 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
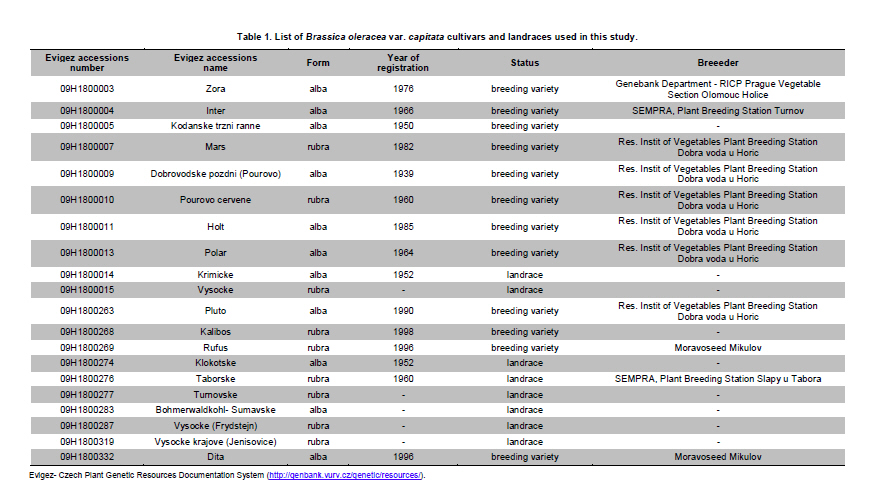
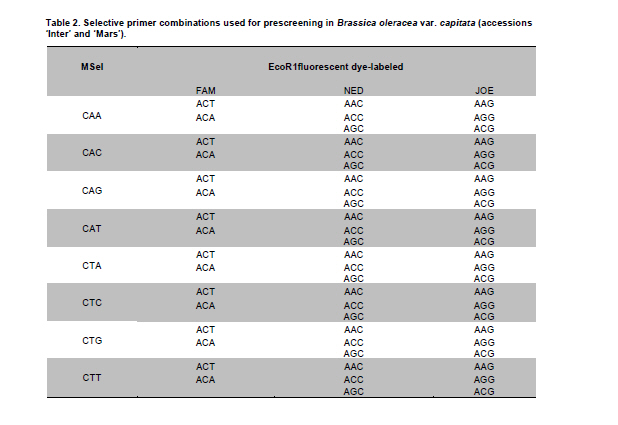
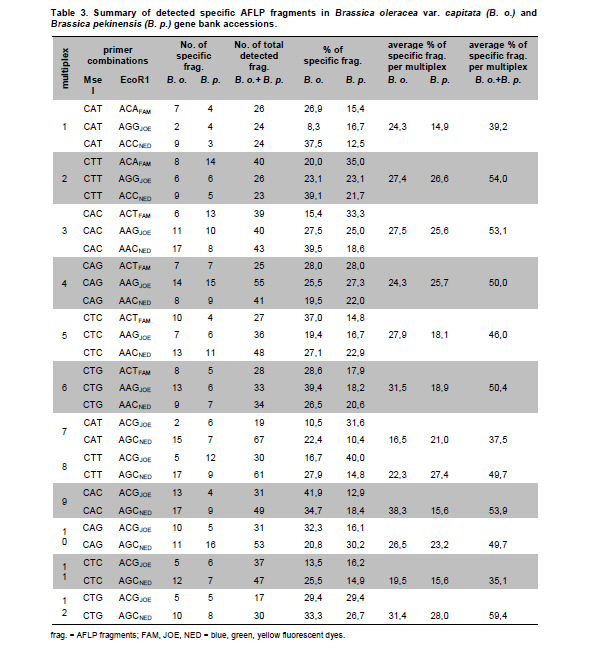
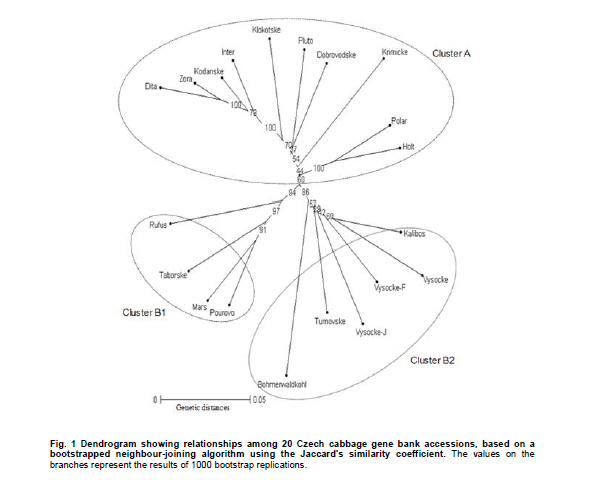
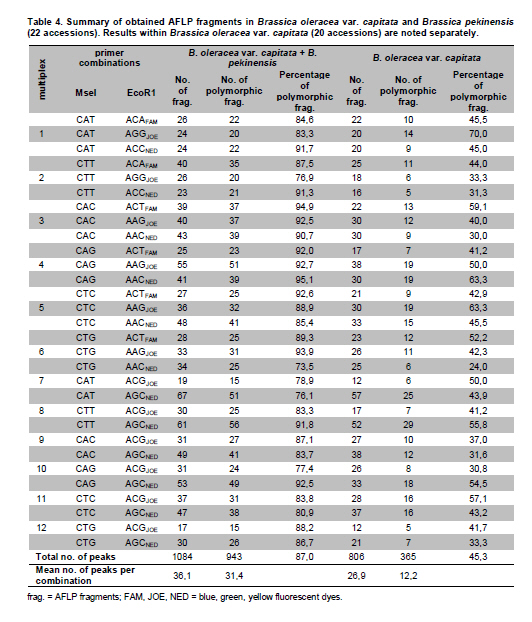
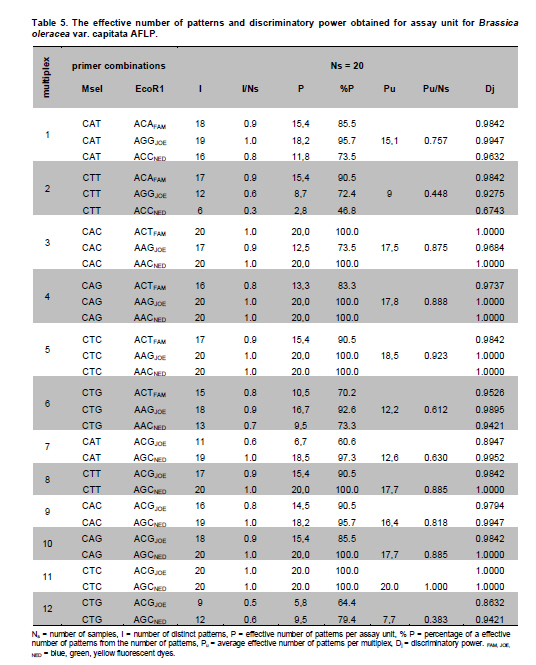
Electronic Journal of Biotechnology, Vol. 14, No. 3, May 15, 2011 Genetic diversity of Brassica oleracea var. capitata gene bank accessions assessed by AFLP Zuzana Faltusová*1 · Ladislav Kučera1 · Jaroslava Ovesná1 1Department of Molecular Biology, Division of Plant Genetics, Breeding and Product Quality, Crop Research Institute, Czech Republic *Corresponding author: faltusova@vurv.cz Financial support: Ministry of Agriculture, Institutional research plan MZE0002700604- Systems for sustainable cultivation of agricultural crop plants and production of high-quality and safe foodstuffs, feedstuffs and raw materials (2009-2013, MZE). Received: August 31, 2010 Accepted: March 4, 2011 Code Number: ej11034 AbstractThe genetic diversity of 20 cabbage (Brassica oleracea var. capitata, including sub.var. alba and rubra) cultivars and landraces from the Gene bank of Crop Research Institute was estimated using amplified fragment length polymorphism (AFLP) marker technology. Two cultivars of Brassica pekinensis (syn. Brassica rapa var. pekinensis) were used as outliers in the study. Thirty AFLP primer combinations produced a total of 1084 fragments. A total of 806 fragments, 364 (45%) of them polymorphic, were found across 20 Brassica oleracea var. capitata accessions. The accessions were clustered into two main groups. Special subgroups, reflecting place of origin, were observed within these groups. Ten selective primer pairs were found to be most informative because each of these uniquely identified all of the accessions used. Furthermore, two accessions of Brassica pekinensis were clearly differentiated from the Brassica oleracea var. capitata accessions. AFLP is an efficient tool for determination of genetic diversity of cabbage gene bank accessions. Keywords: amplified fragment length polymorphism, Brassica oleracea var. capitata, cabbage, discriminatory power, genetic diversity. IntroductionBrassica oleracea L. var. capitata (cabbage) is one of the most important crop plants of the species Brassica oleracea L., which has a long growing tradition in the Czechia. It is an herbaceous, biennial, dicotyledonous flowering plant with leaves forming a characteristic compact head. The cabbage is differentiated into white head cabbage (Brassica oleracea var. capitata sub.var. alba) and red head cabbage (Brassica oleracea var. capitata sub.var. rubra). The influence of cabbage consumption on human health is evident and is, in addition to being a source of vitamins and fibre, connected with secondary metabolites called glucosinolates, which are known to possess anticarcinogenic properties (summarised by Sarikamiş et al. 2009). Some variation in active substances between cultivars has been described. Singh et al. (2006) found variability in antioxidant phytochemicals (ascorbic