 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 3, May 15, 2011 Developmental rates of bovine nuclear transfer embryos derived from different fetal non transfected and transfected cells Ricardo Felmer D.*1 · María Elena Arias1 1Laboratorio de Biotecnología Animal, Instituto de Investigaciones Agropecuarias, INIA - Carillanca, Temuco, Chile *Corresponding author: rfelmer@inia.cl Financial support: This research was supported by grants from FONDECYT 1080216 and FONDEF D03I1074. Received: December 12, 2010 Accepted: March 14, 2011 Code Number: ej11028 Abstract Since the first successful somatic cell nuclear transfer (SCNT) experiments were carried out, a number of domestic and agriculture species have been cloned using donor cells derived from different sources and origin. However, differences in nuclear transfer efficiency both in vitro and in vivo have been generally observed. These differences may be accentuated when transgenic cell lines are used as nuclear donors in an attempt to generate transgenic cloned offspring. The present study examined the suitability of cell lines derived from 3 different fetal sources and the effects of genetic manipulation of donor fetal fibroblasts with a red fluorescent plasmid, on the in vitro developmental potential and quality of nuclear transfer derived bovine embryos. We observed no differences in the cleavage rate of nuclear transfer embryos generated with any of the cell lines evaluated. However, the blastocyst rate was significantly affected when cell lines were derived from the 3 different fetal sources (21, 18 and 11%, respectively) or from 2 transgenic clonal cell lines that had originated from the same primary fetal cell (18 and 10%, respectively). Despite this difference, quality of embryos as measured by the total number of cells and by assessing some morphology aspects of their appearance was not different. Together these results indicate that fetal fibroblast cell lines derived from different fetal sources and transgenic clonal cell lines that had originated from the same fetus results in different in vitro developmental potential when used as donors for nuclear transfer experiments. Further studies, including evaluation of pregnancy rates, development to term, and epigenetic modifications of these cell lines will be necessary to better understand the differences observed in nuclear transfer efficiency. Keywords: bovine embryos, fetal
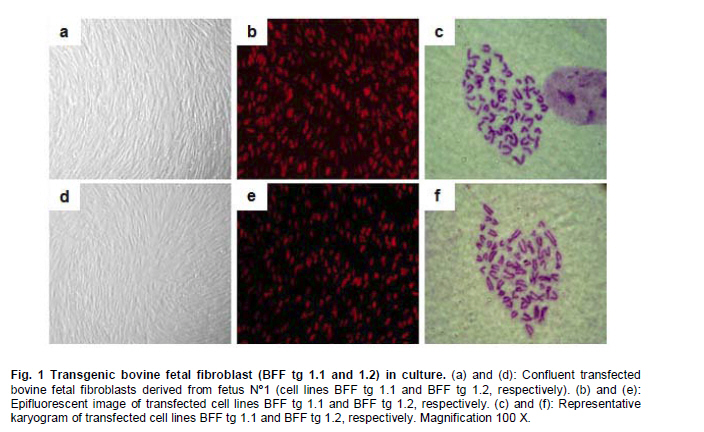
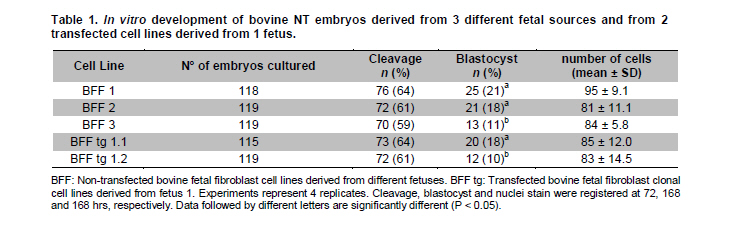
fibroblasts, nuclear transfer, transfected cells. Cloning by somatic cell nuclear transfer (SCNT) has been successfully achieved in a number of domestic and agriculture species (for a comprehensive review see Campbell et al. 2007). A range of embryonic, fetal or adult cells have been used as nuclear donors, including mammary gland cells, cumulus cells, oviductal cells, leukocytes, granulosa cells, germ cells, liver cells and muscle cells (Brem and Kühholzer, 2002). However, it is still unclear which cell type is the most successful for nuclear transfer into oocytes and despite the success with these cells, different efficiencies of in vitro and in vivo development of nuclear transfer embryos have been generally observed (Zakhartchenko et al. 1999; Inoue et al. 2003; Poehland et al. 2007). For instance, when the efficiencies of various cell types from adult, newborn and fetal donor cells were compared in cattle, no significant difference were found in the percentage of embryos reaching to the blastocyst stage (Kato et al. 2000). Similar results were also observed in mice when macrophages, fibroblasts and spleen cells were used (Wakayama and Yanagimachi, 2001). However, a number of studies have also shown significant differences depending on the cell type source, age and/or sex of the donor cell used for nuclear transfer (Lagutina et al. 2005). Nuclear transfer technology also allows the opportunity for animal genetic manipulation (Clark, 1998; Houdebine, 2009). In order to do this, nuclear donor cells must be genetically manipulated in vitro to target the desired modification in the animal. Therefore, an extended period of culture is required for selection of transgenic cells resistant to the marker gene. This extended period of culture may have profound effects on the developmental potential of transgenic cells used as nuclear donors as described by a number of reports (Zakhartchenko et al. 2001; Arat et al. 2002; Iguma et al. 2005; Wang et al. 2007). Therefore, in the present study we examined the effect of cell lines derived from different fetal sources and genetic manipulation of these cells, on the