 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
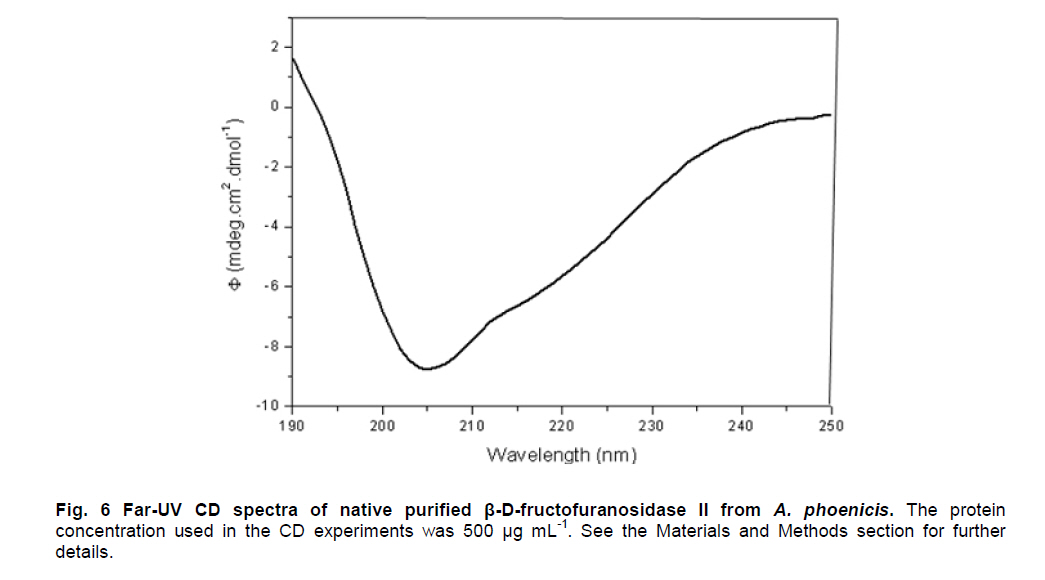
Electronic Journal of Biotechnology, Vol. 14, No. 2, March 15, 2011 RESEARCH ARTICLE Biochemical properties of an extracellular β-D-fructofuranosidase II produced by Aspergillus phoenicis under Solid-Sate Fermentation using soy bran as substrate Cynthia Barbosa Rustiguel1 · Arthur Henrique Cavalcanti de Oliveira2 · Héctor Francisco Terenzi1 · João Atílio Jorge1 · Luis Henrique Souza Guimarães*1 1Departamento
de Biologia, Faculdade de Filosofia, Ciências e Letras, Ribeirão Preto, Universidade de São
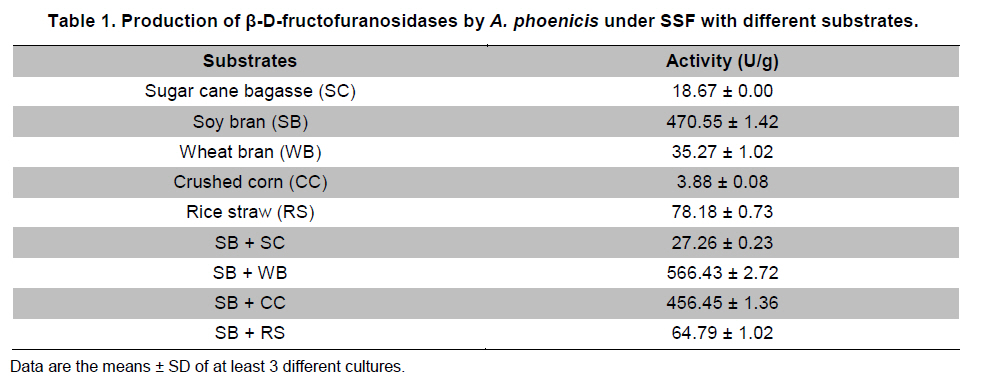
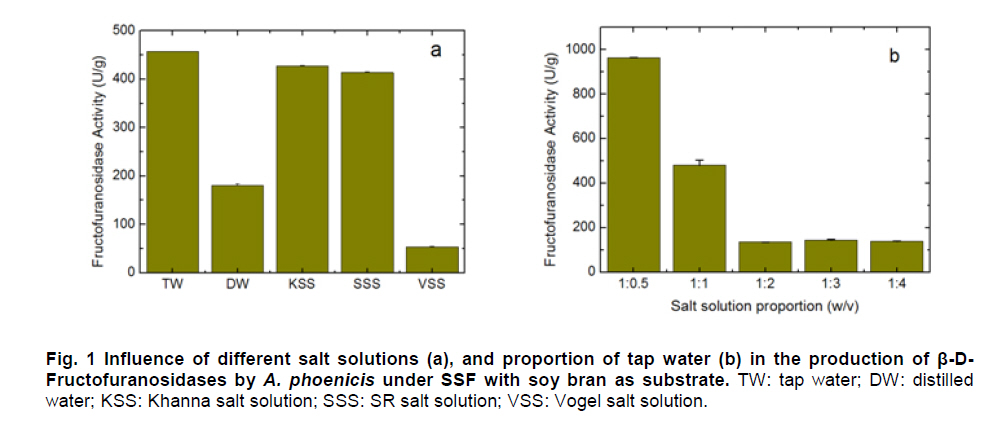
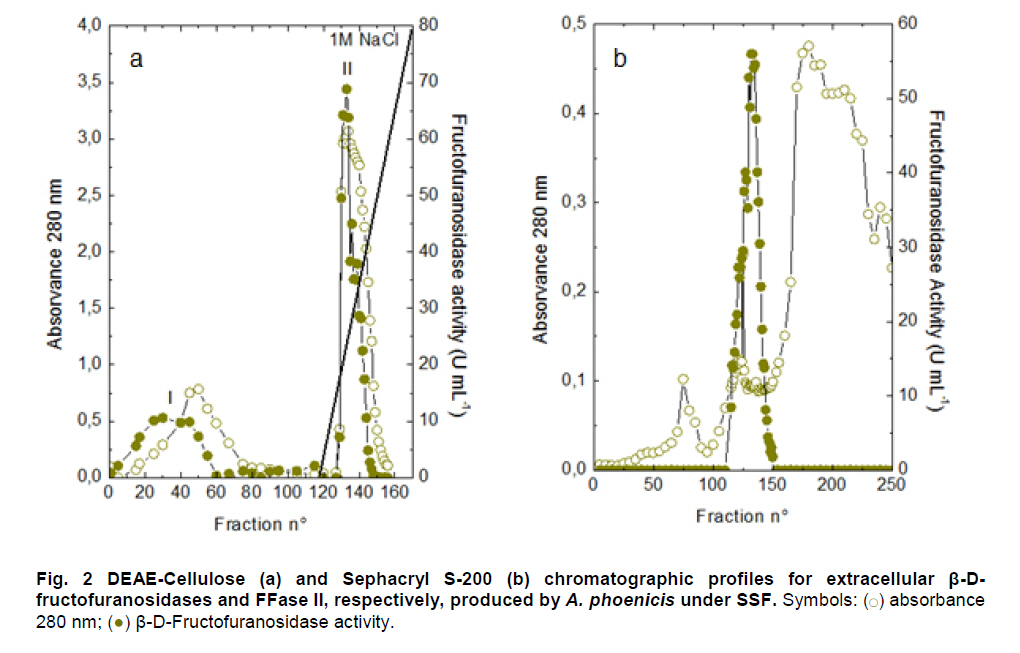
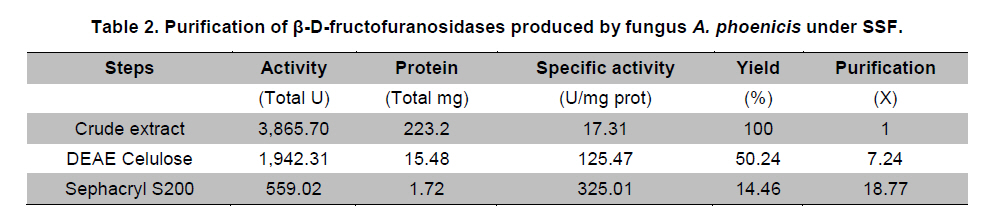
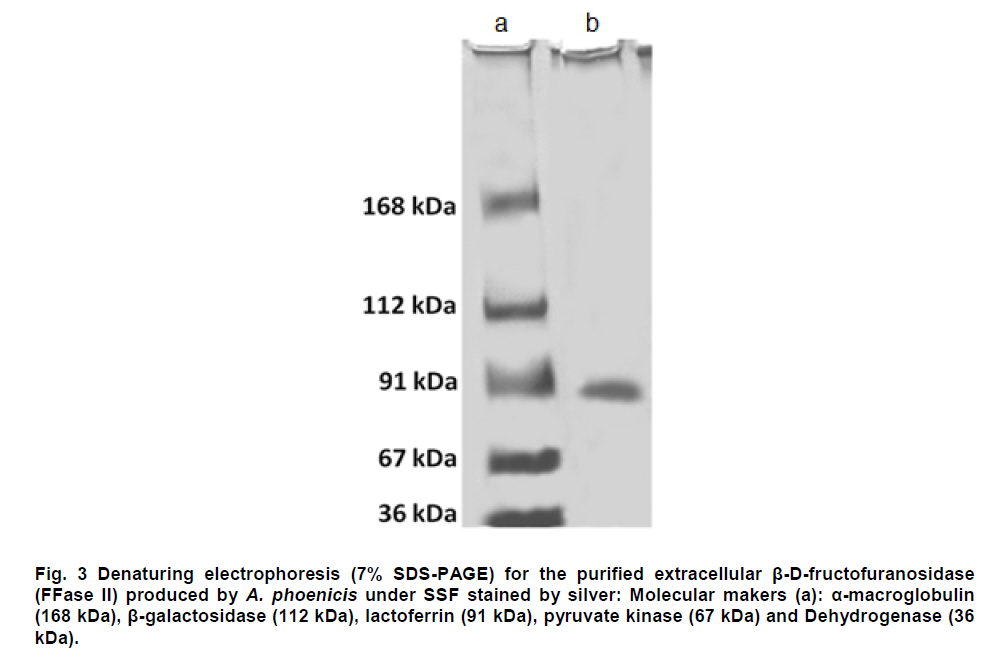
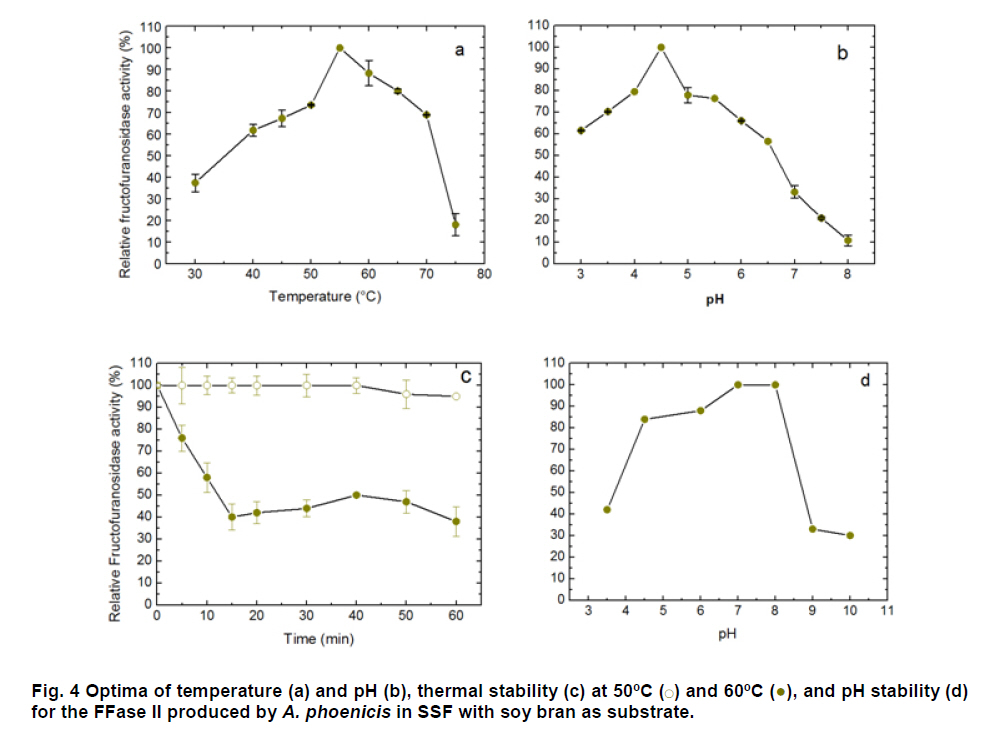
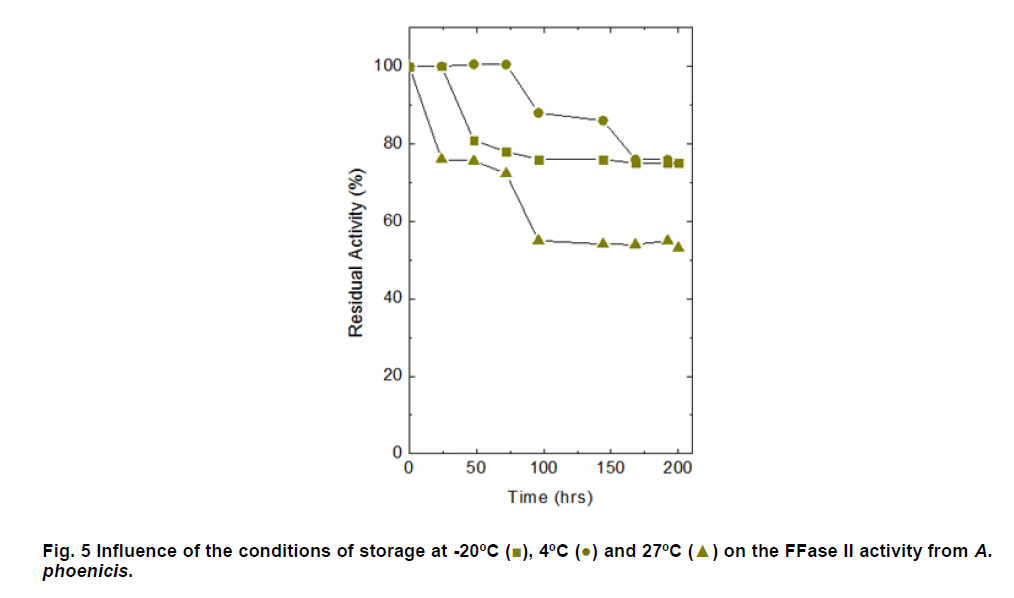
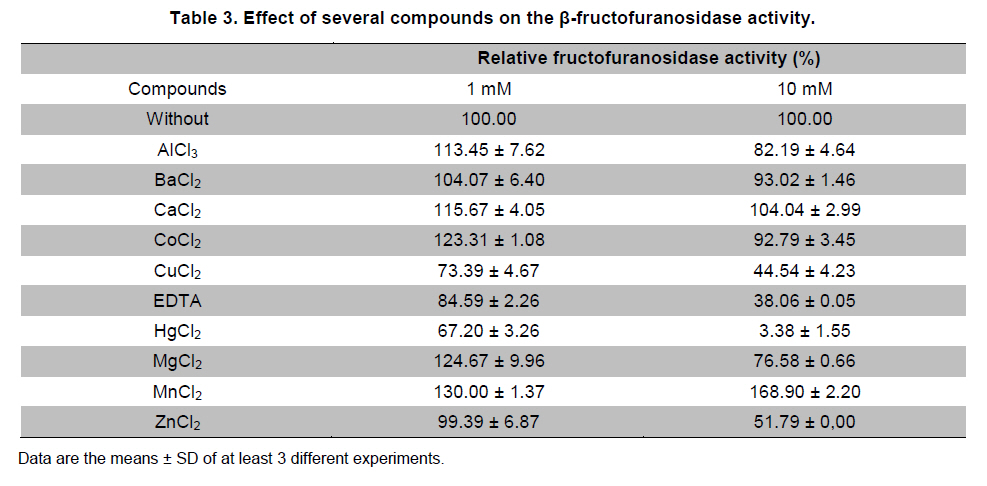
Paulo, São Paulo, Brasil *Corresponding author: lhguimaraes@ffclrp.usp.br Financial support: This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). Code Number: ej11014 AbstractThe filamentous fungus A. phoenicis produced high levels of β-D-fructofuranosidase (FFase) when grown for 72 hrs under Solid-State Fermentation (SSF), using soy bran moistened with tap water (1:0.5 w/v) as substrate/carbon source. Two isoforms (I and II) were obtained, and FFase II was purified 18-fold to apparent homogeneity with 14% recovery. The native molecular mass of the glycoprotein (12% of carbohydrate content) was 158.5 kDa with two subunits of 85 kDa estimated by SDS-PAGE. Optima of temperature and pH were 55ºC and 4.5. The enzyme was stable for more than 1 hr at 50ºC and was also stable in a pH range from 7.0 to 8.0. FFase II retained 80% of activity after storage at 4ºC by 200 hrs. Dichroism analysis showed the presence of random and β-sheet structure. A. phoenicis FFase II was activated by Mn2+, Mg2+ and Co2+, and inhibited by Cu2+, Hg2+ and EDTA. The enzyme hydrolyzed sucrose, inulin and raffinose. Kd and Vmax values were 18 mM and 189 U/mg protein using sucrose as substrate. Keywords: agroindustrial substrate, Aspergillus, β-D-fructofuranosidase, solid state fermentation, sucrose. IntroductionEnzymes with biotechnological potential, such as β-D-fructofuranosidases, can be produced by many organisms, especially microorganisms like bacteria, yeast and filamentous fungi. These enzymes are grouped in