acid, lutein, β-carotene, DL-α- tocopherol and phenolics) in 18 cabbage cultivars. Variation in terms of glucosinolate profiles and levels were observed in different cabbage genotypes (Sarikamiş et al. 2009; Kabouw et al. 2010). For white cabbage, samples derived from different regions of Europe had some differences in glucosinolate and protein profiles (Kusznierewicz et al. 2008). The plants with high glucosinolate contents will be further utilised as a potential genetic source for breeding. The same importance as identification of differences in active substances has determination of genetic variability between cultivars. The identification of genetic markers that strictly differentiate single cultivars is helpful for effective conservation of plant material in gene banks and for breeders. Only a few studies (Kresovich et al. 1992; Phippen et al. 1997; Van Hintum et al. 2007) have investigated genetic variation within cabbage cultivars. Furthermore, characterisation of diversity in genetic resources and genotype fingerprinting are important for managing gene banks. Different methods were used for this characterization, including the AFLP (Amplified Fragment Length Polymorphism) technique developed by Vos et al. (1995). This method has been successfully used for the assessment of genetic diversity in many plant species (e.g., Zea mays L.- Hartings et al. 2008; Solanum lycopersicum- Van Berloo et al. 2008; Gossypium spp.- Murtaza, 2006; Olea europaea- Sanz-Cortés et al. 2003; Lolium perenne L.- Guthridge et al. 2001). In the case of the genus Brassica, Negi et al. (2004) demonstrated the great value of AFLP markers in the management of gene banks by using them to estimate the genetic diversity of 18 Brassica nigra cultivars. AFLP-based genetic diversity has also been described in Brassica juncea (Srivastava et al. 2001), Brassica carinata (Jiang et al. 2007), Brassica rapa (Zhao et al. 2005; Liu and Meng, 2006; Takuno et al. 2007; Warwick et al. 2008), Brassica campestris (syn. Brassica rapa- Das et al. 1999; Huh and Huh, 2001) and Brassica napus (Čurn et al. 2002; Liu and Meng, 2006). The detection of polymorphisms by AFLP is more efficient than with other marker systems (Das et al. 1999- Brassica campestris L. cultivars, Lombard et al. 2000- Brassica napus L. cultivars). The aim of our study was to find AFLP markers for fingerprinting and estimating genetic diversity within Czech cabbage accessions deposited in the gene bank. Materials and MethodsA group of 20 cabbage accessions (cultivars and landraces) (Brassica oleracea var. capitata) (Table 1) were acquired from Crop Research Institute Prague, Gene bank Olomouc. Young leaves collected from 20 accessions of cabbage seedlings and from 2 cultivars of Brassica pekinensis, ‘Hilton’ and ‘Nozaki’ were homogenised in liquid nitrogen and genomic DNA was isolated using the DNeasy Plant Mini Kit (Qiagen, Germany). The AFLP assay was carried out according to the Perkin-Elmer protocol (Part number 402083, Rev.A, November 1995). In brief, DNA was restriction digested with EcoRI (Fermentas) and MseI (New England Biolabs) and AFLP ligation