in vitro developmental potential and quality of nuclear transfer derived bovine embryos in order to establish if donor source, transfection and longer culture of these cells, could have an effect on the developmental potential of nuclear transfer embryos. Materials and Methods Derivation and transfection of donor cells Bovine fetal fibroblasts (BFF) were isolated from 3 different Holstein female 50-70 days old fetuses, recovered at a local slaughterhouse. Cell explants were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco) supplemented with 10% FCS (HyClone) at 38.5ºC in a humidified atmosphere of 5% CO2. When cells reached confluency, they were collected by trypsinization and frozen for future nuclear transfer experiments. A primary cell line derived from the female fetus 1 (BFF 1) was used for transfection experiments on passage 5. Two µg of a linearized plasmid (Afl II digested) containing the red fluorescence protein marker (pHcRed1-nuc; Clontech) was combined with Lipofectamine (Invitrogen), according to the manufacturer instructions. Cells were selected with 800 µg/ml Geneticin (G418, HyClone) for 7-10 days and single colonies isolated by ring cloning. A portion of cells was used for DNA isolation, confirmation of construct integration by PCR and chromosome analysis. Remaining cells were frozen for future nuclear transfer experiments. Oocyte collection, maturation and nuclear transfer Bovine ovaries were collected at a local slaughterhouse (Frigorifico Temuco) and grade 1 cumulus oocyte complexes (COCs) were selected for maturation, that was carried out in Medium 199 (Sigma) supplemented with 10% FCS (HyClone) and hormones at 38.5ºC in a humidified atmosphere of 5% CO2 in air. After 17 hrs of maturation, oocytes were stained with Hoechst 33342 (Sigma) and enucleated by aspiration of the MII plate using an inverted microscope (Nikon TS100F) and Narishige micromanipulators. Prior to nuclear transfer, donor cells (transfected and non-transfected) were grown to confluency for 5 days to induce quiescence. These cells were microsurgically placed into the perivitelline space evacuated during enucleation and cell-cytoplast complexes were fused in sorbitol media with a single DC pulse of 170 volts/mm and 15 µseconds delivered by an Electrocell Manipulator 830 (BTX). Activation was carried out with 5 µM Ionomicin for 5 min followed by incubation in KSOM (Millipore) media containing cycloheximide (10 µg/ml, Sigma) and cytochalasin B (5 µg/ml, Sigma) for 5 hrs. After activation, the nuclear transfer units were cultured in 30 µl drops of KSOM media under mineral oil at 38.5ºC and at 5% CO2, 5% O2, 90% N2. On day 3 (nuclear transfer = day 0) cleavage rate was recorded and embryos were transferred to KSOM+5% FCS and cultured until day 7, time when blastocyst rate and total number of cells were recorded. All the cell lines were evaluated on the same day and the experiments were replicated 4 times. Cell number count and statistical analysis Cell number of blastomeres in day 7 blastocysts was scored by incubating embryos in media (TCM-199) containing 10 µg/ml bisbenzamide in absolute ethanol at room temperature for 10 min. Blastocysts were then treated in 50% (v/v) glycerol, mounted onto a glass slide, and visualized under epifluorescent microscope coupled with a Filter UV-2E/C DAPI. Data analysis for embryo development and cell count was carried out by descriptive statistics based on average and standard error calculated for each of the variables, using Stat graphics Plus 5.1 Software. One-way ANOVA was used to test for statistically significant differences (P < 0.05) among groups for cleavage, blastocysts rate and cell counting. In cases where statistically significant differences were observed, the least significant difference test (LSD) was invoked to determine where those differences existed. ResultsBovine fetal fibroblast (BFF) cell lines were successfully established from all 3 fetuses recovered at the slaughterhouse (described herein as BFF1, BFF2 and BFF3, respectively). Cell lines were cultured up to passage 4 and nuclear transfer experiments were carried out with these cells on passage 5-6. Stably transfected cells were also generated with the marker gene pHcRed1-nuc, allowing selection both by resistance to neomycin and the intensity of red fluorescence protein signal (Figure 1). Fifteen neomycin resistant cell lines were generated, 5 of which expressed red fluorescent protein at a higher level. Karyotyping analysis of these cells showed a normal chromosome complement (2n = 60; XX) in 4 out of 5 cell lines analyzed. This analysis allowed the selection of 2 normal