the GH-32 family of glycosidases that hydrolyze the alpha-beta linkage of carbohydrates with fructofurasyl residue, such as sucrose, producing a mixture of D-glucose and D-fructose known as invert sugar (Alberto et al. 2004). This catalytic property has attracted the attention of several industrial sectors, like as confectionery, food and beverage industries, which use fructose for formulation of candies, chocolates and soft drinks. Fructose is a monosaccharide with higher sweetener power and non crystallisable. In addition, it can be also used by diabetic people. Fructofuranosidases with fructosyltransferase activity can be used to produce fructooligosaccharides (FOS), as 1-kestose and 1-nystose, prebiotic substances that have important functional properties (Toledo et al. 2004; Maiorano et al. 2008). Microorganisms have been used to produce these enzymes, especially filamentous fungi such as Aspergillus niger (Rubio and Navarro, 2006), Aspergillus japonicus (Cheng et al. 2005), Aspergillus ochraceus (Guimarães et al. 2007), Aspergillus niveus (Guimarães et al. 2009) and Cladosporium cladosporioides (Almeida et al. 2005), among others. The production of β-D-fructofuranosidase by fungi is conducted under Submerged Fermentation (SbmF) or under Solid State Fermentation (SSF) (Romero-Gómez et al. 2000; Aranda et al. 2006). SSF has several advantages over SbmF, such as the similarity with natural conditions found by microorganisms, use of cheap substrates, as for instance agroindustrial residues and products, and higher levels of production. In addition, the water activity in SSF is an important factor for microorganism grown and enzyme production. Despite these positive facts, the control of culture conditions such as temperature and pH is very difficult. Hence, our aim was to characterize the β-D-fructofuranosidase II produced by Aspergillus phoenicis in SSF at high levels using agroindustrial products as alternative low cost substrate/carbon source. Basic information, as the biochemical properties, is important to determine the best conditions for enzyme application in the industrial process. Materials and MethodsMicroorganism and culture condition Aspergillus phoenicis (= Aspergillus saitoi) was isolated from soil and identified by André Tosello Foundation, Campinas, SP, Brasil. The microorganism was maintained on slants of PDA medium (Acumedia) at 40ºC for 7 days, and then stored at 4ºC. Solid-State