 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 13, No. 5, September 15, 2010 Research Article Characterization of the antiyeast compound and probiotic properties of a starter Lactobacillus plantarum DW3 for possible use in fermented plant beverages Duangporn Kantachote*1, Pakorn Prachyakij2, Wilawan Charernjiratrakul2, Metta Ongsakul2, Yodsawee Duangjitcharoen3, Chaiyavat Chaiyasut3, Teruhiko Nitoda4,Hiroshi Kanzaki4 1Department
of Microbiology
Faculty of Science
Prince of Songkla University
Hat-Yai 90112, Thailand
E-mail: duangporn.k@psu.ac.th Financial support: This work was supported by National Science and Technology Development Agency (NSTDA), Thailand and the project numbers are CO-B-22-2C-10-483 and CO-B-22-2C-18-4801 including the Graduate School, Prince of Songkla University. Code Number: ej10046 Lactobacillus plantarum DW3 produced antifungal compounds that inhibited the growth of Rhodotorula mucilaginosa DKA, contaminating yeast in fermented plant beverages (FPBs) and various potential human pathogens. Phenyllactic acid (PLA) identified by gas chromatography- mass spectrometry (GC-MS) was produced at 31 mg/L PLA in MRS medium and 5 mg/ml inhibited growth of the target yeast in vitro by 90%. Other inhibitors were also present but not specifically identified. Results of in vitro tests showed that DW3 also had probiotic properties as it survived various human biological barriers resistance to pH 3, bile salts, growth without vitamin B12 and the presence and absence of oxygen. Its inhibitory effect against food borne pathogenic bacteria and spoilage organisms was higher than that found for a commercial strain Lactobacillus casei R. An acute oral toxicity test on ICR mice at a high single dose of either 109 and 1012 cells per mouse for 14 days showed that DW3 had no adverse effect on the general health status and there was no evidence of bacteremia. Mice fed DW3 had a reduced weight gain compared to the control. No significant difference (p > 0.05) was found for the spleen weight index (SWI) among the treatment and control groups whereas there was a significant difference (p < 0.05) for the liver weight ratio (LWR) in a group fed with 1012 cells per mouse when compared with the control group. Keywords: fermented plant beverage, Lactobacillus plantarum, phenyllactic acid, probiotics, Rhodotorula mucilaginosa, weight control.
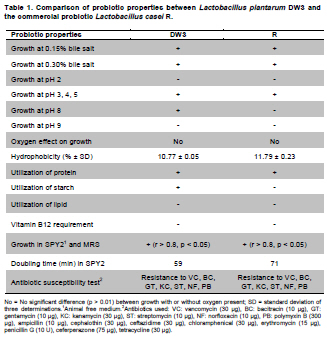
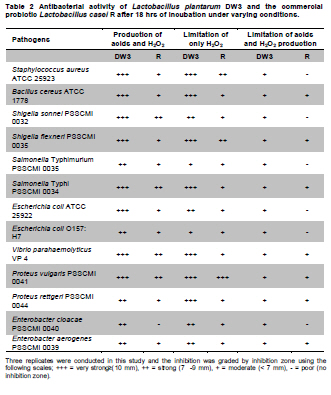
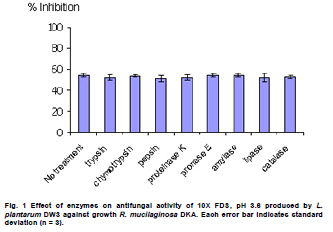
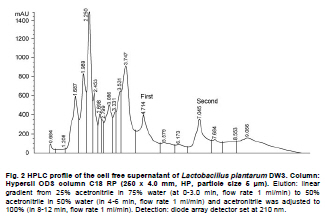
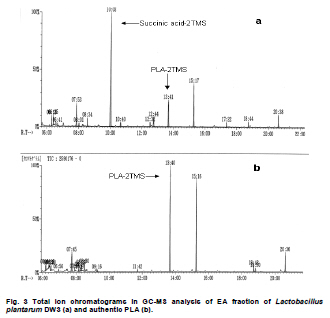
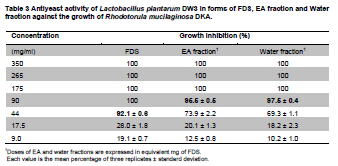
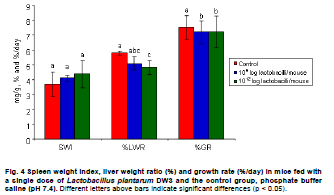
Fermented plant beverages (FPB or FPBs) are a traditional household product in Thailand. They are easily prepared by mixing plant material (cereals, fruits or vegetables) with sugar or honey and water in the proportions of 3:1:10 (w/w/v) and are incubated for 1-3 months with a restricted air supply (Kantachote and Charernjiratrakul, 2008a). These FPBs are nonalcoholic being produced by lactic acid bacteria (LAB) even though yeast contamination is frequently found in the finished product (Prachyakij et al. 2007; Kantachote and Charernjiratrakul, 2008a). They are considered to be a novel functional food product and have been recently approved as a local product by the Thai Industrial Standards Institute, Ministry of Industry. Contaminating yeasts that are frequently found in FPBs are Rhodotorula sp., Pichia sp., Hansenula sp., Saccharomyces sp. and Candida sp. (Prachyakij et al. 2007). Our previous studies have shown that using starter LAB cultures such as DW3 improved the quality of the finished FPB products (Kantachote and Charernjiratrakul, 2008b; Prachyakij et al. 2008). However, an antiyeast compound produced by our isolate DW3 should be identified. FPBs provide nutrients and can also provide antimicrobial compounds that originated from the raw materials used or as metabolites from the LAB itself (Kantachote and Charernjiratrakul, 2008a and Kantachote and Charernjiratrakul, 2008b). In addition, a probiotic LAB could be used as a starter cultures just as happens in the dairy industry. Many research workers have investigated probiotic lactobacilli e.g. (Reid, 1999; Vinderola and Reinheimer, 