 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
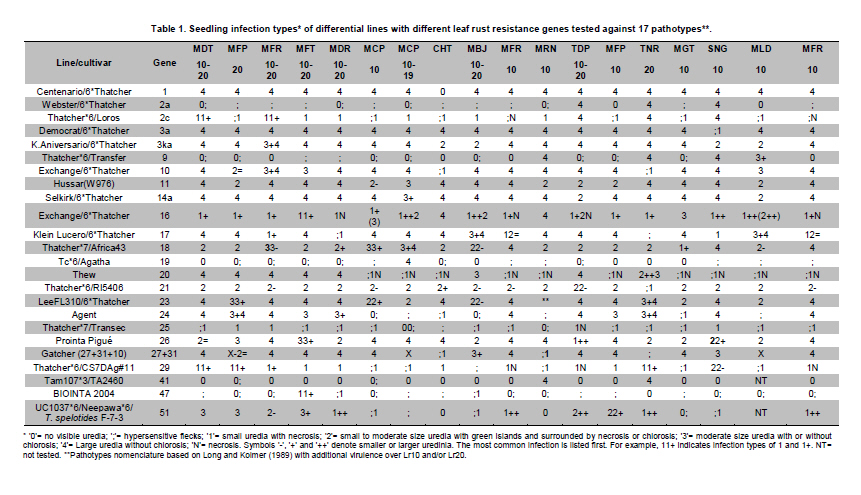
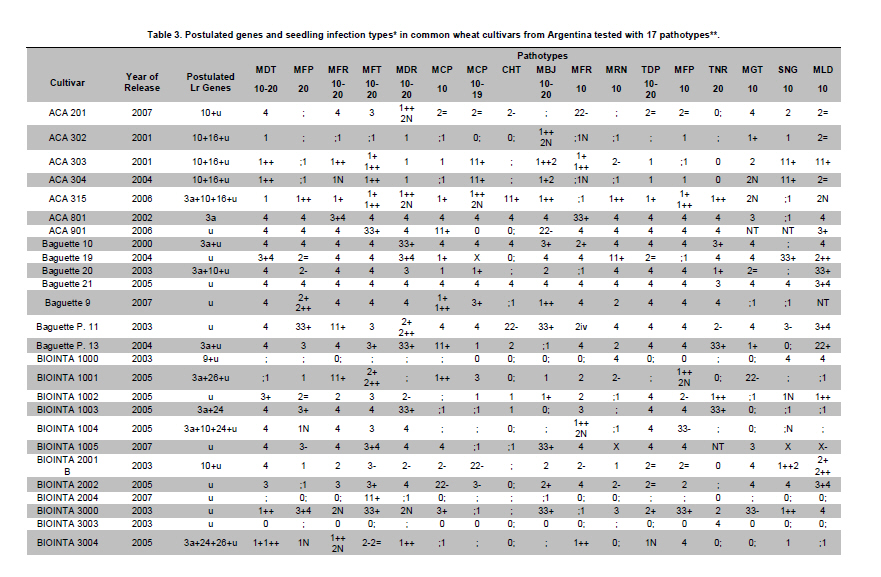
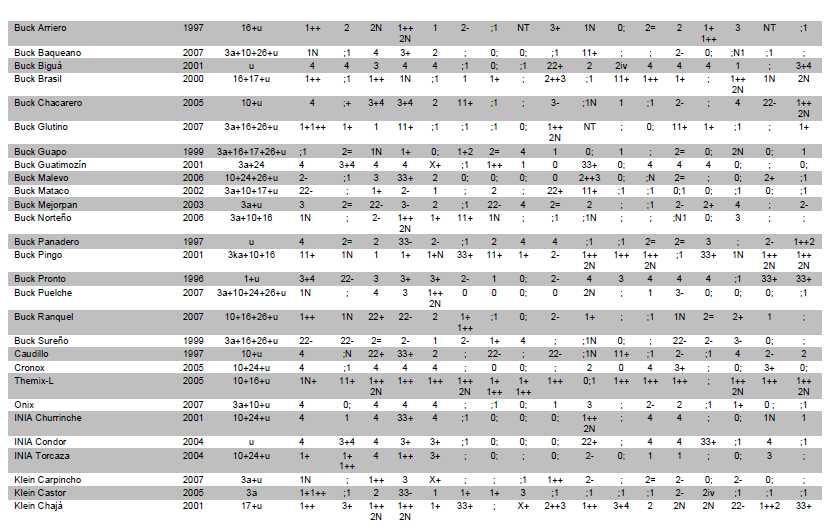
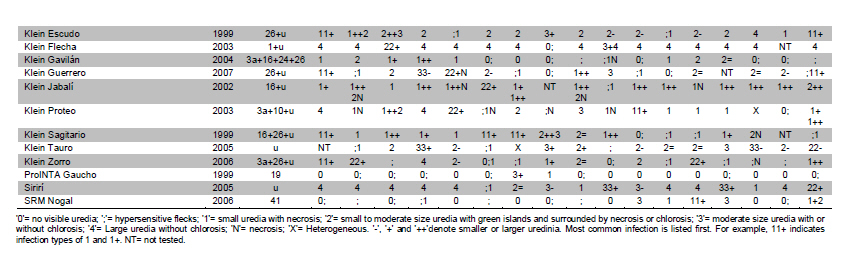
Electronic Journal of Biotechnology, Vol. 14, No. 3, May 15, 2011 Identification of leaf rust resistance genes in selected Argentinean bread wheat cultivars by gene postulation and molecular markers Leonardo Sebastián Vanzetti§1 · Pablo Campos§2 · Melina Demichelis1 · Lucio Andres Lombardo1 · Paola Romina Aurelia1 · Luis María Vaschetto1 · Carlos Tomás Bainotti3 · Marcelo Helguera*1 1Grupo
de Biotecnología y Recursos Genéticos, INTA EEA Marcos Juárez, Córdoba,
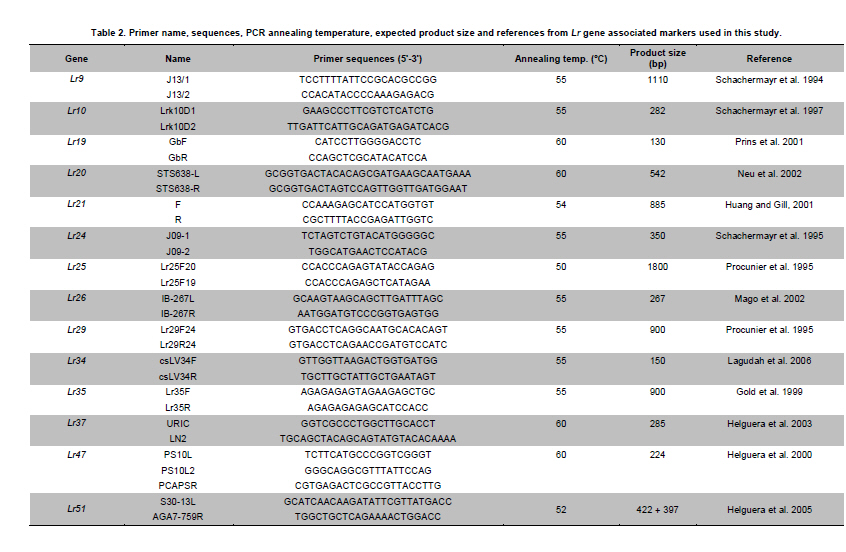
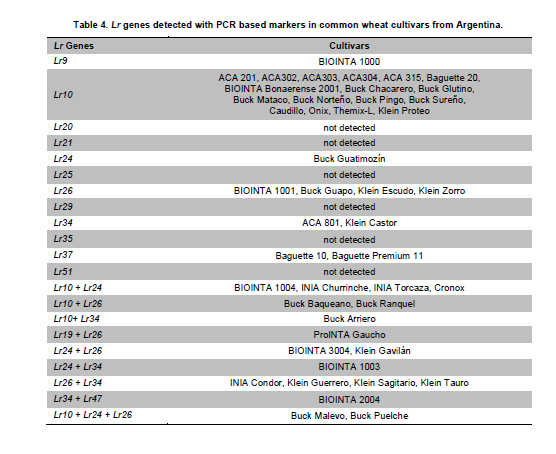
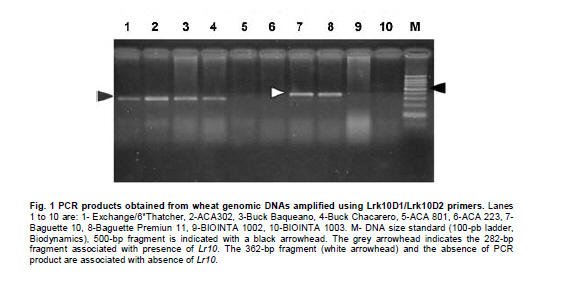
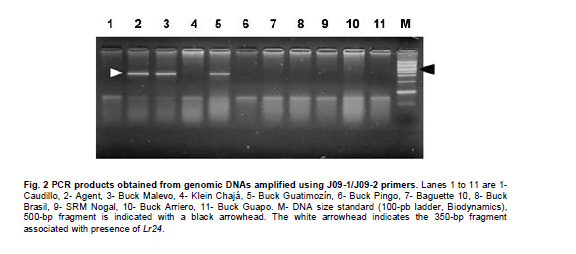
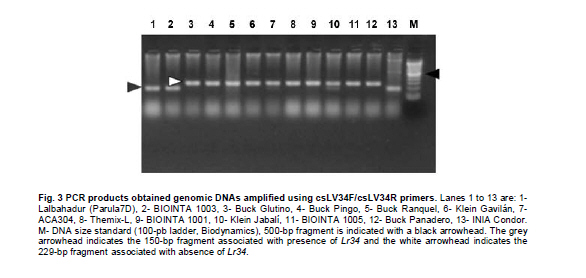
Argentina *Corresponding author: mhelguera@mjuarez.inta.gov.ar §This authors equally contributed to this work. Financial support: Grants from the Government of Argentina, INTA. Recieved: August 11, 2010 Accepted: March 31, 2011 Code Number: ej11032 Abstract Leaf rust, caused by Puccinia triticina Eriks. is a common and widespread disease of bread wheat (Triticum aestivum L.), in Argentina. Host resistance is the most economical, effective and ecologically sustainable method of controlling the disease. Gene postulation helps to determine leaf rust resistance genes (Lr genes) that may be present in a large group of wheat germplasm. Additionally presence of Lr genes can be determined using associated molecular markers. The objective of this study was to identify Lr genes that condition leaf rust resistance in 66 wheat cultivars from Argentina. Twenty four differential lines with individual known leaf rust resistance genes were tested with 17 different pathotypes of leaf rust collected from Argentina. Leaf rust infection types produced on seedling plants of the 66 local cultivars were compared with the infection types produced by the same pathotypes on Lr differentials to postulate which seedling leaf rust genes were present. Presence of Lr9, Lr10, Lr19, Lr20, Lr21, Lr24, Lr25, Lr26, Lr29, Lr34, Lr35, Lr37, Lr47 and Lr51 was also determined using molecular markers. Eleven different Lr genes were postulated in the material: Lr1, Lr3a, Lr3ka, Lr9, Lr10, Lr16, Lr17, Lr19, Lr24, Lr26, Lr41. Presence of Lr21, Lr25, Lr29, and Lr47 could not be determined with the seventeen pathotypes used in the study because all were avirulent to these genes. Eleven cultivars (16.7%) were resistant to all pathotypes used in the study and the remaining 55 (83.3%) showed virulent reaction against one or more local pathotypes. Cultivars with seedling resistance gene combinations including Lr16 or single genes Lr47 (detected with molecular marker), Lr19 and Lr41, showed high levels of resistance against all pathotypes or most of them. On the opposite side, cultivars with seedling resistance genes Lr1, Lr3a, Lr3a + Lr24, Lr10, Lr3a + Lr10, Lr3a + Lr10 + Lr24 showed the highest number of virulent reactions against local pathotypes. Occurrence of adult plant resistance genes Lr34, Lr35 and Lr37 in local germplasm was evaluated using specific molecular markers confirming presence of Lr34 and Lr37. Our data suggest that combinations including seedling resistance genes like Lr16, Lr47, Lr19, Lr41, Lr21, Lr25 and Lr29, with adult plant resistance genes like Lr34, SV2, Lr46 will probably provide durable and effective resistance to leaf rust in the region. Keywords: genetic control of pathogens, molecular markers, plant breeding. IntroductionLeaf rust of bread wheat (Triticum aestivum L.), caused by the fungus Puccinia triticina Eriks. is one of the most common foliar diseases of this crop worldwide (Oelke and Kolmer, 2004; Mebrate et al. 2008). In South America, leaf rust occurs annually in approximately 9 million ha of wheat, causing yield losses of over 50% in severe epidemics and is the primary reason of cultivar replacement. Considering the large areas sown to cultivars that require chemical control in an average epidemic (a very high proportion of the wheat area in the Southern Cone is planted with susceptible or moderately susceptible cultivars), the total annual cost of fungicide applications to control leaf rust in the region is about US$50 million (Germán et al. 2007). The use of resistant cultivars offers the most effective and ecologically