Fermentation (SSF) cultures were carried out using different agroindustrial products as carbon source/substrate in 125 mL Erlenmeyer flasks with 8 g of substrate humidified with water or SR (Rizzatti et al. 2001), Khanna (Khanna et al. 1995) or Vogel (Vogel, 1964) salt solutions, in the proportion 1:1 (w/v), plugged with cotton and autoclaved for 20 min. The media were inoculated with 2 mL of a spore suspension (105 spores/mL) and maintained at 40ºC at relative humidity of 70% monitored with a digital thermo-hygrometer. Crude extract and enzymatic assay 50 mL of cold distilled water was added into SSF culture flasks and submitted to agitation for 30 min using a magnetic stirrer, at 4ºC. Then, the suspension was vacuum filtered through gauze. The obtained filtrate was centrifuged (23000 g) to remove particles and the clear supernatant was the source of crude β-D-fructofuranosidase activity. β-D-fructofuranosidase activity was determined using 1% sucrose as substrate in sodium acetate buffer, 100 mM, pH 4.5. The reaction mixture was composed by 200 µL of buffer containing substrate and 200 µL of diluted enzyme. The reaction was carried out at pre-determined times and temperatures (30ºC-75ºC). The reactions were stopped by addition of 3,5-Dinitro-salicylic acid (DNS) and the reducing sugars were quantified according to Miller (1959), at 540 nm. One unit of enzyme activity (U) was defined as the amount of enzyme that releases 1 µmol of glucose per min under the assay conditions. The values of enzymatic activity were expressed as U/g of substrate. Protein and carbohydrate quantification Protein was quantified according to Lowry et al. (1951) using bovine serum albumin (BSA) as standard. The carbohydrate content in the purified enzyme was determined according to Dubois et al. (1956), using mannose as standard. Purification The crude extract was loaded into a DEAE-Cellulose chromatographic column (10.0 cm x 2.0 cm) equilibrated with 10 mM Tris(hydroxymethyl)-aminomethan (TRIS)-HCl buffer, pH 7.5 (buffer A) and eluted with a NaCl linear gradient (0-1 M NaCl) in the same buffer. 