2003). Recently, the use of the probiotic Lactobacillus plantarum has been reported in food not containing milk or milk products (Molin, 2001; Goossens et al. 2006; Qin et al. 2007). Starter cultures could also be used in order to improve the quality of FPBs, by controlling yeasts in the finished products and to give beneficial properties to consumers. Therefore, a starter culture of L. plantarum DW3 previously isolated in our laboratory was studied for its probiotics properties in addition to characterizing its antiyeast metabolite. Preparation of cell free concentrated supernatant (Freeze dried supernatant: FDS) L. plantarum DW3, isolated from a fermented seaweed (Gracilaria fisheri) in our laboratory was selected as it has been shown to inhibit various food borne pathogens and produces antiyeast compounds (Prachyakij et al. 2007; Prachyakij et al. 2008). The isolate DW3 was inoculated into MRS broth and incubated at room temperature for 24 hrs, then adjusted to 1.5 x 108 cells/ml by dilution to 0.5 McFarland units. A 2% inoculum was added to a 400 ml MRS broth and incubated at room temperature (28 ± 3ºC) for 48 hrs without shaking. A cell free supernatant was obtained by centrifugation at 12,000 rpm (Sorvall RC 5C Plus) for 15 min and then filtered through a 0.45 µm cellulose acetate filter and finally concentrated by freeze drying. A 10-fold concentration of the freeze dried supernatant (10X FDS) was prepared and was sterilized by filtration through a 0.45 µm cellulose acetate filter prior to use. The isolate DW3 is able to inhibit a variety of yeasts such as Issatchenkia ocidentalis DKB, Pichia membranifaciens DKC, Pichia anomala DKD and R. mucilaginosa DKA (our preliminary study). However, R. mucilaginosa DKA was selected as a target yeast because it is the most common yeast contaminant in the final FPBs (Prachyakij et al. 2007). This yeast was also isolated in our laboratory and the microtiter plate well method according to Magnusson et al. (2003) was used for testing the antiyeast activity of DW3. Briefly, 100 µl of malt extract broth containing 104 yeast cells per ml were added to each well of a microtiter plate. 100 µl of test samples (i.e. 10X FDS, water and EA fractions see section GC-MS) were transferred to the corresponding wells and incubated at 30ºC for 48 hrs while 10X MRS served as a negative control. The yeast growth was measured by a microplate reader at a wavelength of 550 nm. The growth inhibition was calculated by comparison with the negative control of 10X MRS that showed no inhibition. This indicates that bioactive compounds in the samples tested are able to inhibit the target yeast. In order to investigate the possibility that the antiyeast compounds were proteins, lipids or carbohydrate compounds, the 10X FDS sample was treated with the following enzymes at a final concentration of 0.1 mg/ml; lipase in 0.05 M Tris-HCl (pH 8.0) with 0.01 M CaCl2,α-chymotrypsin in 0.05 M Tris-HCl (pH 8.0), trypsin in 0.05 M Tris-HCL (pH 8.0), pepsin in 0.2 M citrate (pH 6.0), pronase E in 20 mM Tris-HCl pH (7.8), proteinase K in 1 M NaOH (pH 6.5) and amylase in 1 M NaOH (pH 6.5). Catalase in 10 mM potassium phosphate (pH 7.0) was used to test for H2O2. All these solutions were filter-sterilized through 0.22 μm filters and then added to the 10X FDS (v/v, 1/1). The 10X MRS was also treated with the various enzymes and immediately sampled for use as a negative control whereas the positive control was a 10X FDS without addition of enzymes. The treatments and positive controls were incubated at 37ºC for 1 hr, except for the treatments and control containing trypsin, α-chymotrypsin and catalase that were incubated at 25ºC (Magnusson and Schnurer, 2001). Before evaluating the antiyeast activity, the pH of the supernatants was readjusted to the initial pH value 3.6 as this was the final pH of the cell free culture fluid. The remaining antiyeast activity was determined by the microtiter plate well method as described above. High performance liquid chromatography (HPLC) A rapid method of HPLC (Armaforte et al. 2006) was used to separate the antiyeast compounds in the cell free culture medium. The R-HPLC, was equipped with a quanternary pump, diode array detector set at 210 nm, a Hypersil ODS column C18 RP (250 x 