sustainable method of control of the disease, therefore, incorporating genetic resistance to this pathogen into adapted germplasm is a major goal in most wheat breeding programs. Plant disease resistance can be classified into two categories: qualitative resistance, conferred by a single resistance gene (also termed as major, seedling, or race specific resistance) and quantitative resistance, mediated by multiple genes or quantitative trait loci (QTLs) (also termed as adult plant, race non-specific or slow rusting resistance) with each providing a partial increase in resistance (Kou and Wang, 2010). More than sixty genes for leaf rust resistance (Lr), most of them major, seedling or race specific genes, have been catalogued to date in wheat (McIntosh et al. 2008; Samsampour et al. 2010). However, the gene-for-gene interaction between host resistance genes and pathogen virulence genes combined by virulence shifts in pathogen populations have reduced the effectiveness of a significant number of major leaf rust resistance genes (Johnson, 2000; Bulos et al. 2006). Replacement of highly variable land races by higher yielding, pure-line varieties in many parts of the world, including the South Cone, has further reduced the wheat gene pool and favored virulence shifts events in pathogen populations. In this context, a better knowledge on the identity of effective Lr genes present in adapted cultivars that can be used as donors of resistance in wheat breeding programs could greatly improve the efficiency of developing resistant cultivars by using these genes per se or by stacking different resistant genes in a given cultivar, a process also known as gene pyramiding (Messmer et al. 2000); thereby helping to avoid the release of cultivars that are genetically uniform (Mebrate et al. 2008). Before gene pyramiding is practiced, it is advisable to identify effective and genetically different sources of resistance. Gene postulation is the most frequent method to determine the presence of the probable race-specific seedling resistance genes (Lr genes) in a host cultivar, many researchers have used this method to for identifying Lr genes in a group of wheat genotypes (Kolmer, 2003; Oelke and Kolmer, 2004; Wamishe and Milus, 2004; Hysing et al. 2006; Mebrate et al. 2008). In this procedure, a host cultivar is evaluated against a collection of Puccinia triticina Eriks. Isolates carrying different avirulence/virulence gene combinations (pathotypes) on the basis of phenotypic expressions in the form of infection types (ITs). Infection types produced on a series of differential genotypes carrying individual Lr resistant genes are the basis for comparing the ITs of wheat cultivars with unidentified genes for leaf rust resistance. Gene postulation can be complicated by interactions between resistance genes and it is better suited for resistance genes that are expressed at seedling stage (qualitative resistance genes) than at adult plant (in general, quantitative resistance genes). Certain adult plant resistance genes like Lr34 and Lr46 are very important for breeding because they proved to confer durable resistance over a long period of time in different environments, as well as against diverse pathotypes of the fungus (Schnurbusch et al. 2004). Alternatively to gene postulation, presence of Lr genes can be determined by testing host cultivars with molecular markers linked to resistance genes. This approach overcomes some of the problems associated with traditional gene postulation, such as gene interactions and plant stage of gene expression. Recently there have been advances in the mapping (and development of molecular markers) of several leaf rust resistance genes (Helguera et al. 2000; Prins et al. 2001; Helguera et al. 2003, Helguera et al. 2005; Gupta et al. 2006; Lagudah et al. 2006; Bansal et al. 2008; Mebrate et al. 2008; Kuraparthy et al. 2009; Sun et al. 2009; Samsampour et al. 2010). Once these genetic factors are mapped, they can be controlled by molecular markers and the corresponding genotypes of individuals can be assessed easily. As a consequence, the identification of cultivars carrying favourable alleles at these loci will provide valuable genetic material for the development of new improved varieties. The objective of this study was to identify Lr genes that condition leaf rust resistance in 66 bread wheat cultivars from Argentina using gene postulation and molecular markers. Materials and Methods