was carried out using an AFLP Ligation and Preselective Amplification Kit for Regular Plant Genomes (Applied Biosystems). T4 DNA ligase (Fermentas) was used for ligation. Next, 5 µl of the restriction ligation product was visualised on 1.5% agarose and 9 µl of the rest of the ligation mixture was diluted with 91 µl TE0.1. The preselective amplification reaction was prepared using 2 µl of the diluted restriction-ligation DNA prepared earlier, 0.5 µl of the preselective primer pairs and 7.5 µl of AFLP Core mix. A UNO II (Biometra) thermo cycler was used for the PCR. 5 µl of preselective amplification product was visualised on a 1.5% agarose gel and the remaining 5 µl were diluted by 45 µl TE0.1. The selective amplification reaction was prepared as a multiplex PCR in a 10 μl-reaction mixture [0.2 mM dNTP, 1 μM MseI primer, 3x 0.1 μM fluorescently-marked EcoRI primers, 0.5 U Taq polymerase (Qiagen, Germany), 1x buffer with 25 mM MgCl2 and 1 µl diluted preselective amplification product] in the ABI PRISM 7700 thermocycler (Applied Biosystems, USA). Samples were prepared for electrophoresis using 4.8 µl of formamide, 0.5 µl of GS-500 ROX-labelled size standard and 1 µl of selective amplification product. The amplification products were separated by capillary electrophoresis in a Perkin Elmer Genetic Analyser ABI PRISM 310 and analysed using GeneScan™ and Genotyper™ software (Applied Biosystems, USA). AFLP markers were generated with the Selective Amplification Start Up Kit for Regular Plant Genomes (Applied Biosystems, USA). Initially, 64 primer combinations involving 8 MseI and 8 EcoR1 primers grouped in 24 multiplexes (Table 2) were screened for polymorphisms using two different cabbage cultivars, ‘Inter’ (white form) and ‘Mars’ (red form). After the pre-screening, 30 primer combinations (arranged as multiplex groups 1 to 12) involving 8 EcoR1 and 6 MseI primers with the highest number of detectable polymorphic peaks within a size range of 100 to 500 bp were used for analyzing all accessions. Using the Genescan™ and Genotyper™ software (Applied Biosystems), the peaks representing AFLP fragments were exported into Excel as a 1-0 matrix for additional data analysis. DARwin software was employed to determine the level of dissimilarity between accessions, and a dendrogram was constructed based on a bootstrapped neighbour-joining algorithm using the Jaccard's similarity coefficient. The total and mean numbers of distinct AFLP profiles generated by each selective primer combination were obtained. The discriminatory power (Dj = 1-C) and confusion probability (Cj) of the jth assay (Tessier et al. 1999) were calculated according to Cj = ∑Ii= 1 pi (Npi - 1)/N-1, where pi represented the frequency of the ith pattern, N the sample size and I the total number of patterns generated by the jth assay. The confusion probability reflects the probability that two randomly chosen accessions share an identical pattern. The effective number of patterns per assay was given by P= 1/1-DL (Belaj et al. 2003), where DL = lim(Dj) = 1-∑Ii= 1 pi2, as sample size tends toward infinity. ResultsThe 30 