4.0 mm, HP, particle size 5 µm) and run at room temperature. The filtered culture fluid was fractionated using a linear gradient of 25% acetronitrile in 75% water (pH 3) (at 0-3.0 min and a flow rate of 1 ml/min) up to 50% acetronitrile in 50% water (pH 3) (between 4-6 min and a flow rate of 1 ml/min) and acetronitrile was adjusted to 100% (between 8-12 min and a flow rate of 1.3 ml/min). Chem Station for LC 3D. Rev.A.10.02 (1757) was used for data acquisition and processing on a personal computer. Gas chromatography-mass spectrometry (GC-MS) An aqueous solution of FDS (1 g/ml in distilled water) was adjusted to pH 2 using 1 M HCl then extracted three times with a half volume of ethyl acetate (EA). The EA fractions were separated from the water fraction by centrifugation at 6000 rpm (Sorvall RC 5C Plus) for 10 min and this provided the EA and water fractions for testing antiyeast activity. Some portion of the organic extract was dried over Na2SO4 and concentrated in vacuo to obtain an EA fraction suitable for analysis by GC-MS. Trimethylsilyl (TMS) derivatization and GC-MS analysis of the EA fraction were performed according to the method described by Chaves and Gianfagna (2006). Briefly, the dried EA fraction (prepared from 16 mg of FDS) was treated with 50 µl of N, O-bis (trimethylsilyl) acetamide (BSA) at 70ºC for 15 min and then injected into the GC column without further purification. GC-MS analyses were performed using a Hewlet Packard (HP) gas chromatograph 5890 series, equipped with an Agilent column DB-1 MS (30 m long, 0.25 mm i.d., 0.25 µm film thickness) using helium as a carrier gas. The oven temperature was set at 70ºC, held for 2 min and increased to 240ºC at 10ºC per min and held for 30 min. The MS source temperature was set at 230ºC. The compounds corresponding to the peaks in the total ion chromatogram were tentatively identified by a library search of the JEOL data system (Lucy version 5.0 software, JEOL). This preliminary identification was confirmed by co-injection of an authentic compound. The amount of phenyllactic acid (PLA) was quantified by using four different concentrations of authentic DL-3-PLA (4-16 µg) and p-hydroxybenzoic acid was used as the internal standard. Investigation of probiotic properties in vitro The following series of investigations were carried out to provide evidence for the suitability of DW3 to perform as a probiotic when used as a starter culture for FPBs by comparing it with a commercial probiotic strain, Lactobacillus casei (R) (Nagao et al. 2000). Growth in MRS broth supplemented with 0.15 and 0.30% (w/v) bile salts (Fluka, Switzerland) and also tolerance to pH values of 2, 3, 4, 5, 8 and 9 was tested (Vinderola and Reinheimer, 2003). Growth as a facultative anaerobe was investigated in MRS broth, with the culture flasks incubated separately in either an anaerobic jar or in an aerobic incubator at 37ºC for 24 hrs. Bacterial growth was measured as turbidity using a spectrophotometer at OD660nm and all data were analyzed using the pair t-test. Adhesion of the isolate DW3 to hydrocarbons like n-hexadecane was determined for its hydrophobicity according to the method of Vinderola and Reinheimer (2003). The following agar plates of casein, starch and tributyrin were used to determine the utilization of protein, starch and lipid, respectively with any clear zone around bacterial colonies in each agar medium indicating that the organism had utilized the tested substrates. A requirement for vitamin B12 was examined in a vitamin B12 assay medium by comparing its growth to that of Lactobacillus delbrueckii subsp. lactis. Additionally, the time course of growth of both cultures in MRS and SPY2 (without animal derived ingredients) (Heenan et al. 2002) were examined and compared statistically as a correlation analysis. Antibiotic susceptibility tests to 15 antibiotics in common use (Table 1) were conducted by the agar disc diffusion method, while antibacterial activity against 13 strains of potentially pathogenic food borne bacteria and spoilage bacteria (Table 2) were carried out using the agar spot diffusion method (Kantachote and Charernjiratrakul, 2008a). Bacterial indicators used in this study were provided by the Microbiology Department, Faculty of Science, Prince