 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
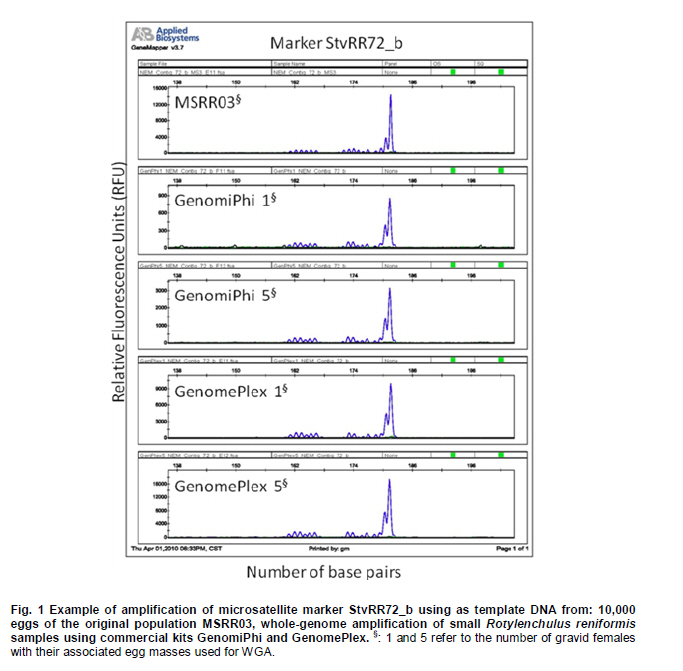
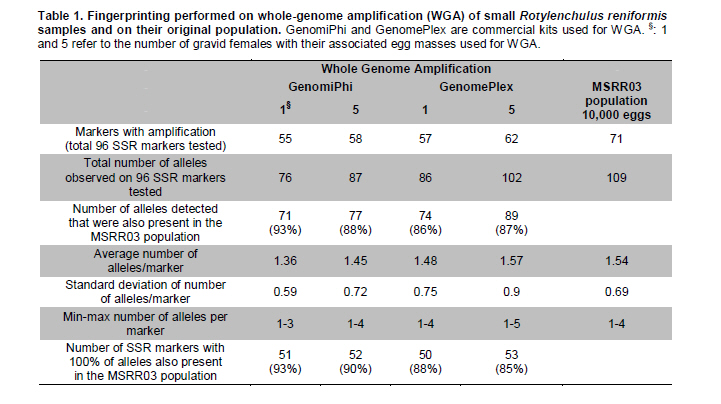
Electronic Journal of Biotechnology, Vol. 14, No. 3, May 15, 2011 Comparison of whole-genome amplifications for microsatellite genotyping of Rotylenchulus reniformis Renée S. Arias*1 · Salliana R. Stetina2 · Brian E. Scheffler1 1USDA-ARS
MSA Genomics and Bioinformatics Research Unit, Stoneville, MS, USA *Corresponding author: Renee.Arias@ARS.USDA.GOV Financial support: This work was supported by USDA-ARS project no. 6402-22000-005-00D, and 6401-21000-001-00D. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. Recieved: July 17th, 2010 Accepted: March 21, 2011 Code Number: ej11037 Abstract Currently, a large number of microsatellites are available for Rotylenchulus reniformis (reniform nematode); however, two barriers exist for genotyping samples from different geographical areas. The limited amount of nucleic acids obtained from single nematodes which would require their multiplication to obtain enough DNA for testing; and the strictly regulated transport of live samples and multiplication in greenhouse for being a plant pathogen. Whole-genome amplification (WGA) of samples consisting of one and five dead gravid females with their associated egg masses was successfully performed on disrupted tissue using three commercial kits. DNA yield after WGA ranged from 0.5 to 8 µg and was used to test 96 microsatellite markers we previously developed for the reniform nematode. The results were compared to those of fingerprinting the original population (MSRR03). Out of 96 markers tested, 71 had amplicons in MSRR03. Using WGA of single gravid females with their associated egg masses, 86-93% of the alleles found on MSRR03 were detected, and 87-88% of the alleles found on MSRR03 when using WGA of samples composed of five gravid females with their associated egg masses as template. Our results indicate that reniform nematode samples as small as a single gravid female with her associated egg mass can be used in WGA and direct testing with microsatellites, giving consistent results when compared to the original population. Keywords: genotyping, reniform nematode, SSR, STR, WGA. Introduction The plant pathogen Rotylenchulus reniformis (Linford & Oliveira), or reniform nematode, has a wide range of plant hosts, affecting a vast number of agriculturally important species in temperate and tropical environments (Robinson et al. 1997). In order to develop breeding programs for plant resistance to pathogens, it is necessary to know their genetic variability (Silva et al. 2008). To analyze the potential genetic variability of reniform nematode across populations, space and time, the protocol developed should take into account the limited size of individuals, the accurate taxonomic identification of the samples, and the strict regulations for the transport and greenhouse multiplication of this pathogen. Microsatellites or simple sequence repeats (SSRs) have become one of the most powerful genetic markers in biology (De Luca et al. 2002). These are short tandem DNA repeats, with 1-bp to 8-bp motifs (Richard et al. 2008), and are ideal markers because of their abundance, high polymorphism, high transferability, high reproducibility, co-dominant inheritance and for being based on polymerase chain reaction (PCR) (Varshney et al. 2005). SSRs have been successfully used on other plant-pathogenic nematodes, i.e., to accurately determine the geographical origin of Globodera in Europe (Plantard et al. 2008) and to effectively determine the type of sexual reproduction in Xiphinema (Villate et al. 2010). Recently, 156 SSR markers developed for R. reniformis allowed detection of genetic variation on isolates from different geographical areas (Arias et al. 2009). However, given the size of the reniform nematode, 0.38-0.52 mm body length in females (Wang, 2001), regular extractions from single individuals cannot yield enough DNA for standard testing of a large number of microsatellites. We have obtained up to 2 micrograms DNA using 10,000 eggs of reniform nematode, but having to collect that many eggs from various geographical locations for population genetics studies would be a limiting factor. On the other hand, transport of live parasitic nematodes within the country or from overseas, as well as their multiplication in greenhouses is subject to strict regulations, and lengthy permit processing. Thus, collecting reniform nematodes from distant areas to further multiply them and then extract their DNA is not a practical approach. The objectives of this work were to explore a protocol that would enable the collection of samples from multiple locations, and study the feasibility of genotyping samples consisting of very small numbers of closely-related nematodes. The protocol had to include the accurate taxonomic identification of the samples, killing