primer-AFLP combinations used in this study produced 1084 fragments in total, of which 286 fragments were species-specific for B. oleracea var. capitata (detected in all samples of cabbage) and 231 fragments were present only in B. pekinensis. Most of the species-specific markers (17 fragments) in B. oleracea var. capitata were produced from primer combinations CAC/AAC (from multiplex 3), CTT/AGC (from multiplex 8) and CAC/AGC (from multiplex 9). In B. pekinensis, most of the species-specific markers were produced from primer combinations CAG/AGC (from multiplex 10) (16 fragments), CAG/AAG (from multiplex 4) (15 fragments) and CTT/ACA (from multiplex 2) (14 fragments). For discrimination between B. oleracea var. capitata and B. pekinensis, the most informative primer combinations were multiplexes 12, 9, 2 and 3 (Table 3). Accessions of B. oleracea var. capitata and B. pekinensis were strictly differentiated. Across 20 B. oleracea var. capitata (cabbage) accessions, we detected 806 different fragments, 365 of which were polymorphic (45%). The average number of polymorphic fragments per combination was 12. The most informative primer combinations were CAT/AGG (from multiplex 1), with 70% polymorphic fragments and CAG/AAC (from multiplex 4) and CTC/AAG (from multiplex 5), with 63.3% polymorphic fragments for both of them (Table 3 ). Accessions were clustered into two main groups, with this division correlating the white and red forms of cabbage. Only one of the white cabbages, ‘Bohmerwaldkohl’ was clustered with the red cabbage accessions. Special subgroups were observed within the groups, reflecting the region of plant breeding and pedigree relationships. In cluster A, cultivar ‘Holt’ was bred from cultivar ‘Polar’ (bootstrap value 100); in cluster B1, cultivar ‘Mars’ was bred from cultivar ‘Pourovo cervene’ (bootstrap value 91); and in cluster B2, cultivar ‘Kalibos’ was bred from landrace ‘Vysocke’ (bootstrap value 60) (Figure 1). Finally, we found that all 30 used primer pairs showed high discrimination power for the differentiation of cabbage accessions. Ten of the primer pairs were most informative within cabbage because they showed a discriminatory power value of 1 (Table 5). Each of these 10 primer combinations was able to differentiate 20 cabbage accessions, some of them closely related. The complex of these 10 primer combinations is very effective for assessing genetic diversity within cabbage and was found to be very efficient for gene bank accession fingerprinting. For the multiplex-arranged primer combinations for AFLP, we suggest the use of multiplex 3, 4, 5 or 11. In addition, we detected three markers that would be candidate markers for white cabbage accessions. These markers were not present in tested red cabbage accessions, whereas they were present in most of the white cabbage accessions. Marker CTG/AAG192 was detected in 8 white cabbages accessions, and markers CAG/AAC487 and CAT/AGC160 were detected in 7 white cabbage accessions. DiscussionCharacterisation of diversity in the genetic resources of gene banks is important for managing these gene banks. Among