A set of 66 bread wheat cultivars registered in Argentina were used to identify Lr genes with molecular markers and/or traditional gene postulation. Seed stocks were obtained from Instituto Nacional de Tecnología Agropecuaria (INTA) Marcos Juárez Wheat Germplasm Bank (Marcos Juárez, Argentina). Sixteen near isogenic lines of Thatcher that differ for single leaf rust resistance genes and eight wheat cultivars and breeding lines with known leaf rust resistance genes were used as controls for molecular markers and/or traditional gene postulation studies (McIntosh et al. 1995, Table 1). Additionally, differential lines carrying adult plant resistance Lr genes ‘VPM1’ (Lr37+), ‘Lalbahadur + Lr34’ (Lr34 has been transferred from Parula7D) and ‘Thatcher*6/RL5711’ (Lr35+) were used as controls for molecular markers. Tests for P. triticina resistance (gene postulation studies) At least eight plants from 66 common wheat cultivars from Argentina and 24 differential lines carrying single Lr genes (Table 1) were evaluated for seedling resistance to leaf rust at the Cereal Disease Laboratory in INTA Bordenave during 2008. Plants were grown in a greenhouse set at 18 to 22ºC with natural day light. Seedlings were inoculated with leaf rust pathotypes MDT 10-20, MFP 20, MFR 10-20, MFT 10-20, MDR 10-20, MCP 10, MCP 10-19, CHT, MBJ 10-20, MFR 10, MRN 10, TDP 10-20, MFP 10, TNR 20, MGT 10, SNG 10 and MLD 10, which are the most common isolates in Argentina. Seedlings were inoculated at 8 days after planting, when the primary leaves were fully extended. For each pathotype, an oil-spore mixture was atomized onto the seedling and seedlings were incubated in a dew chamber for 18 hrs with no light at 18-22ºC. After incubation, the plants were placed in a greenhouse set at 18-22ºC. Infection types were scored (first and second leaf) 14 days after inoculation using a scale of 0-4 according to Stakman et al. (1962). Pathotypes designations were according to the North American nomenclature described in Long and Kolmer (1989) with additional virulence over Lr10 or Lr20 genes. DNA extraction Genomic DNA was extracted from fresh leaves of single plants using a fast, small-scale DNA isolation procedure based on Helguera et al. (2005). Detection of Lr genes with molecular markers Molecular markers for fifteen different Lr genes were included in this study. Primer names, sequences, annealing temperatures, expected product sizes and original references from Lr gene associated markers are detailed in Table 2. In all cases, Polymerase Chain Reaction (PCR) reactions were performed in 25 µl aliquots in a PTC-100 (MJ Research) thermal cycler. The reaction buffer contained 100 ng of genomic DNA (template), 1X Taq polymerase buffer (Promega Corp. Madison, WI), 1.0 U Taq DNA polymerase (Promega), 0.2 mM of each deoxynucleotide, 0.2 µM of each primer and 1.5 mM of MgCl2. Forty cycles of 45 seconds at 94ºC, 45 sec at 55-65ºC depending on the primer combination (Table 2) and 1 min at 72ºC were performed, followed by a final elongation step of 10 min at 72ºC. After that, PCR reactions (10 µl each) were run on 2% agarose (Promega) gel in SB buffer at constant power (100 V) for about 30 min. In the case of markers for Lr37 and Lr51 PCR products (10 µl) were digested with restriction enzymes Dpn II and Pst I, respectively, and later electrophoresed. After electrophoresis the gels were stained with ethidium bromide (0.5 g/L) and visualized with Ultra Violet (UV) light. ResultsSeedling resistance by gene postulation The infection types (ITs) of the 17 P. triticina pathotypes to 24 differential lines are listed in Table 1. Table 3 presents the ITs of wheat cultivars considering the same 17 P. triticina pathotypes used to infect the 24 differential lines. The ITs of the cultivars and the differential lines to the different P. triticina pathotypes determined the gene postulations (Table 3, Column 2). Eleven different Lr genes were postulated in the material: Lr1, Lr3a, Lr3ka, Lr9, Lr10, Lr16, Lr17, Lr19, Lr24, Lr26, Lr41. Presence of Lr21, Lr25, Lr29, and Lr47 could not be determined with the seventeen pathotypes used in the study because all were avirulent to these genes (to date, no virulent pathotypes against Lr21, Lr25, Lr29, and Lr47 have been detected in Argentina). Of the 66 tested cultivars, one cultivar had no detectable seedling resistance (Baguette 21) and 50 cultivars had one or more known Lr genes including 42 cultivars that also