the nematodes by freezing to avoid lengthy regulations on pathogen transport and assume the use of regular ice for shipping of samples. Upon arrival, the samples would be subject to whole-genome amplification (WGA) followed by SSR fingerprinting of the amplified DNA. Materials and MethodsThe amphimictic population MSRR03 of R. reniformis was multiplied in greenhouse using tomato (Solanum lycopersicon L. ‘Rutgers’) as host as previously described (Arias et al. 2009). Because positive taxonomic identification of R. reniformis in the field relies on morphology of the adult female, females that had reached reproductive maturity as evidenced by the presence of egg masses were used. To replicate conditions to be used in future experiments, individual gravid females or groups of five gravid females of this population were used accompanied by their egg masses. Gravid females and associated egg masses were removed from root systems by hand, placed in 0.5 mL centrifuge tubes with 100 µL sterile water, and stored in a -80ºC freezer. In forthcoming sample collections, we expect the nematodes will be shipped on regular ice; thus, to mimic those conditions, samples were removed from the freezer and left for 48 hrs on ice before being processed for WGA. Sample processing was done in a sterile hood using filter-pipette tips and sterilized materials. The nematodes and associated egg masses were washed three times with 100 µL sterile water and then transferred to 2 mL Fast-Prep tubes (MP Biomedicals, Irvine, CA) containing one 4-mm-diam stainless-steel bead and two 2-mm-diam glass beads. Two microliters of sterile water were added to the samples and ground in a Fast Prep-24 (MP Biomedicals, Irvine, CA) twice for 20 sec at 4 m/s speed. The tubes were centrifuged for 1 min at 8,000 g, and then 1 µL of supernatant transferred to a clean 0.5 mL tube. Supernatants were placed on ice and immediately processed for WGA. Three commercial kits were used according to manufacturer instructions for genome amplification, REPLI-g Ultra-fast amplification of purified genomic DNA (Qiagen, Valencia, CA), GenomePlex single-cell whole-genome amplification kit WGA4 (Sigma-Aldrich, St. Louis, MO), and GenomiPhi V2 DNA-amplification kit (GE Healthcare Biosciences, Pittsburgh, PA). WGAs were performed twice on samples consisting of one or five gravid females accompanied by their egg masses using each of the kits. Amplified DNA was precipitated and quantified by NanoDrop 1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE). DNA resulting from WGA from all samples was tested with 96 SSR markers previously developed (StvRR_3_a to StvRR_284_a) (Arias et al. 2009). To compare results of SSR amplification from these smaller samples, we used the same DNA extraction from 10,000 eggs of the MSRR03 R. reniformis population performed using DNeasy Plant Maxi Kit (Qiagen, Valencia, CA) that was used by Arias et al. (2009). The sample had been stored in a freezer at -20ºC. Percentage of alleles detected on DNA samples processed by GenomiPhi and GenomePlex kits were transformed to ArcSin of the square root, and the two methods compared by analysis of variance (ANOVA) using STATISTIX (Analytical Software, Tallahassee, FL). ResultsThe three methods of WGA utilized resulted in amplified DNA from R. reniformis. Quantitation of WGA products obtained from samples consisting of one or five gravid females with their associated egg masses using NanoDrop after precipitation were 0.6 and 0.8 μg DNA for REPLI-g, 1.1 and 4.0 μg DNA for GenomiPhi, and 6.0 and 8.0 µg DNA for GenomePlex, respectively. The total reaction volumes for the three methods by following manufacturer’s instructions were 20 μL for REPLI-g, 20 μL for GenomiPhi, and 75 μL for GenomePlex. In our current protocol for fingerprinting, testing 96 SSR markers requires 1 µg DNA. Since less than 1 µg DNA was obtained from the samples twice with REPLI-g, only the amplifications obtained with GenomiPhi and GenomePlex were used to test 96 SSRs. As example of SSR amplification marker StvRR72_b is shown using DNA of the original population MSRR03, and using WGA of one and five gravid females with their associated egg masses by GenomiPhi and GenomePlex commercial kits (Figure 1). The results of running the 96 SSR markers on these four samples and on DNA from the MSRR03 population using 10,000 eggs are shown in Table 1. The number of alleles observed after WGA using GenomiPhi and GenomePlex compared to the alleles present in the original MSRR03 population ranged from 86 to 93%, without significant difference between the two methods (p ≤ 0.27). In a previous experiment where we used 156 SSR markers to compare reniform nematode populations of geographically distant origin (Arias et al. 2009), the percentage of alleles shared with MSRR03 were 36.5, 47.9 and 40.2% for populations from Texas, Georgia and Louisiana, respectively, data not shown. We did not