transfected cell lines for further nuclear transfer experiments (Figure 1; clones BFF-Tg 1.1 and BFF-Tg 1.2, respectively), that were used on passage 9-10. The assessment of transgenic cloned embryos expressing red fluorescent protein during the embryonic development up to the blastocyst stage, was described previously (Felmer and Arias, 2008). The efficiency of nuclear transfer generated bovine embryos was compared using these transfected and non-transfected bovine fetal fibroblast cell lines on independent replicates. No significant difference in terms of cleavage rate was observed between embryos generated for any of the transfected and non-transfected cell lines (59 to 64%). However, a significant difference in the rate of blastocyst generated from cell lines derived from the non-transfected and/or transfected group was observed (Table 1). One transfected (BFF tg 1.2) and one non-transfected (BFF 3) cell line had the lowest rate of blastocyst (10 and 11%, respectively), while transfected (BFF tg 1.1) and non-transfected (BFF 1 and 2) cell lines had the best developmental capability (21, 18 and 18%, respectively). The quality of nuclear transfer embryos generated by these cells, as measured by the total number of cells and by assessing some morphology aspects of their appearance, was not different for any of the cell lines used as nuclear donors. DiscussionIn this report, we investigated the suitability of cell lines derived from 3 different fetal sources and the effects of genetic manipulation of these cells on the in vitro developmental potential of nuclear transfer derived bovine embryos. We chose to use fetal cells as donors for nuclear transfer experiments, because these cells are believed to have less genetic damage and better proliferative ability than adult somatic cells and therefore are more robust for transfection experiments. We observed a significant effect between cell lines derived from different fetal sources despite the fact that all cell lines belonged to the same breed of origin, were isolated within the same day and were cultivated under equal culture conditions. Furthermore, all the cell lines were at the same passage number by the time of nuclear transfer experiments were carried out (between 5-6 passages) and were subjected to the same cell cycle synchronization protocol, as described in Material and Methods section. Although there was no difference in the cleavage rate for any of the fetal fibroblast cell lines used, a significant difference was observed in the rate of blastocyst that was consistently higher for 2 of the 3 fetal fibroblast cell lines assessed (21, 18 and 11% for BFF 1, BFF 2 and BFF 3, respectively). Results described previously using different primary cell lines of adult bovine somatic cells indicated that the primary donor cell culture affects in vitro blastocyst development, initial pregnancy rates, and the percentage of live births (Miyoshi et al. 2003; Poehland et al. 2007). Similar results were obtained in pig cloning studies, where not only different cell lines, but also different clones derived from one primary cell line used as nuclear donors resulted in different development beyond day 90 of gestation (Kühholzer et al. 2001) and in horse cloning studies where cumulus, granulose, fetal and adult fibroblast cells had all in vitro and in vivo differential developmental potential (Lagutina et al. 2005). However, contradictory results have also been observed when the efficiency of nuclear transfer using a number of somatic cell types from adult, newborn, and fetal female and male donor cattle were compared with no difference in the percentage of embryos that developed to the blastocyst stage being reported (Kato et al. 2000). Similar results were also observed using various cell types derived from mice of different strains, sexes and ages (Wakayama and Yanagimachi, 2001). This marked difference observed for any of the cell types and/or cell lines used in nuclear transfer experiments may be due to epigenetic effects, because even within a primary cell culture, the generation of cell lines from that culture showed some lines more suitable than others for nuclear transfer (Kühholzer et al. 2001). Modifications that may occur during cell culture might result in epigenetic changes to the genomes making these cells more or less capable of being reprogrammed. In fact, wide-spread abnormalities in gene expression in embryos and cloned animals are beginning to emerge, which suggests that transcriptional regulation mechanisms were impaired possibly at different levels during the reprogramming process (Dean et al. 2001; Beyhan et