3 mL fractions were collected at a flow rate of 1.9 mL/min and two enzymatic forms β-D-fructofuranosidase (FFase I and II) were obtained. The fractions with FFase II activity were pooled, dialyzed overnight against distilled water at 4ºC, lyophilized, ressuspended in 50 mM TRIS-HCl buffer, pH 7.5 containing of 50 mM NaCl (buffer B) and loaded into a gel sieving Sephacryl S-200 column (80.0 cm x 2.0 cm), equilibrated in the same buffer (B) and maintained in a cold room at 4ºC. 1 mL fractions were collected at a flow rate of 0.38 mL/min. The fractions with activity were pooled, dialyzed as mentioned above and used for biochemical characterization. Molecular mass determination The purified FFase II was submitted to denaturing electrophoresis (7% SDS-PAGE) performed according to Laemmli (1970) (120 V; 40 mÅ) and the protein bands were stained by silver nitrate (Blum et al. 1987). Molecular mass makers (Sigma®) were α-macroglobulin (168 kDa), β-galactosidase (112 kDa), lactoferrin (91 kDa), pyruvate kinase (67 kDa) and dehydrogenase (36 kDa). The native molecular mass was estimated by gel filtration using Sephacryl S-200 chromatographic column according to the conditions mentioned above. It was used alcohol dehydrogenase (150 kDa), BSA (66 kDa), egg albumin (43 kDa) and carbonic anhydrase (29 kDa), obtained from Sigma®, as molecular mass makers. Thermal and pH stabilities The purified enzyme was incubated for 1 hr at 50ºC or 60ºC, and also at different pH values using 100 mM sodium acetate buffer (pH 3.5 to 4.5), 50 mM Mes buffer (pH 6.0), 50 mM Tris-HCl buffer (pH 7.0 to 8.0) and 50 mM CAPS buffer (pH 9.0 to 10.0) and then assayed to determine the residual β-D-fructofuranosidase activity. The influence of the conditions of storage (-20ºC, 4ºC and 27ºC) at different periods (until 200 hrs) was also analyzed. Circular dichroism UV circular dichroism spectra (190-250nm) were measured with a JASCO 810 spectropolarimeter (JASCO Inc., Tokyo, Japan) using 1 mm-pathlength cuvettes at a protein concentration of 500 µg mL-1. All measurements were performed in 20 mM phosphate/citrate buffer (pH 5.5) at 25ºC. A total of ten spectra were collected, which after averaging were corrected by subtraction of a buffer blank. Determination of kinetic parameters The kinetic parameters as Vmax, Kd and Vmax/Kd (catalytic efficiency) were determined with sucrose as substrate (from 0.5 mM to 300 mM) using the Lineweaver-Burk representation and the data were processed by Sigraf software (Leone et al. 1995). Hydrolysis products analysis The purified enzyme was incubated with 20% sucrose as substrate in 100 mM sodium acetate buffer, pH 4.5 at 60ºC for 24 hrs. Samples were withdrawn at 0, 2, 12 and 24 hrs and submitted to thin layer chromatography analysis (TLC) (20 x 10 cm) using butanol:ethanol:water (5:3:2; v/v/v) as mobile phase. After the run, the plates were dried at room temperature, sprayed with a solution of 0.2% orcinol in sulfuric acid and methanol (1:9; v/v) and heated at 100ºC for visualization of the dyed spots. Five µL of 1% sucrose, 1% glucose and 1% fructose were used as standards. Results and DiscussionProduction and purification of β-D-fructofuranosidase A. phoenicis produced high levels of β-D-fructufuranosidase (470.5 U/g of substrate) under SSF using soy bran as carbon source/ substrate (Table 1). However, a mixture of soy bran and wheat bran increased by 20% the amount of enzyme. Probably, soy bran offered additional nutrients improving the fungus development and also increasing the production of the enzyme. High levels of β-D-fructufuranosidase by A. phoenicis were also obtained under submerged fermentation, but using wheat bran as carbon source (Rustiguel et al. 2010). The use of agroindustrial products or residues for SSF is an interesting alternative because it lowers cost for industries in the process of production of biomolecules, high recovery of the main product and availability of great variety of substrates. In addition, SSF reproduces the conditions found by microorganisms in natural environments, although it is difficult to control the cultivation conditions. Considering soy bran as substrate, the maximal production was obtained after 72 hrs of growth using 1:0.5 (w/v) tap water to humidify this substrate (Figure 1). The production under such conditions was 30-times higher than that obtained under SbmF (Rustiguel et al. 2010). According to Ashokkumar et al. (2001) the maximal production of β-D-fructofuranosidase by Aspergillus niger under SSF was also obtained after 72 hrs. Tap water may have salts that can contribute to the fungus development, which were not present in the other tested solutions. If the humidity of the medium is increased, the β-D-fructofuranosidase production diminished, perhaps due to lower aeration in the culture medium. The β-D-fructofuranosidase activity produced by A. phoenicis in SSF was resolved by DEAE-Cellulose chromatography into two enzymatic forms (FFase I and FFase II) (Figure 2a). FFase I did not interact with the resin and FFase II was eluted with 271 mM of NaCl. FFase II was submitted to Sephacryl S-200 chromatography, where it was eluted as a single peak (Figure 2b). After these