005) identified a major QTL on chromosome 4DL that accounts for approximately 55% of the genetic variation for Al-resistance (chromosome 4DL accounts for approximately 45% and chromosome 3BL accounts for approximately 11%) in this Atlas 66 cultivar. Recently, other genes that encode membrane transporters have been identified and characterized (Sasaki et al. 2004; Furukawa et al. 2007). These genes confer resistance to toxicity for Al in different cereals such as T. aestivum, H. vulgare, S. bicolor and other species (Table 1). Sasaki et al. (2004) identified a malate transporter, encoded by the major gene ALMT1, in near isogenic lines (NILs) of wheat. This gene might be constitutively expressed in both tolerant and sensitive genotypes although higher levels were observed in the plasma membrane of roots of ET8 lines (Al-tolerant). Location of this malate transporter was also confirmed by transiently-expressing ALMT1 fused to green fluorescent protein in onion epidermal cells and suspension cultured tobacco cells (Yamaguchi et al. 2005). In addition, heterologous expression of ALMT1 conferred Al-resistance to barley plants whereas, in O. sativa, the expression of this gene resulted in a significant increase in the efflux of Al-activated malate, but not in Al tolerance itself (Delhaize et al. 2004; Kikui et al. 2007). This may be due to the intrinsic properties of malate as this organic acid has a lower Al ion-chelating capacity in comparison to oxalate or citrate (Ma et al. 1998; Ma et al. 2001). ALMT1 cosegregates with Al tolerance in F2 and F3 populations derived from crossing isogenic T. aestivum lines (Sasaki et al. 2004; Zhou et al. 2007). In this way, genes that are differentially expressed between two NILs of wheat (Chisholm-T, tolerant and Chisholm-S, sensitive) have also been identified using suppression subtractive hybridization libraries. In this case, root tips from plants exposed during seven days to different Al concentrations were compared with non-treated control plants. Of a total of 1065 possible genes, 57 were differentially expressed during the first Al exposure period. Among these, 28 genes, including ALMT1, ent-kaurenoic, β-glucosidase, lectin, histidine kinase and phosphoenolpyruvate carboxylase, exhibited high amounts of transcripts in Chisholm-T, thus correlating with Al tolerance (Guo et al. 2007). These results suggest that Al-tolerance can be co-regulated not only by specific genes (such as ALMT1) but also by multiple genes with diverse functions in the plant. In addition, recent research using microarrays and NILs of wheat, identified 83 candidate genes associated with Al stress, such as pyruvate dehydrogenase, alternative oxidase and galactonolactone oxidase. In addition, 25 candidate genes which correlate with Al tolerance were identified, including ALMT1 as well as some unexpected genes such as glutathione S-transferase, germin/oxalate oxidase, fructose 1.6-bisphosphatase, cysteine-rich proteins, cytochrome P450 monooxygenase, cellulose synthase, zinc finger transcription factor, disease resistance response protein and F-box containing domain proteins (Houde and Oury Diallo, 2008), although no direct relation among them and the Al-resistance phenotype was established. ALMT1, the most recurrent gene observed in Al-tolerant T. aestivum cultivars is 3968 bp long, and is composed of six exons with a coding region of 1388 bp (Raman et al. 2005). The first 1000 base pairs upstream of the ALMT1 coding region is more variable, and six different patterns or alleles have been distinguished (Types I to VI). Moreover, all non Japanese cereal cultivars correlated positively with different levels of Al resistance (Sasaki et al. 2006). In addition, Raman et al. (2006) identified molecular markers targeting insertions/deletions (indels) within the intron-3 region of the ALMT1 gene. These ALMT1-SSR3a and ALMT1-SSR3b markers exhibited complete co-segregation with Al-resistance and malate efflux. Thus, these markers located in an intron and in the promoter region of TaALMT1 are useful tools to monitor the inheritance of the Al tolerance locus within specific T. aestivum populations. Recently, in H. vulgare, a dominant gene for Al tolerance (named Alt) has been mapped by amplified fragment length polymorphism (AFLP) and restriction fragment length polymorphisms (RFLP) markers. RFLP analysis has located this gene on the long arm of chromosome 4H (4HL), 2.1 cM proximally from the Xbcd1117 marker and 2.1 cM distal from the Xwg464 and the Xcdo1395 markers (Tang et al. 2000; Raman et al. 2002). On the other hand, Wang et al. (2007) using a high resolution map, identified the Alp locus, which is very close (0.2 cM) to the ABG715 and HvGABP markers. In addition, using double haploid and F2 populations from crosses between Dayton (tolerant) and elite sensitive cultivars, the HvMATE gene was identified as a gene belonging to the MATE family, which accounts for the Al tolerance in barley. In this same species, Furukawa et al. (2007) using both mapping and microarray analyses, identified the same gene (HvAACT1) in the Al-tolerant cultivar Murasakimochi. The gene product is responsible