al. 2007; Suzuki et al. 2008). Transgenic cloned offspring has been produced by SCNT in sheep, cows and other species (Schnieke et al. 1997; Cibelli et al. 1998; Bordignon et al. 2003; Echelard et al. 2009). The generation of transgenic animals by this approach requires the genetic manipulation of donor cells, a process that has been observed to cause a dramatic effect on the efficiency of the nuclear transfer technique and in some cases in the failure of generating live offspring (Iguma et al. 2005; Jang et al. 2006). This situation rise the question of whether the efficiency of SCNT in generating transgenic animals would be affected by the transgene itself or the modified donor cells. In this report, we carried out cytogenetic analysis of transfected BFF cells expressing a red fluorescent protein, since it has been reported a direct relationship between chromosomal anomalies in nuclear transfer embryos and in their respective nuclear donor cells (Bureau et al. 2003). A clonal cell line with abnormal chromosome complement was found within 5 clonal cell lines analyzed, which allowed to exclude this cell line for future nuclear transfer analysis. Remaining 4 clonal cell lines, had all normal chromosome complement (2n = 60; XX). Two of these transfected clonal cell lines were chosen for nuclear transfer evaluation. These cell lines (BFF tg 1.1 and BFF tg 1.2, respectively), which derived from the same primary culture (BFF 1), had a significantly different developmental rate to the blastocyst stage (18 v/s 10%, respectively), although cleavage rate was not affected (64 and 61%, respectively). Interestingly, one of the transfected cell lines (BFF tg 1.1) showed no differences in the cleavage (64 and 64%, respectively) and/or blastocyst rate (18 and 21%, respectively), when compared to the primary culture from which it was derived (BFF 1), while the other one (BFF tg 1.2) had a significant decrease in the rate of blastocyst (10 v/s 21%, respectively). This result indicates that transfection and longer passage number of transfected cells not necessarily affect the nuclear transfer efficiency in vitro, but this effect is more dependent on each clonal cell line, due to differences in the site of integration, number of integrated transgene copies, and/or other epigenetic effects as result of the in vitro manipulation. It was previously observed a significant difference in the efficiency in generating nuclear transfer embryos when transfected ear fibroblasts were used as compared to the non-transfected counterpart, regardless of passage number (Bhuiyan et al. 2004). However, contradictory results were observed by others, with no differences in fusion, cleavage, blastocyst or pregnancy rates between transfected and non-transfected ear fibroblasts (Iguma et al. 2005). This discrepancy with transfected cells, has been suggested to be due to differences in vector type and transfection protocols (Bhuiyan et al. 2004), donor cell culture conditions (Wells et al. 2003), and the transgene and/or site of transgene integration (Hodges and Stice, 2003). In fact, a recent study with transfected cells harboring different foreign genes (hFIX, hALB and hTF), confirmed this observation since significant differences in fusion, cleavage and/or blastocyst rate was observed when these cells were used in nuclear transfer experiments (Fu et al. 2008). In conclusion, the results presented here suggest that cloning efficiency is not only affected by the source of cells used as nuclear donors. Variations in nuclear transfer results between experiments and between different cell lines as described here and elsewhere, are probably due to deviations in the epigenetic regulation of the gene expression profile of each donor cell line, as result of genetic disturbances resulting from the culture and/or manipulation conditions. Therefore, whether transfected and non-transfected cells are used, it is necessary first to assess the in vitro developmental potential of each donor cell line in order to select those with the highest rate of blastocyst development and thus the most suitable for nuclear transfer experiments, bearing in mind that a high rate of blastocyst in vitro will not necessarily correlate with implantation and normal development of these embryos after transfer. Further studies, including evaluation of pregnancy rates, development to term, and epigenetic modifications of these cell lines will be necessary to better understand the differences observed in nuclear transfer efficiency. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11028f1.jpg] [ej11028t1.jpg] |
| |||||||||

{kind=link}
{kind=link}