procedures FFase II was purified 18-fold with a recovery of about 15% (Table 2). Interestingly, under SbmF, only a single enzymatic form was obtained, which interacted with DEAE-Cellulose (Rustiguel et al. 2010). Thus, the conditions used in SSF were able to permit the production of another β-D-fructofuranosidase form. Multiple β-D-fructofuranosidases forms have been reported for Aureobasidium pullulans (Yoshikawa et al. 2006). Aspergillus oryzae KB was able to produce two enzymatic forms, one preferentially produced at high sucrose concentration, and another at low sucrose concentration (Kurakake et al. 2008). FFase II was a glycoprotein with native molecular mass of 158.5 kDa and 12.23% of carbohydrate content, and constituted of two subunits of 85.11 kDa, according to denaturing electrophoresis (7.0% SDS-PAGE) (Figure 3). Homodimeric β-D-fructofuranosidases were also observed for A. ochraceus with two subunits of 79 kDa (Guimarães et al. 2007) and for R. glutinis, with subunits of 47 kDa (Rubio et al. 2002). For future biochemical characterization, the FFase I should be purified. Influence of temperature and pH The optimum temperature of FFase II produced by A. phoenicis under SSF was 55ºC (Figure 4a) and its optimum pH was 4.5 (Figure 4b). This temperature optimum was higher than that reported for the enzyme from Bifidobacterium infantis ATCC 15697 (Warchol et al. 2002), Lactobacillus reuteri (Ginés et al. 2000), Fusarium solani (Bhatti et al. 2006), Aspergillus niger IMI303386 (Nguyen et al. 2005) and A. oryzae KB (Kurakake et al. 2008), and similar with that of the β-fructofuranosidases from Aspergillus niger ATCC 20611 and Aureobasidium sp. ATCC 20524 (Yoshikawa et al. 2007). The optimum pH was similar to those of the enzymes from Aspergillus ochraceus (Guimarães et al. 2007) and Aspergillus niveus (Guimarães et al. 2009). The enzyme was stable at 50ºC, and exhibited a half-life of 13 min at 60ºC in aqueous solution (Figure 4c). The enzyme was also stable for more than 1 hr at pH 7.0 and 8.0 (Figure 4d), differing from the invertase from Rodotorula glutinis, which was stable between 20ºC and 60ºC, and at pH from 2.6 to 5.5 for 30 min (Rubio et al. 2002). In addition, the FFase II from A. phoenicis retained full activity after 75 hrs at 4ºC, and remained 80% active after 200 hrs at this temperature and also at -20ºC. The enzyme exhibited a t50 of 200 hrs at 27ºC (Figure 5). The condition of storage