for citrate exudation and it is activated by Al. In this same study, a very high correlation was observed between HvAACT1 expression and citrate secretion in 10 H. vulgare cultivars with differing Al tolerance. Thus HvAACT1, as an Al-activated citrate transporter, is responsible for Al tolerance in H. vulgare (Furukawa et al. 2007). In other studies performed in rye, the most Al-tolerant cereal, four different genes related to Al-tolerance (Alt1, Alt2, Alt3 and Alt4) were discovered. These genes are located on chromosome 6RS, 3RS, 4RL and 7RS, respectively (Ma et al. 2000; Aniol, 2004; Fontecha et al. 2007; Matos et al. 2007). Another Al-tolerant species is X Triticosecale Wittmack (triticale), a hybrid resulting from the cross between T. aestivum and S. cereale. This cereal contains a complete genome copy of the rye chromosomes (AABBRR) that has given it the potential to grow and produce high yields in marginal soils such as those containing toxic levels of Al (Kim et al. 2001). Al tolerance was analyzed in two sets of hexaploid X Triticosecale Wittmack lines with disomic substitution of the D-genome of the T. aestivum chromosome. Of 20 substitution lines developed in winter X Triticosecale Wittmack (Presto) and 18 lines generated in spring X Triticosecale Wittmack (Rhino), six (30%) and nine (50%) lines, respectively, showed an increase in Al tolerance when they were compared with control lines (Budzianowski and Wos, 2004). Based on relative root growth (RRG) and mapping approaches, several QTLs for Al-tolerance have been identified in O. sativa. The lines were derived from crosses between the Al-tolerant Asominori and the Al-sensitive IR24 cultivars. The qRRE-1, qRRE-9 and qRRE-11 QTLs were detected on chromosomes 1, 9 and 11, respectively and the phenotypic variance ranged between 13.5% and 17.7% (Xue et al. 2006; Xue et al. 2007). The alleles of the Asominori cultivar from these three QTLs were all associated with an increase in Al tolerance. qRRE-9 was expressed in the genetic backgrounds of both IR24 and Asominori/IR24 (Xue et al. 2007). On the other hand, for two of the Al-tolerant sorghum cultivars, a unique locus named AltSB has been found to control this trait (Magalhaes et al. 2004). This locus might result in Al-tolerance through citrate exudation from roots. Consequently, by using positional cloning, a MATE gene conferring Al-tolerance in S. bicolor, has been identified (Magalhaes et al. 2007). Hoekenga et al. (2003) identified two major loci in Arabidopsis thaliana (arabidopsis), which account for approximately 40% of the phenotypic variance in Al tolerance observed among recombinant inbred lines derived from Landsberg erecta (sensitive) and Columbia (tolerant) ecotypes. Molecular markers and marker-assisted selection applied to Al-tolerance in cereals Molecular markers (MM) play an important role in the identification of desirable genes or alleles enabling genotype improvement. They also allow the structure and organization of the entire genome to be studied, as well as the physical mapping of BAC clones (Somers, 2004). In recent years, different dominant and co-dominant marker systems have been used. The dominant markers include Random Amplified Polymorphic DNAs (RAPDs) and Amplified Fragment Length Polymorphisms (AFLPs), whereas Restriction Fragment Length Polymorphisms (RFLPs), Simple Sequence Repeats (SSRs) and Single Nucleotide Polymorphisms (SNPs) are codominant markers. These markers have been developed for use with a range of crop species including cereals (Korzun, 2002; Korzun, 2003). Each type of molecular marker presents advantages and disadvantages. For instance, MMs based on PCR such as RAPDs, AFLPs, SSRs and SNPs require very small amounts of DNA, and are fast and easy to use, which facilitates the process of genotyping (Somers, 2004; Chao, 2006). On the other hand, other MMs such as RFLPs require large amounts of highly purified DNA, making the process more laborious. The Polymorphic Information Content (PIC), which gives account of the resolution power of a MM to distinguish different genotypes within populations, is another important feature of MMs. Currently, AFLPs and SSRs are the most popular MMs used in cereals because they provide a suitable genome coverage and PIC, respectively, at a reasonable cost. Nevertheless, with the advent of economically viable sequencing projects in model and economically important plants, SNP markers that represent sequence polymorphisms occurring at the single nucleotide level between varieties of the same species will be the markers of choice (Gupta et al. 2001). Using the current knowledge of SNP markers in cereals, it is possible to implement a high-throughput approach based on oligonucleotide arrays. The polymorphisms discovered from array hybridizations have been referred to as Single Feature Polymorphisms (SFPs) (Borevitz et al. 2003). Array-based SFP detection has been applied to several plant species including H. vulgare (Rostoks et al. 2005), O. sativa (Kumar et al. 2007), Solanum lycopersicum (tomato) (Sim et al. 2009) and Vigna unguiculata (cowpea) (Das et al. 2008). SFP discovery using an oligonucleotide