of Songkla University, Thailand. Antibacterial activity of the isolate DW3 was conducted under 3 conditions according to the methods of Schillinger and Lucke (1989) as follows: production of organic acids and hydrogen peroxide (H2O2) (2% glucose in MRS agar and incubating under aerobic conditions), production of organic acids but limiting the production of H2O2 (incubating in an anaerobic jar) and limiting the production of both organic acids and H2O2 (0.2% glucose in MRS agar and incubating in an anaerobic jar). Safety assessment of L. plantarum DW3 in vivo The isolate DW3 was cultured in MRS broth at 37ºC for 18 hrs, cells were then centrifuged at 12,000 rpm (Sorvall RC 5C Plus) for 15 min. Cell pellets were washed twice in phosphate buffer saline (PBS), pH 7.4 and resuspended in PBS to obtain a cell density at approximately 109 and 1012 cells per mouse. Groups of five, 4-5 week old male ICR mice (Mus musculus) weighing 22-24 g were housed in transparent plastic cages. All cages were placed in a room controlled at 24 ± 2ºC, 55 ± 5% humidity and a light-dark cycle of 12 hrs throughout the study. Animal experiments were conducted at the Southern Institute Animal Laboratory, Faculty of Science, Prince of Songkla University, Thailand. In order to assess the safety of the DW3 isolate, a single 150 µl dose of lactobacilli was administered orally to each mouse and one group of 5 mice was fed with sterile PBS to serve as a control set. After feeding, mice were monitored daily for 14 days to observe any changes in their activities, behaviour and general health. Individual body weight was recorded daily using a balance. In addition, the feces of mice were collected to enumerate the total numbers of lactobacilli everyday for 14 days. The growth rate (GR) was calculated as % per day following the formula (GR: Wt/W0 x 100/t, Wt: body weight on the day of measurement, W0: body weight at the beginning and t: day of measurement). On day 14, all mice were sacrificed by cardiac dislocation and were bled by cardiac puncture. Blood was used to check and count bacteria for investigation of bacteremia while the liver and spleen were weighed. A spleen weight index (SWI) was performed as the actual spleen weight (mg) divided by the last measured live body weight (g) while the percentage liver weight ratio was expressed as the liver weight (g) divided by the body weight (g) multiplied by 100. One way analysis of variance (ANOVA) followed by Tamhane’s T2 was used to compare the results of the probiotic and control groups. Means, standard deviations and significant differences at p value < 0.05 are presented. Characterization of the antiyeast substance The 10X FDS produced a 55.3% inhibition of the growth of the target yeast (R. mucilaginosa DKA), in the positive control set without treatment by enzymes (Figure 1). After treatment with various proteases, the full antiyeast activity remained. Similar results were obtained after treatments with amylase, lipase and catalase. The results indicate that the antiyeast components of the FDS from the isolate DW3 were not proteins and were not related to carbohydrate or lipid moieties. In addition, catalase had no effect on the antiyeast activity therefore hydrogen peroxide (H2O2) was not one of the active compounds. Most LAB possess flavoprotein oxidases and NADH oxidase, that enables them to actively produce H2O2 in the presence of oxygen (Marty-Teysset et al. 2000). However, in this study, the isolate DW3 was grown under static condition with little access to oxygen; therefore, none or little hydrogen peroxide could be produced. To prove that the isolate DW3 may produce antiyeast compounds other than the organic acids such as lactic acid and acetic acid, HPLC was used to test for other antiyeast compounds. HPLC analysis of the cell free supernatant produced two major peaks at 4.714 and 7.045 min (Figure 2). However, only the first peak had antiyeast activity and the retention time corresponded to phenyllactic acid (PLA). Five mg/ml of the first peak showed strong inhibitory activity (88%) against the growth of the target yeast (data not shown). To confirm the presence of PLA in the culture filtrate, GC-MS analyses were performed. The EA fraction was analyzed by GC-MS after TMS-derivatization