is another point that needs to be considered for industrial application, since products with good stability are preferred. Influence of ions and EDTA in the FFase II activity FFase II activity was enhanced by 1 mM of several divalent ions such as Mn2+ (30%), Mg2+ (24%) and Co2+ (23%), among others, and inhibited around of 16-33% by Cu2+, Hg2+ and EDTA (Table 3). However, at 10 mM concentration of these compounds, significant activation was only observed in the presence of Mn2+ (68%). Hg2+ inhibited drastically the FFase II activity. The enzyme produced by R. glutinis also was enhanced by Mg2+ and Co2+ (Rubio et al. 2002). Hg2+ and Cu2+ had negative effect on the invertase activity from L. reuteri (Ginés et al. 2000) and from A. ochraceus (Guimarães et al. 2007). The presence of ions in the protein solution could affect the pH, but it is known that the ions in solutions can interact with the charged amino acid groups in the protein (Kunz, 2006). This interaction may modify the conformational structure of the enzyme, or the ion may act directly on the active site, affecting the interaction with the substrate. Inhibition by Hg2+ is a signal of the presence thiol groups in the enzyme structure, important for its activity. Interestingly, FFase II showed around 67% of activity in the presence of 1 mM Hg+2, despite most enzymes are drastically inhibited by this ion. Circular dichroism The secondary structure of the purified β-D-fructofuranosidase II was evaluated by circular dichroism (Figure 6). The far ultraviolet spectrum of the purified protein shows a negative minimum at 205 nm and a shoulder at 216 nm. The features of the spectra indicate that the purified protein presents a high contribution of random and β-sheet structures in the secondary structure. This result indicates that the purification procedure maintained the secondary native structure of the protein. In addition, the fructofuranosidases from yeast Xanthophylomices dendrorhous (Linde et al. 2009) and from Arabidopsis thaliana (Verhaest et al. 2006) have propeller and β-sheet motifs, as verified for the enzyme from A. phoenicis. These characteristics are found in many glycoside hydrolases from GH32 family, which includes β-D-fructofuranosidases. Enzyme specificity and kinetic parameters FFase II from A. phoenicis was able to hydrolyze sucrose, inulin and raffinose (Table 4). The values of activity obtained with substrate mixtures did not