had one or more unidentified Lr genes. The fifteen remaining cultivars exhibited ITs patterns not corresponding to the 24 differential lines and/or combinations, and were postulated to carry only unidentified Lr genes (Table 3). Eleven cultivars (16.7%) were resistant to all pathotypes used in the study and the remaining 55 (83.3%) showed virulent reaction against one or more local pathotypes. Between the eleven resistant cultivars, ACA 302, ACA 303, ACA 304, ACA 315, Klein Gavilán, Buck Glutino, Themix-L, Buck Ranquel and Klein Jabalí, were postulated to have Lr16 because they had a low IT of 0-2, but preferently 1 with large uredinia (typical of Lr16), in MDT 10-20, MFP 20, MFR 10-20, MFT 10-20, MDR 10-20, MCP 10, MCP 10-19, MBJ 10-20, MFR 10 TDP 10-20, MFP 10, TNR 20, SNG 10 and MLD 10). Additionally in these cultivars, low ITs in CHT, MRN 10 and MGT 10 were explained by: presence of Lr3a + Lr24 + Lr26 (Klein Gavilán); Lr10 and unknown Lr genes (ACA 302, ACA 303, ACA 304, Themix-L); Lr10 + Lr26 + unknown Lr genes (Buck Ranquel); Lr3a + Lr10 + unknown Lr genes (ACA 315); Lr3a + Lr26 + unknown Lr genes (Buck Glutino); unknown Lr genes (Klein Jabalí) (Table 3). Buck Mataco had also low IT (of ; to 22+) to all pathotypes which was explained by presence of Lr3a + Lr10 + Lr17 associated with additional unknown Lr genes. As mentioned before, Lr genes 21, 25, 29, and Lr47 showed also low infection types to all tested pathotypes, however none of the ten resistant cultivars described before showed the typical low ITs of 0; to ; associated with Lr47 (observed in BIOINTA 2004) or ITs of 2, associated with Lr21, therefore, presence of these Lr genes was discarded. Lr25 and Lr29 showed ITs more similar to previous resistant cultivars (mostly 1, Table 1), therefore, presence of these genes cannot be discarded by gene postulation. In these cases, the use of Lr genes diagnostic markers can be a valuable tool to complement gene postulation data. Procunier et al. (1995) developed PCR markers diagnostic for Lr25 and Lr29 which were used in our study and confirmed that none of the selected local cultivars had these sources of leaf rust resistance (Table 4). In the remaining 56 susceptible cultivars, a high variation in the ITs responses to the 17 pathotypes was observed, defined by the number of virulent reactions per cultivar, which varied between 1 and 17. In general, combination of seedling resistance genes including Lr19, Lr41, Lr9, Lr16, Lr26, Lr10, Lr3a, Lr16 + 10, Lr16 + 26, Lr16 + 26 + 10, Lr16 + Lr17, Lr16 + Lr10 + Lr3a, Lr16 + Lr26 + Lr17 + Lr3a, Lr26 + Lr3a, Lr26 + Lr10 + Lr3a, Lr26 + Lr10 + Lr24, Lr26 + Lr10 + Lr24 + Lr3a, frequently with additional unknown genes, were detected in cultivars with virulent reactions (high ITs) against one, two or three pathotypes and avirulent reactions (low ITs) against complementary pathotypes (for example, Buck Norteño, Klein Sagitario, Buck Brasil, Buck Guapo, BIOINTA 1001, Klein Zorro, BIOINTA 3004, Buck Sureño, Buck Arriero, Buck Pingo, Buck Baqueano, Klein Guerrero, Klein Escudo, Buck Malevo, Buck Puelche) (Table 3a, b, c). In contrast with previous situation, cultivars with a higher number of virulent reactions (high ITs) against local pathotypes (eight or more) were associated with presence of a different set of Lr genes including Lr1, Lr3a, Lr3a + Lr24, Lr10, Lr3a + Lr10, Lr3a + Lr10 + Lr24, also frequently in combination with additional unknown genes (for example, ACA 801, Baguette 10, Klein Flecha, Buck Pronto, BIOINTA 1003, Baguette P. 13, Buck Guatimozin, Baguette 20). Identification of leaf rust resistance genes using molecular markers An important field in which molecular markers are used in wheat resistance breeding is the determination of designated resistance genes in genotypes where the genetic background has not yet been clarified, like most commercial cultivars. Using this approach we screened the set of 66 adapted cultivars previously evaluated by gene postulation for presence of 15 Lr genes (Table 2), and eight genes were detected: six seedling genes (Lr9, Lr10, Lr19, Lr24, Lr26, Lr47) and two adult plant resistance genes (Lr34, Lr37). Genes Lr20, Lr21, Lr25, Lr29, Lr35 (adult plant resistance gene) and Lr51 were not detected in tested cultivars (Table 4). Seedling resistance genes detected in adapted cultivars using molecular markers The diagnostic PCR fragment associated with Lr9, was detected only in BIOINTA 1000 and in the differential