array would be an efficient way to develop a large number of markers that may be used for high-resolution genetic mapping and marker-assisted breeding. The widespread use of DNA polymorphisms along with the growing technology of MMs has had a significant impact on plant improvement with regards to genetic diversity and genotyping studies, linkage map construction, trait tagging, gene cloning, and Marker-Assisted Selection (MAS). The MAS procedure is based on the concept of genetic linkage between two loci located close together on the same chromosome, resulting in co-inheritance or co-transmission to the progeny. Thus, by identifying the genotype of a specific marker, the phenotype of a linked locus might be deduced (Stam, 2003). The application of MMs for the selection of superior genotypes in plant breeding is most beneficial for those traits that are difficult to select phenotypically, are subject to high experimental error, or are expensive to assess (Kuchel et al. 2007), such as Al resistance. Since one of the aims of plant breeding is the introgression of one or more favorable alleles from a donor line into an elite variety, MAS enables the breeder to accelerate the recovery of the elite or recurrent parent genome (RPG) by backcrossing where only few rounds are necessary to introgress the target gene. Reductions in the time and expense of the whole process are additional positive aspects of MAS (Frisch et al. 1999; Tang et al. 2005). Comparison between conventional improvement and MAS are shown in Figure 1. Perhaps one of the most studied species regarding Al resistance is wheat. Riede and Anderson, (1996) established the association between the RFLP marker bcd1230 and the Al tolerance gene (AltBH) on 4DL at 1.1 cM. Subsequently, this information allowed SSR markers to be linked, which have facilitated the identification and introgression of this gene. Thus, the SSR markers WMC331, WMC457 and GWM165 have been tightly linked to the ALMT1 gene in different Al tolerant cultivars (Ma et al. 2004; Inostroza-Blancheteau et al. 2005; Raman et al. 2005; Zhou et al. 2007). By cloning the ALMT1 gene and analyzing the promoter region, Sasaki et al. (2004) and Sasaki et al. (2006) designed the marker ALMT1-4, which was used to identify six different alleles correlating 100% with levels of Al tolerance. In rye, efforts to identify MMs useful for the selection of Al tolerant genotypes have been reported by Gallego et al. (1998) using the SCAR markers ScR01600 and ScB157900, linked at 2.1 and 5.5 cM from the Alt1 locus, respectively. On the other hand, Collins et al. (2008) showed that the Alt4 Al-tolerance locus identified in the M39A-1-63/M77A-1 cross is located on the short arm of rye chromosome 7R. This agrees with the claim by Benito et al. (2005) that this locus was on 7RS, but disagrees with the chromosome 4R location originally determined by Miftahudin et al. (2002). The identification of an Alt locus on 4R has been recently reported by Benito et al. (2010). Matos et al. (2005) linked the RAPD markers SCIM8111376, SCIM812626, SCIM8121138 to the Alt4 locus. Using primers designed against the wheat TaALMT1 gene, Fontecha et al. (2007) amplified, cloned and sequenced an ALMT1 gene in rye located on chromosome 7RS that they named ScALMT1. Polymorphisms detected by SNP markers for this gene were detected among the parents of three F2 rye populations for the Alt4 locus. In H. vulgare, the most sensitive species of the Triticeae family, SSR markers such as Bmag353, Bmac310, HVM68, HVMCABG and ABG715 have been identified that are closely linked to the HvMATE gene (Raman et al. 2002; Furukawa et al. 2007; Wang et al. 2007) and represent a good set of putative molecular markers for use in identifying Al-tolerant plants. In diploid Avena strigosa (oat), RFLP markers linked to Al-tolerance have been identified through comparative mapping and QTL analysis, indicating that these QTLs are orthologous for Al-tolerance in other species (Wight et al. 2006). Currently, the availability of gene sequences related to Al tolerance have enabled the cloning of genes with similar functions in different cereals such as Triticum urartu (wheat), Aegilops speltoides (goatgrass), A. sativa, Z. mays and P. vulgaris, all sharing at least 72.3% similarity with the TaALMT1 sequences (Jardim, 2007). The knowledge of these sequences will substantially improve the potential to develop polymorphic molecular markers and apply them for breeding. Genetic maps to improve Al-tolerance in cerealsA major use of MMs in cereals is for constructing genetic maps by analyzing the co-segregation of markers and traits in defined populations (Korzun, 2002). Genetic maps match the positioning of markers in different linkage groups or chromosomes based on the percentage of genetic recombination that exists between two loci or markers observed in a segregating population (Dear, 2002). Since map distances are somewhat additive, the combination of distances for various genes or genetic marker loci for assembling maps is possible, so that the order of these genes/marker loci along chromosomes can be elucidated and therefore genetic maps