expect to detect all the alleles present in the 10,000 egg population in the smaller samples. All alleles observed after WGA of samples consisting of one and five gravid females with their associated egg masses had been observed in the multiple-egg population, indicating that both methods of WGA did not generate artefacts by SSR amplification. For 50 to 53 markers, the alleles detected after WGA were exactly the same found in the MSRR03 population, regardless of sample size. In several instances, alleles observed after WGA had low fluorescence signal in the MSRR03 population, indicating that WGA of samples comprised of fewer nematodes could allow detection of alleles that probably have low frequency within the population. DiscussionFor molecular work involving phytopathogenic nematodes, one limiting factor is their small size. Although the use of a pool of individuals could yield enough DNA to test large number of markers, this would limit the analysis of intraspecific variation (Skantar and Carta, 2005). Here we show the successful WGA performed on previously frozen R. reniformis gravid females with their associated egg masses and their effective microsatellite fingerprinting using amplified DNA. WGA has been proved a suitable solution when the small size of samples limits their use for extensive molecular work. WGA by multiple displacement amplification has been used on nematodes, such as the human pathogen Brugia malayi (McNulty et al. 2008) which has a body length of 1.0 to 1.7 mm at the larval stage that occurs inside mosquitoes (Eberhard and Lowrie, 1985), and the plant pathogenic Meloidogyne spp. (Skantar and Carta, 2005) which have a body length of 0.4 to 1.9 mm (Eisenback et al. 1994). In the present work we tested three commercial WGA kits, two based on multiple displacement amplification (GenomiPhi and REPLI-g) and one on fragmentation and library construction (GenomePlex); the template in all cases was disrupted tissue from nematodes instead of purified DNA. We have shown that WGA of small samples of R. reniformis with subsequent testing of SSR markers gave results consistent with those observed on the original population of the nematode. From 96 SSR markers previously developed for R. reniformis (Arias et al. 2009) and tested here, markers that amplified DNA from samples consisting of one or five gravid females with their associated egg masses after WGA were subsets of the same 71 markers that amplified DNA of the original population, indicating that no artefacts were generated by the process of WGA. While the DNA in our samples probably originated predominantly from the female parent(s) (i.e., female somatic tissue plus DNA in the fertilized eggs), between 86 and 93% of the alleles detected in samples consisting of one gravid female with her associated egg mass, and between 87 and 88% of the alleles detected in samples consisting of five gravid females with their associated egg masses after WGA by GenomePlex and GenomiPhi respectively, matched the alleles detected in the original population. We did not expect to detect 100% of the alleles of the original population when analyzing samples based on fewer nematodes as the allele frequency could vary given the small sample size or the limited contribution from the male parents in the sample. However the high proportion of alleles detected in the samples show this is a robust method for genotyping gravid females and their associated eggs to get a decent picture of the genetic constitution of small samples. Previous experiments, using 156 SSR markers showed that only 37 to 48% of the alleles present in MSRR03 were present in R. reniformis samples from geographically distant areas. Several advantages have been pointed out regarding the WGA of phytopathogenic nematodes, such as the possibility to store large amounts of DNA of “type specimens”, generation of a set of DNA standards, and the feasibility to exchange material from different locations without regulation restrictions (Skantar and Carta, 2005). As the present work was intended to find a robust, non-labour intensive method for processing large number of samples, we used disrupted tissue as template for WGA instead of purified DNA. In conclusion, it was feasible to perform WGA on R. reniformis samples as small as one gravid female with her associated egg mass, with the SSR markers tested on the amplified DNA giving consistent results with those obtained from DNA of the original population. This approach could be used as a tool to understand the spatio-temporal genetic variation of R. reniformis among and within populations. AcknowledgementsWe thank Ms. Sheron A. Simpson for testing the SSR markers. Any library/sequence information requirements can be addressed to Dr. Brian E. Scheffler at brian.scheffler@ars.usda.gov. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11037t1.jpg] [ej11037f1.jpg] |
| |||||||||

{kind=link}
{kind=link}