line ‘Thatcher*6/Transfer’ (Table 4). Gene postulation studies showed thirteen leaf rust pathotypes with avirulence to Lr9, and only four pathotypes with virulent reaction (SNG 10, MLD 10, MRN 10 and TNR 20, Table 1). Despite its low frequency in local wheat cultivars, the wide range of effectiveness positioned Lr9 as a useful source of resistance when deployed in combination with complementary Lr genes like Lr51, Lr21, etc. Lr9, derived from Triticum umbellulata, has also been detected in low frequency in some European countries and in the USA (Urbanovich et al. 2006; Kolmer et al. 2007a). Lr10 is a T. aestivum L. gene located in the short arm of chromosome 1A. Schachermayr et al. (1997) developed a diagnostic marker amplifying a 282-bp fragment associated with the presence of Lr10 (Figure 1). Using this marker, we detected the diagnostic fragment in 23 local cultivars (34%) (Table 4). Gene postulation studies using ‘Exchange/6*Thatcher’ differential line showed avirulent reaction with MFP 20, CHT and TNR 20 and virulent reaction with the remaining 14 pathotypes (Table 1). This data posicionate Lr10 as of limited usefulness in agriculture because of its narrow range of effectiveness. Lr10 is the first cloned resistance gene in wheat, its sequence encodes a CC-NBS-LRR type of protein with similarities to RPM1 in Arabidopsis thaliana and to resistance gene analogs in rice and barley (Feuillet et al. 2003). The Lr19 gene was introgressed to the hexaploid wheat from Agropyron elongatum, into the wheat substitution line ‘Agrus’ (McIntosh et al. 1995). We detected the Lr19 diagnostic fragment in ‘Thatcher*7/Translocation 4’ differential line and in only one local cultivar (‘ProINTA Gaucho’) between 66 tested cultivars (Table 4). Molecular data was confirmed by gene postulation studies, Lr19 avirulent reaction in 16 pathotypes and virulent reaction in only MCP-10-19 was present in Thatcher*7/Translocation 4’ differential line and in ProINTA Gaucho (Table 1, Table 3). Lr19 in combination with other major leaf rust resistant gene like Lr51, can be an effective source of resistance against leaf rust, however, a special attention must be taken when using this gene in breeding because the alien chromosome segment carrying Lr19 is associated with undesiderable yellow pigment in flour (Zhang et al. 2005). Presence of Lr24 was detected by PCR markers developed by Schachermayr et al. (1995) in ten cultivars between 66 (Figure 2, Table 4). Additional sources of Lr24 with a smaller chromosomal segment that is not detected with the marker used in our study have been reported (Schachermayr et al. 1995; Prabhu et al. 2004), therefore we cannot discard presence of additional cultivars carrying the smaller translocation carrying Lr24. Gene postulation studies showed 7 pathotypes with avirulent reaction and 10 with virulent reaction (Table 1) and positionate Lr24 as a moderately useful source of resistance when deployed in combination with complementary Lr genes like Lr19, Lr21, etc.The gene Lr24, derived from Agropyron elongatum, has also been used in hard red and soft red winter wheats from USA (Kolmer et al. 2007a). The source of the Lr26 resistance is hybrid derivatives of cultivated wheat and rye cv. ‘Petkus’ with a chromosomal translocation 1BL/1RS, which were produced in the 1930s in Germany. The good level of resistance against leaf rust (as well as resistance to stripe and stem rusts and to powdery mildew) that was conferred by this translocation has made it highly popular among breeders. Commercial wheat cultivars with the Lr26 gene were introduced in the 1950s and over 400 wheat cultivars that carry the 1BL/1RS translocation have been produced over the years (Kosman et al. 2004). In Argentina, sixteen cultivars between 66 (24%) showed presence of the 1BL/1RS rye translocation (Table 3). Gene postulation studies including control line ‘ProINTA Pigué’ showed 7 pathotypes with avirulent reaction and 10 with virulent reaction (Table 1). These data would positionate Lr26 as a moderately useful source of resistance if deployed in combination with complementary Lr genes like Lr19, Lr21, etc. First reports of leaf rust pathotypes virulent for Lr26 in Argentina come from early nineties, which agree with observed in Kosman et al. (2004) in many locations around the world, however, it is interesting to highlight the high frequency of recent cultivars carrying 1RS/1BL translocation even though leaf rust resistance