correspond to the hypothetical sum of the values obtained when each substrate was tested separately, suggesting that these substrates are hydrolyzed at the same active site. The analysis by TLC showed only the products of hydrolysis glucose and fructose after 24 hrs of reaction at 60ºC, using 20% sucrose as substrate (data not shown). It has been cited that at low concentration of sucrose the hydrolytic activity is fostered and the fructosyltransferase activity could be observed at high concentration of sucrose (Chen and Liu, 1996). Under that condition, many fructofuranosidases exhibit fructosyltransferase activity being able to produce fructooligosacharides (FOS), as for instance the enzymes from Aspergillus sp. 27H (Fernández et al. 2004) and A. niger IMI 303386 (Nguyen et al. 2005). However, the FFase II produced by A. phoenicis in SSF had no fructosyltransferase activity. On the other hand, this characteristic permits the use of the enzyme in the production of invert sugar using high concentration of sucrose, increasing the recovery of product. The kinetic parameters Km and Vmax obtained with sucrose were 18.6 mM and 189.2 U/mg protein, with catalytic efficiency of 10 U/mg mM-1. The β-D-fructofuranosidase II from A. phoenicis exhibited higher affinity for sucrose than the β-fructofuranosidase I from Aureobasidium pullulans DSM 2404 (Km of 230 mM) and the fructofuranosidases from A. niger ATCC 20611 (Km of 290 mM) and Aureobasidium sp. ATCC 20524 (Km of 470 mM) (Nguyen et al. 2005). Concluding RemarksA. phoenicis produced two types of β-fructofuranosidase when cultured in SSF using an agroindustrial product as carbon source, what is very interesting for industrial application. The FFase II was a typical enzyme from the GH32 family, and exhibited properties that could justify its use for industrial production of invert sugar associated with the low cost of the carbon source/substrate, high affinity for sucrose, added to good thermal and storage stabilities. AcknowledgmentsWe thank Prof. Dr. Richard J. Ward for access to the Jasco 810 spectropolarimeter and Maurício de Oliveira for technical assistance. References
Copyright © 2011 by Pontificia Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11014t1.jpg] [ej11014f2.jpg] [ej11014f1.jpg] [ej11014t4.jpg] [ej11014t2.jpg] [ej11014f3.jpg] [ej11014t3.jpg] [ej11014f5.jpg] [ej11014f6.jpg] [ej11014f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}