with BSA (N, O-bis (trimethylsilyl)-acetamide). The compound corresponding to the peak with retention time of 13.7 min (13 min 41 sec) was identified as PLA by comparing its electron ionization (EI)-MS spectrum with that recorded in the mass spectrum library (data not shown). The total ion chromatograms from the GC-MS analysis of the EA fraction and authentic PLA are shown (Figure 3). Apart from PLA, a succinic acid peak with a retention time of 10.0 min was found in the EA fraction. Production of succinic acid by the isolate DW3 was of interest because it can be a precursor of a variety of chemicals such as 1, 4 butanediol, γ-butyrolactone, tetrahydrofuran and some biodegradable polymers (Willke and Vorlop, 2004). Identifications were also confirmed by co-chromatography with the authentic PLA. The peak area of the compound (tR = 13.7 min) in the EA fraction was significantly increased when co-injected with the authentic PLA (data not shown). Besides, the peak for PLA another peak (tR = 15.3 min: 15 min 16 sec and 15 min 17 sec) was present in both the EA fraction and the authentic PLA (Figure 3), therefore we suspect that this compound was not a natural component of the FDS. Co-chromatographic analysis obviously indicated that the additional peak observed in both the EA fraction and the authentic PLA was the same compound because a significant increase of this peak area was observed after co-injection (data not shown). Hence, this compound might have been produced from PLA in the derivatization step. Role of PLA as an antimicrobial compound It has long been known that LAB play an important role in fermented foods as they produce a variety of compounds such as organic acids, hydrogen peroxide and bacteriocins that can control spoilage organisms and also potential pathogens (Messens and De Vuyst, 2002; Simsek et al. 2006; Tharmaraj and Shah, 2009). Among the bioactive compounds produced by LAB, PLA is a novel antimicrobial compound due to its ability to control both Gram positive bacteria and Gram negative bacteria, a wide range of fungi and some mycotoxigenic species (Lavermicocca et al. 2003, Prachyakij et al. 2008). Therefore, PLA is a biopreservative that could be used instead of propionic acid and its salt in bakery products. It is well recognized that PLA is produced by some LAB i.e. L. plantarum 21B and L. plantarum MiLAB393 (Lavermicocca et al. 2000; Strom et al. 2002) and in this study we have shown that PLA was also produced by L. plantarum DW3. The PLA content was determined by GC-MS analysis using p-hydroxybenzoic acid as an internal standard. The PLA content was found to be 0.85 µg/mg FDS, namely 31 mg/L of supernatant. Compared to the finding of Li et al. (2007), 60 of 112 strains of LAB produced between 16-61 mg/L PLA, L. plantarum DW3 is therefore a moderate producer of PLA. All forms of pure PLA (D-, L-, DL-PLA) require 5 mg/ml to produce 90% inhibition of the growth of R. mucilaginosa (data not shown). Table 3 shows that 44 mg/ml of pure FDS gave a 92.1% inhibition of growth of the target yeast, whereas the concentration of PLA contained in the FDS was only 0.031 mg/ml. Therefore, it can be concluded that PLA from the isolate DW3 was not the main antiyeast compound in the FDS. Additionally, results in Table 3 also show that the antiyeast activity of FDS was higher than that found in EA and the water fractions. This means that there are antiyeast compounds other than PLA in the FDS. In addition, the isolate DW3 has proved to be a successful starter culture to control contaminating yeasts in a fermented seaweed beverage (Prachyakij et al. 2008). Detection of probiotic properties in vitro Both the isolate DW3 and a commercial probiotic strain R, grew with both 0.15% and 0.30% bile salts in a modified MRS medium (Table 1). In addition, 90% of both strains survived a pH of 3 and 4 during a 3 hrs incubation period. The DW3 isolate also had a 50% survival at pH 8, but the R strain died. However, neither of them grew at pH 2 or 9. Biological barrier tests are required to prove a bacterium has probiotic potential because normally they must survive transit through the acidic conditions of the stomach and bile in the intestine (Vinderola and Reinheimer, 2003). In addition, ability to survive in the