is broken. This impact appear to have arisen with the high yield and widespread adaptability frequently found in wheat carrying 1RS/1BL translocation. In line with that argument, the still effective resistance to local isolates of stem rust has also promoted its use in breeding programs. The leaf rust resistance gene Lr47 was transferred from chromosome 7S of Triticum speltoides to chromosome 7A of Triticum aestivum into a interstitial translocation. PCR analysis using a molecular marker associated with Lr47 showed presence of this source of resistance only in ‘BIOINTA 2004’ with no additional cultivars carrying Lr47 detected in the study. In gene postulation studies all tested pathotypes showed avirulent reaction against Lr47 at seedling stage (Table 3, Table 4). This source of resistance has not been widely exploited in breeding; however some Lr47-isolines from commercial cultivars have been recently released in California and Argentina (Brevis et al. 2008; Bainotti et al. 2009). Adult plant resistance Lr genes detected in cultivars using molecular markers The Lr34 gene originates from bread wheat and is located on chromosome arm 7DS. Lr34 has been recently cloned and it codes a protein that resembles an Adenosine Triphosphate (ATP) binding transporter of the pleiotropic drug resistance subfamily (Krattinger et al. 2009). Lagudah et al. (2006) developed a codominant PCR marker distant 0.5 cM of the Lr34 locus that we used in our study to detect this gene and ten cultivars amplified the 150-bp fragment associated with Lr34 (Figure 3, Table 3). This gene has supported resistance to leaf rust in wheat for more than fifty years and is extensively used in breeding programs worldwide (McIntosh et al. 1995; Krattinger et al. 2009). Lr37 is an adult plant leaf rust resistance gene located within a segment of Triticum ventricosum (Taush.) Cess. chromosome 2NS translocated to the short arm of bread wheat chromosome 2AS (McIntosh et al. 1995). In agreement with Bulos et al. (2006), two between 66 tested cultivars showed the 285-bp diagnostic DNA fragment associated to Lr37. ‘Baguette 10’ and ‘Baguette Premium 11’ share European germplasm carrying Lr37 in their genealogy (for example VPM1) suggesting a common origin of Lr37. In contrast with Lr34 and Lr35, Lr37 has been found susceptible against specific pathotypes at adult plant stage (Chicaiza et al. 2006). Moreover, special consideration must be taken with Lr37 in breeding programs because even though presence of virulent pathotypes for this gene have been reported, additional genes of agronomic interest like Yr17, Sr38 and Cre5 are also present into the 2NS T. ventricosum segment carrying Lr37 (Jahier et al. 2001). Discussion One of the main disadvantages in using single gene resistance is that because of rapid changes in predominant rust pathogen races (pathotypes) in nature, single-gene resistance in a cultivar may become ineffective soon after it is released. For example, in USA, Lr9 was initially used in soft red winter wheat in the 1970s and initially gave complete immunity to leaf rust. However, within a few years, races with virulence to Lr9 appeared and soon became widespread in the Easter USA. A similar story suffered Lr18, Lr24 and Lr26. Leaf rust races in US with virulence to Lr41 and Lr50 were identified even before cultivars with genes were released (Kolmer et al. 2007a). In Argentina we have identified races with virulence to Lr19, Lr26 and Lr41 in cultivars that originally were resistant to leaf rust, and races with virulence to Lr51 before a cultivar carrying this gene was released. In this context, stacking two or more effective resistance genes in one background can enhance durability and the level of rust resistance, at least, considering dominant pathotypes in one environment. In line with this argument, Oelke and Kolmer (2004) observed that hard red spring wheat cultivars from USA with seedling resistance genes Lr16 and Lr24 with additional adult plant resistance were highly resistant to leaf rust. Kolmer et al. (2007a) consider that the future development of wheat germplasm with high level of resistance will depend on the ability to select genotypes that have combinations of effective resistance genes such us Lr16 and Lr23 with the adult plant resistance gene Lr34. In our work, nine of the eleven local cultivars with high levels of resistance to all pathotypes showed combinations of the seedling resistance genes that