can be constructed. Although genetic maps are crucial in the localization of genes and the estimation of the proportion of parental genomes present in the progenies, it is difficult to visualize simultaneously the whole genome for hundreds of plants and MMs across backcross populations. Fortunately, van Berloo, (1999) designed the software named Graphical Genotype (GGT) which allows the rapid visualization of molecular marker data in a user-friendly color format. The GGT concept was described by Young and Tanksley, (1989), and it has speeded up the modern genetic improvement for Al tolerance in cereals (Figure 1). Currently, the availability of well-saturated genetic maps facilitates the introgression of Al resistance in many cereals. For instance, a genetic map for wheat based on SSR markers has been developed (Röder et al. 1998; Gupta et al. 2002; Somers et al. 2004), and used to associated markers to the TaALMT1 gene on 4DL (Raman et al. 2005; Zhou et al. 2007; Cai et al. 2008). In H. vulgare, genetic maps based on SSR markers are also available (Liu et al. 1996; Ramsay et al. 2000). These maps have led to the localization of the HvMATE gene on 4H (Tang et al. 2000; Raman et al. 2002; Ma et al. 2004; Furukawa et al. 2007; Wang et al. 2007). More recently, a linkage map was developed in rye where locus Alt4 and the ScALMT1 gene for Al tolerance were located on chromosome 7R (Matos et al. 2005; Matos et al. 2007). In addition, several QTLs have been mapped in O. sativa (Nguyen et al. 2003) and Z. mays (Ninamango-Cardenas et al. 2003) which account for the Al tolerance observe in segregating populations. Today this valuable information could propel breeding programs around the world in order to satisfy the current demands for new Al tolerant cultivars of crops and cereals. Concluding RemarksConventional plant breeding is primarily based on the phenotypic selection of superior individuals among segregating populations. Although significant progress has been made in crop improvement through phenotypic selection, considerable difficulties are encountered during this process. The problem of genotype-environment interactions might generate unreliable data or expensive field experiments due to the nature of the target trait, for example, evaluation of abiotic stress such as Al toxicity. In the last 20 years, advances in molecular plant biology and more recently, in plant genomics have generated what some scientists call the new Green Revolution (Dubcovsky, 2004), which has lead to the development of molecular markers, genetic linkage maps and comparative mapping among related species such as T. aestivum, H. vulgare, S. cereale and O. sativa. This allows a much better understanding of their genomic organization, gene functions and helps to identify homologous genes with the same or similar effect, thus simplifying their cloning and fine mapping, and facilitating the application of marker (gene) assisted selection. The recent cloning of TaALMT1 (Sasaki et al. 2004) and HvAACT1 (Furukawa et al. 2007) that control the exudation of malate and citrate in Al-stressed T. aestivum and H. vulgare, respectively, are examples of these useful marker-trait associations. However, studies detailing successful implementation of molecular markers in pragmatic breeding programs are limited and they are often restricted to examples where the selection has focused on only one or two traits at a time (Kuchel et al. 2007). The current molecular information available regarding TaALMT1, AtALMT1, HvAACT1, ScALMT1 and ZmALMT1 and well-known in wheat, A. thaliana, H. vulgare, S. cereale and Z. mays Al-tolerant germoplasm, making it possible to generate new Al-tolerant varieties. Moreover, exploiting wild relatives or by using landraces as donor alleles would allow researchers and breeders to take advantage of polymorphisms between parent lines, identifying easily the target gene and the undesirable genomic regions coming from the donor parent, by using current molecular markers in conjunction with currently-available linkage maps. In conclusion, the next step should be the application of this knowledge in pragmatic breeding programs to overcome the Al toxicity problem in crops worldwide. Complementation between molecular technologies and conventional breeding is a crucial step for developing comprehensive research strategies aimed towards more efficient crop improvement in the near future (Figure 1). With the current rate of human population growth, cultivation of acidic soils will soon be necessary to satisfy the increasing food demand around the world. This issue concerns all important economic crops, therefore the extrapolation of the advances achieved in related species using genomic approaches, comparative mapping and bioinformatics, will be vital to accelerate the production of superior varieties for Al toxicity and other important traits. AcknowledgmentsThe authors would like to thank J. Larraín-Linton, Loreto Muñoz and Michael Handford for their assistance in language support. References
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10043t1.jpg] [ej10043f1.jpg] |
| |||||||||

{kind=link}
{kind=link}