absence or presence of oxygen is another human physiological barrier that many possible probiotic bacteria in commercial products fail (Shah et al. 2000). Fortunately, both strains tested showed no significant differences (p < 0.01) in their growth in the presence or absence of oxygen. Again, the hydrophobicity of both strains tested was quite similar at 10.77 and 11.79% for the strain DW3 and the commercial probiotic strain, respectively. Other probiotic strains such as L. casei and Lactobacillus rhamnosus, have hydrophobicity values that range from 10.9 to 24.1% (Vinderola and Reinheimer, 2003; Tamang et al. 2009). Both strains also had no requirement for vitamin B12 as they grew to an OD660nm > 2.00 in a cobalamin deficient medium. This means that they would be unlikely to compete with the human body for cobalamin. A supply of vitamin B12 is especially important for vegetarians due to its low levels in their foods (Elmadfa and Singer, 2009). Some probiotic microbes display an array of enzymes that might be useful for improving human digestion. The DW3 isolate hydrolyzed both protein and starch, but the R strain hydrolyzed only protein probably because it was isolated from a commercial milk product. Hence, the isolate DW3 may provide more benefit to the human digestive system than the commercial strain R. Comparisons of the time course of growth and the relative viable cell counts between MRS and SPY2 broths showed that both strains had Pearson’s correlation coefficients (r) level > 0.8 (p < 0.05). Therefore, the isolate DW3 is suitable for use as a starter culture in any medium containing either animal or plant ingredients. It was also of interest that, the isolate DW3 had a shorter generation time of only 59 min in SPY2 medium compared to 71 min for the R strain. Hence, it is likely that the isolate DW3 can respond more quickly than the R strain to any possible source of substrate. The antibiotic susceptibility tests showed that both strains were resistant to 7 of the 15 antibiotics tested as follows: 30 µg vancomycin, 10 µg bacitracin, 10 µg gentamicin, 30 µg kanamycin, 10 µg streptomycin, 10 µg norfloxacin and 300 µg polymyxin B (Table 1). Results of a clinical trial have indicated that the incidence of antibiotic associated diarrhea and Clostridium difficile associated diarrhea can be reduced after consumption of a probiotic drink containing L. casei, Lactobacillus bulgaricus and Streptococcus thermophilus (Hickson et al. 2007). Therefore, co-administration to patients might facilitate a change in the imbalance of the intestinal microflora to become normal. Table 2 shows that the inhibitory effect of the isolate DW3 against 13 strains of food borne pathogenic bacteria and spoilage bacteria was higher than that found for the commercial strain R in all conditions tested. One of the important attributes for a lactobacillus strain to be an effective probiotic organism is that it has antimicrobial activity by producing organic acids, H2O2 and other inhibitory compounds (Reid, 1999; Ito et al. 2003). Isolate DW3 produces organic acids and other bioactive compounds like PLA. Thus it would be of interest to see if FPBs produced with starting cultures of probiotic bacteria together with some antibiotic would assist in curing some diseases. Safety assessment of L. plantarum DW3 in vivo As a safety assessment has been listed as an important criterion for selecting potential probiotic strains prior to being incorporated into food products (Zhou et al. 2000; Bernardeau et al. 2002), a mouse acute oral toxicity safety assay was conducted on the isolate DW3. Fourteen days after force-feeding mice with a very high dose (≈1012 cells per mouse, 3.15 x 1013 cells/kg), a high dose (≈109 cells per mouse, 3.17 x 1010 cells/kg) and the control group, there were no detectable differences in behaviour or activity and no diarrhoeal deaths. Also blood samples collected by cardiac puncture showed no viable bacteria on MRS or PCA from any mice. This clearly indicated that isolate DW3 did not exhibit gross acute oral toxicity effects on the general health status, growth and development of the mice tested and it is reasonable to conclude that the isolate DW3 showed no ability to infiltrate to or proliferate in areas outside of the