included Lr16, which corroborate a central role of this Lr gene in the development of germplasm with high level of resistance against leaf rust also in Argentina. Kolmer et al. (2007a) suggest a positive interaction of seedling (Lr23) and adult plant (Lr34) Lr genes with Lr16 boosting leaf rust resistance in cultivars carrying this combination of Lr genes. In highly resistant local cultivars carrying Lr16 neither Lr34 nor Lr23 were detected, suggesting a combination of Lr genes different than Lr16 + Lr23 + Lr34 also associated with high level of resistance. In Argentina, several old cultivars that show durable resistance to leaf rust as Sinvalocho MA, Pergamino Gaboto and Buck Manantial have been identified. In Buck Manantial (released in 1964), Altieri et al. (2008) identified seedling leaf rust resistance genes Lr3, Lr16, Lr17, and an adult plant resistance gene named BMP1. Buck Manantial (Rafaela MAG/Buck Quequen) has been used as a source of resistance to wheat leaf rust not only in Argentina, but also in Uruguay, Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) and North America (Kolmer et al. 2007b), and it could be a possible donor of Lr16 in local cultivars evaluated in our study. Cultivars carrying Lr16 like Klein Gavilán (Buck Yapeyu/Klein Cacique) and Klein Jabalí (Klein Orion/Klein Toledo//Klein Cacique) share Klein Cacique (Buck Cimarrón/25348//VEE’S’); and Buck Cimarrón and Buck Manantial share Rafaela MAG, which could be a possible donor of Lr16 in Buck Manantial, Klein Gavilán and Klein Jabalí. A similar situation can be observed in Buck Guapo and Buck Sureño, siblines selected from the cross BCHA//Buck Patacón/CRCO, where Buck Patacón (CAEREN 2.4.2/3/RMAG/BPAM//BAGE/KLPET) also has Rafaela MAG in its pedigree as a possible donor of Lr16. ACA 302, ACA 303, ACA 304 and ACA 315 are siblines originated from the cross Buck Poncho/Buck Ñandú, unrelated by pedigree with Buck Manantial or Rafaela MAG, suggesting a different origin of Lr16 in these materials, probably donated by Buck Poncho based on gene postulation data observed in Barcellos et al. (2008). Unfortunately, the probable origin of Lr16 in Buck Glutino, Themix-L and Buck Ranquel could not be traced because the pedigree of these cultivars is unavailable. The high level of resistance against local pathotypes in BIOINTA 2004 and Buck Mataco was due to combinations of seedling resistance genes not including Lr16, like Lr47 and Lr3a+Lr10+Lr17+u, respectively. Adult plant resistant gene Lr34 has also been detected in BIOINTA 2004 (Bainotti et al. 2009), moreover, it is likely that BIOINTA 2004 also had additional seedling or adult plant resistant gene not detected because of limiting factors in gene postulation (lack of virulent pathotypes to Lr gene isolates to perform the postulation, in this case Lr47, and limited number of Lr gene isolates evaluated), and in the number of Lr genes available with diagnostic markers. Presence of unknown additional seedling resistance genes has also been observed in a high number of local cultivars (64%, Table 3). In the eleven highly resistant cultivars identified in our study, gene postulation analysis showed nine cases with presence of additional unknown seedling resistance genes, suggesting effective combinations of between two and four (or more) seedling Lr genes, probably with additional adult plant Lr genes different from Lr34, Lr35 and Lr37, not present in these cultivars (Table 4). In cultivars postulated to have unknown Lr genes, genetic analysis of segregating populations remains the best method to determine the genetic basis of seedling and adult plant resistance of a cultivar. This approach has been successfully used to isolate novel sources of leaf rust resistance in local germplasm like an adult plant leaf rust resistant gene detected in 3BS chromosome arm of the cultivar Sinvalocho named SV2 (Ingala et al. 2007). Kolmer et al. (2007a) used a similar approach to determine the genetics of leaf rust resistance in three landrace-derived wheat cultivars from Uruguay based on segregation and infection type data and four unknown seedling (2) and adult plant (2) Lr genes were detected. In this context, gene pyramiding of effective Lr genes is probably the faster strategy to develop leaf rust resistant wheat cultivars. Gene pyramiding can be greatly facilitated with associated markers through marker assisted selection programs (MAS), this is particularly true in the field of wheat