the generally used molecular techniques, AFLP is a powerful technique for cultivar identification and fingerprinting (Powell et al. 1996; Lombard et al. 2000; Garcia et al. 2004). In Brassica species, the most-studied cultivated plants were Brassica rapa (Zhao et al. 2005; Liu and Meng, 2006; Takuno et al. 2007; Warwick et al. 2008) or Brassica napus (Lombard et al. 2000; Čurn et al. 2002; Liu and Meng, 2006) and some varieties of Brassica oleracea (Gu et al. 2008- B. o. var. botrytis), compared with the rare AFLP studies of genetic diversity performed on Brassica oleracea var. capitata (Van Hintum et al. 2007). Compared to other studies, we used extensive sets of selective primers. We pre-screened 64 primer combinations and selected 30 primer combinations with the highest number of detectable polymorphic peaks; 45% of the detected fragments within B. oleracea var. capitata were polymorphic. The rate of polymorphism in AFLP fragments described for Brassica is from 23% (B. carinata- Warwick et al. 2006) to 86.6% (B. rapa- Takuno et al. 2007). The value of polymorphism is dependent on the diversity of the chosen plant material to a certain degree because the high level of polymorphism described by Takuno et al. (2007) was obtained from plants of morphologically distinct Brassica rapa subspecies. Despite this fact, the value of polymorphic fragments in our collection of cabbage is high. Furthermore, 10 selective primer pairs having a discrimination power value of 1 could be effectively used for fingerprinting cabbage accessions. Some of the primers that we selected for cabbage fingerprinting have also been used in other AFLP Brassica studies. The most commonly used selective primer combination, CAC/AAG (Lombard et al. 2000, Srivastava et al. 2001; Lionneton et al. 2002; Liu and Meng, 2006; Takuno et al. 2007), showed a discrimination power of 0.9684. The second most commonly used primers, CTC/ACT (Lombard et al. 2000; Srivastava et al. 2001; Negi et al. 2004; Takuno et al. 2007), showed a discrimination power of 0.9842. Primers CTC/AAG (Lombard et al. 2000; Srivastava et al. 2001; Lionneton et al. 2002; Takuno et al. 2007) and CTC/AAC (Srivastava et al. 2001; Lionneton et al. 2002; Negi et al. 2004; Sabharwal et al. 2004) showed discrimination power values of 1 among cabbage accessions used in our study. The 20 cabbage accessions were divided according to their origin and form. Only the white accession ‘Bohmerwaldkohl’ was clustered with the red cabbages, but the group in which it was clustered are old landraces that originated in mountain regions with similar conditions, which may explain this genetic similarity. AFLP was successfully used for the assessment of genetic diversity within cabbage cultivars and landraces, and we suggest effective primer combinations for cabbage gene bank accession fingerprinting. Furthermore, candidate AFLP markers for white cabbage were identified. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11034f1.jpg] [ej11034t5.jpg] [ej11034t1.jpg] [ej11034t4.jpg] [ej11034t2.jpg] [ej11034t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}