intestines. Generally, adherence and colonization is considered to be an important property of probiotic strains and adherence can be considered to be the first step of colonization (Wold, 2002). In this study, hydrophobicity and a fecal count of lactobacilli were determined to assess the persistence of lactobacilli in mice to ensure that colonization has occurred. It was found that there were no significant differences for the numbers of lactobacilli in feces between the treated groups and control group (9 log cfu/g-10 log cfu/g) (data not shown) throughout the 14 days of the experiment. Results indicate that at least the isolate DW3 persisted in the intestines. However, after 14 days the growth rate of mice in both groups treated with the isolate DW3 was significantly lower than that found in the control group (Figure 4). However, the weight gain of all groups over the 14 days was within the normal value for mice (1.28 ± 0.29 g in the control group and 0.2 ± 0.05 - 0.25 ± 0.08 g in the probiotic groups) based on the report of The National Laboratory Animal Center (2007). This control of weight gain in the mice fed with L. plantarum DW3 is similar to that found in our previous work with L. plantarum SS2 in mice when there was also a reduced weight gain compared to a control (Duangjitcharoen et al. 2009). Recently, weight management is considered to be an additional positive role for probiotic microbes. For instance, Salminen (2009) reported that there is evidence that specific probiotics have an impact on weight gain and might help to control weight gain for instance during pregnancy. Probiotics might reduce fat storage by changing the intestinal microbiota. Obese subjects harbor more microbes of the phylum Firmicutes whereas lean subjects harbor more bacteroidetes (Ouwehand, 2009). Therefore, probiotics like bifidobacteria and lactobacilli may act by reducing Fermicutes. However, it would seem unrealistic to assume that obesity could be solved by using probiotic bacteria but it would be of interest to explore this possibility. Macroscopic examination of the visceral organs in the treated groups and the control group did not reveal any obvious differences in their size or appearance (data not shown). The spleen and liver weight ratios of mice fed with the isolate DW3 and the control groups are shown in Figure 4. There was no significant difference (p > 0.05) in SPI among the mice receiving the bacterium at both levels of cell density in comparison with the control mice that received PBS. The evidence of no change in spleen weights in the treated mice indicates that the isolate DW3 did not cause or induce systemic infections in experimental mice and being non-invasive is likely to be safe for human use. However, a very high dose of the bacterium (1012 cells per mouse) significantly decreased LWR as compared to the control group while there was no statistical difference between the two treated groups (109 and 1012 cells per mouse) (Figure 4). A similar result has been found in rats that received a dietary probiotic with a high fat and high cholesterol content. A significantly decreased liver weight was found in contrast to that found using dietary rice bran while the total cholesterol concentration in the serum of the probiotic group was significantly lower than that of the control group (Fukushima and Nakano, 1995). In conclusion, results in this study indicate that L. plantarum DW3 is a potential probiotic bacterium with an ability to control foodborne pathogens, spoilage organisms and contaminating yeasts in FPBs by producing a number of antimicrobial compounds including organic acids and phenyllactic acid and is likely to be safe for human consumption. Hence, the isolate DW3 is a promising strain to be used as a starter culture for improving the quality of the FPBs and providing significant benefits to the consumers. We would like to thank Dr. Brian Hodgson for his suggestions for improving the manuscript and checking the English.
Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. Copyright © 2010 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej10046t3.jpg] [ej10046f1.jpg] [ej10046f4.jpg] [ej10046f2.jpg] [ej10046t2.jpg] [ej10046t1.jpg] [ej10046f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}