breeding for leaf rust resistance where PCR-based markers are already available for almost half of the 60 or more designated resistance genes and alleles (Samsampour et al. 2010). For example, MAS has been used successfully to introgress into adapted germplasm from Hungary gene combinations Lr9 + Lr24, Lr9 + Lr25 and Lr9 + Lr29 (Vida et al. 2009). Our gene postulation data showed four seedling Lr genes (Lr21, Lr25, Lr29, Lr47) with high levels of resistance against local leaf rust pathotypes and with molecular markers available for MAS (Table 1, Table 2). Additionally, a second group of seedling Lr genes (Lr2a, Lr2c, Lr9, Lr19, Lr41, Lr51, and particularly, Lr16) showed good levels of resistance against a high number of local pathotypes in such a way that cultivars with combinations of complementary Lr genes could show high levels or resistance against leaf rust. In this group some Lr genes have molecular markers suitable for MAS, like Lr9 (Schachermayr et al. 1994; Gupta et al. 2005), Lr19 (Prins et al. 2001; Gupta et al. 2006), Lr51 (Helguera et al. 2005), Lr16 (McCartney et al. 2005) and Lr41 (Sun et al. 2009). Unfortunately, to our knowledge, no markers suitable for MAS associated with Lr2a or Lr2c have been published to the date. Other seedling resistance Lr genes with molecular markers suitable for MAS that have not been evaluated with local pathotypes but could be useful in the development of highly resistant germplasm based on new combinations of Lr genes are Lr53 (Dadkhodaie et al. 2010), Lr56 (Marais et al. 2010a), Lr57 (Kuraparthy et al. 2009), Lr59 (Marais et al. 2008), Lr62 (Marais et al. 2009), Lr63 (Kolmer et al. 2010) and Lr66 (Marais et al. 2010b). However a special care must be taken when using in MAS the last group of Lr genes, because in all cases they were transferred into wheat in the form of wheat-alien translocations that may have an agronomic penalty because of the linkage drag (Helguera et al. 2003). Finally, a third group of Lr genes which is considered critical in the development of durable and highly resistant germplasm are the adult plant resistance Lr genes also known as slow-rusting, or partial resistance genes (Kou and Wang, 2010). In our study, the identification of adult plant resistant genes Lr34, Lr35 and Lr37 was performed using molecular markers also suitable for MAS. In the case of Lr34, Lagudah et al. (2009) reported gene-specific markers that were not tested in our study, therefore, future analyses should be done with local germplasm using these functional markers in order to confirm presence of Lr34. Additional adult plant resistance Lr genes with associated markers suitable for MAS are SV2 (Ingala et al. 2007), Lr22a (Hiebert et al. 2007), Lr12 (Singh and Bowden, 2010), Lr46 (Mateos-Hernandez et al. 2006), Lr48 (Singh et al. 2010) and Lr67 (Herrera-Foessel et al. 2011). Lowe et al. (2011) consider that with the number of mapped partial (slow rusting) rust resistance genes increasing rapidly as a result of ongoing advances in marker and sequencing technologies, breeding programs needing to select and prioritize genes for deployment confront a fundamental question: which genes or gene combinations are more likely to provide durable protection against these evolving pathogens? These authors argue that a refined classification of partial rust resistance genes is required to start answering this question, one based not merely on disease phenotype but also on gene cloning, molecular functional characterization, and interactions with other host and pathogen proteins. Our phenotypic data would support that ideally, multiple minor, adult plant resistance and major seedling resistance genes should be combined to optimize both the level of resistance and its durability in a wheat cultivar. Using this strategy it will be possible to capitalize the two ways that plants respond to pathogen infection: basal resistance (mediated mainly by minor, adult plant resistance genes) and race-specific resistance (mediated by major, seedling resistance genes). Still, a rigorous evaluation of the agronomic effect of new resistance gene combinations on a host phenotype will be required to discard an eventual decrease in host fitness. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11032f2.jpg] [ej11032t4.jpg] [ej11032t3c.jpg] [ej11032f3.jpg] [ej11032t2.jpg] [ej11032f1.jpg] [ej11032t3.jpg] [ej11032t3b.jpg] [ej11032t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}