 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 4, July 15, 2011 RESEARCH ARTICLE Removal of heavy metals by exopolymeric substances produced by resistant purple nonsulfur bacteria isolated from contaminated shrimp ponds Saijai Panwichian1 · Duangporn Kantachote*1,3 · Banjong Wittayaweerasak2,3 · Megharaj Mallavarapu4 1Department
of Microbiology, Faculty of Science, Prince of Songkla University, Hat Yai,
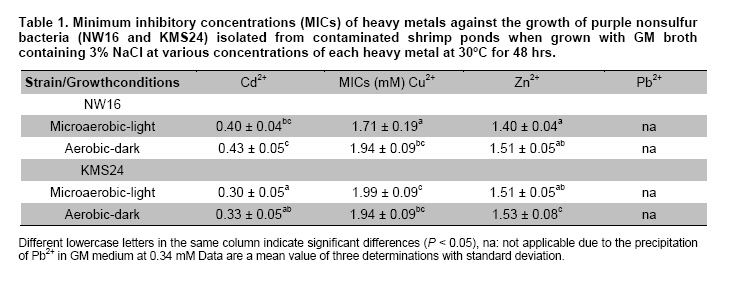
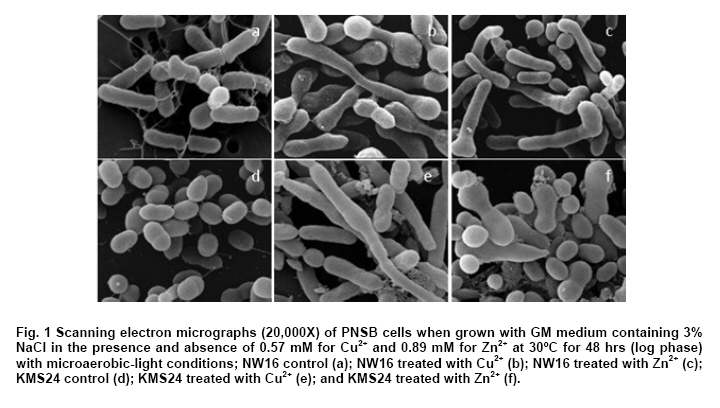
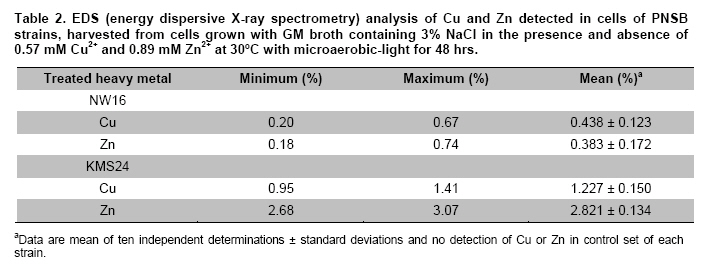
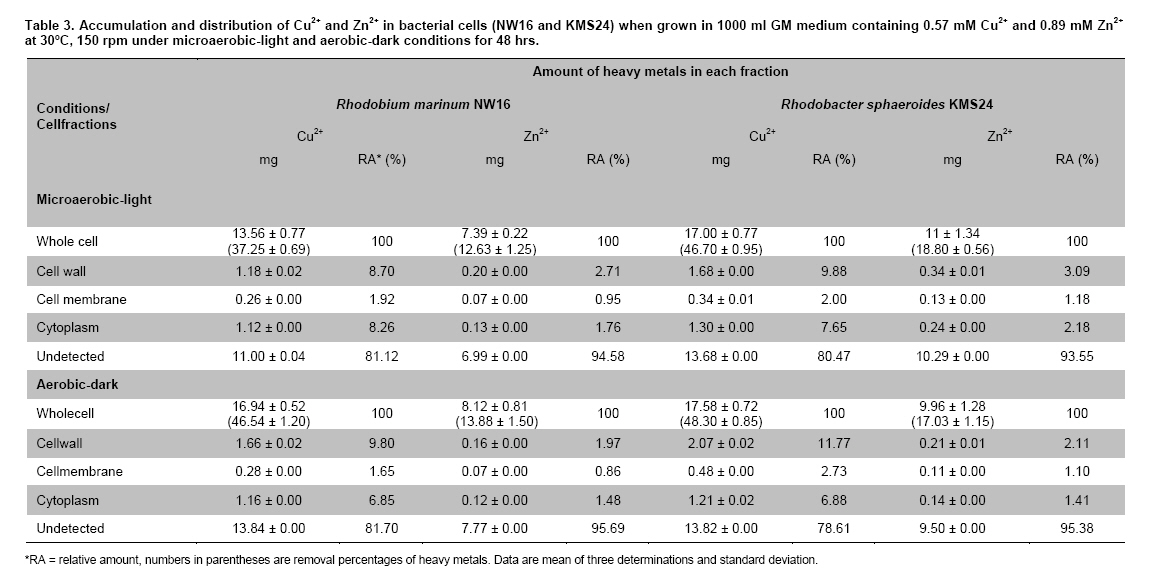
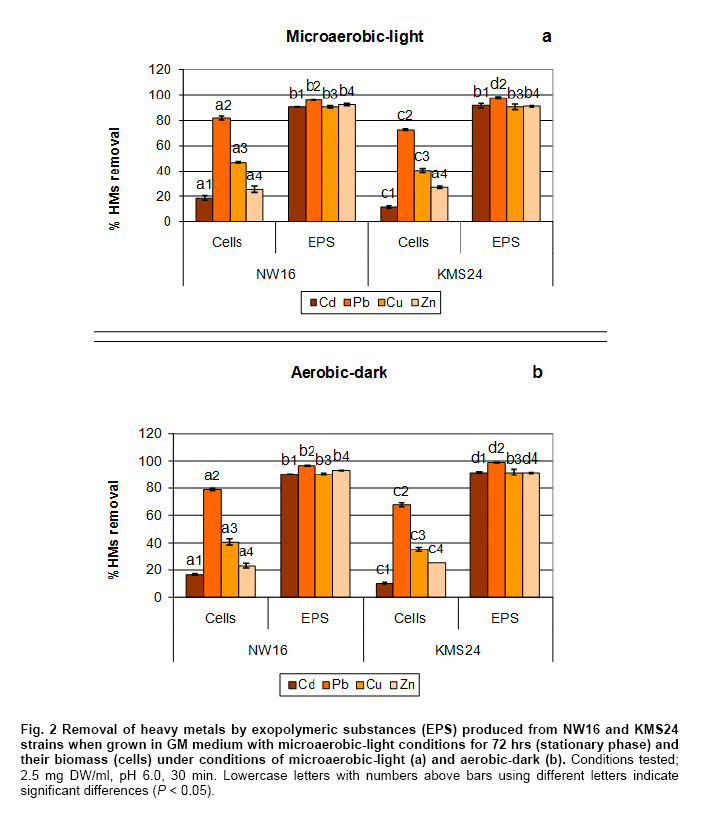
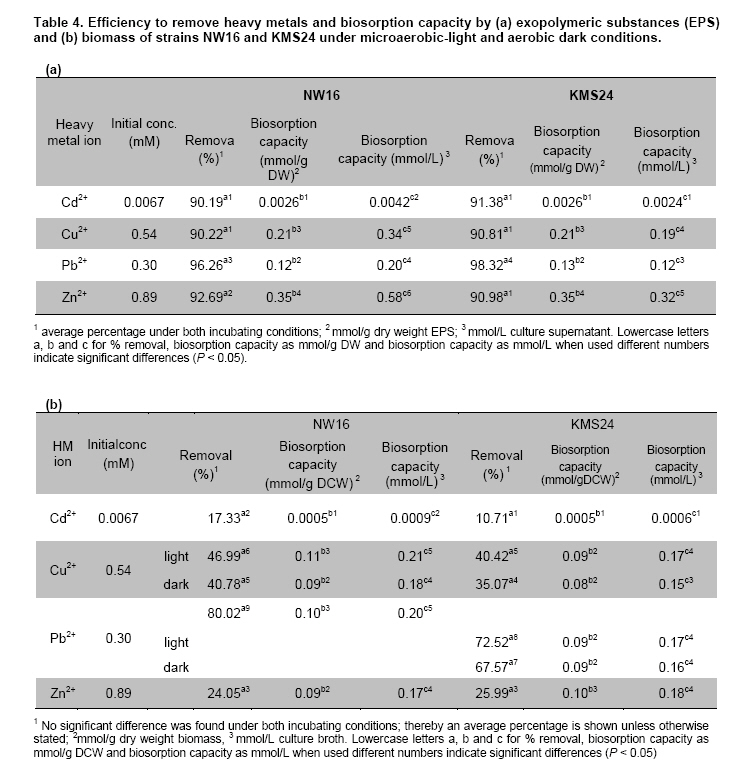
Thailand *Corresponding author: duangporn.k@psu.ac.th Financial support: This work was granted by Office of the Higher Education Commission, Saijai Panwichian was supported by CHE PhD. Scholarship. This work was also supported by a project number SCI520001S, Price of Songkla University Code Number: ej11039 AbstractTwo purple nonsulfur bacteria (PNSB) strains, Rhodobium marinum NW16 and Rhodobacter sphaeroides KMS24 were investigated for their potential to remove heavy metals (HMs) from contaminated shrimp pond water. Tolerance of both PNSB strains growing with both microaerobic-light and aerobic-dark conditions, based on their minimum inhibitory concentrations, was in the order of Cu2+ > Zn2+ > Cd2+ (Pb precipitation occurred at 0.34 mM). Results from a scanning electron microscope equipped with an energy dispersive X-ray spectrometer (SEM-EDX) indicated that Cu2+ and Zn2+ altered the cellular morphology of both strains and accumulated HMs were found in their cells. The highest amounts of both cations were found in their cell walls followed by the cytoplasm and cell membrane. Using the highest concentrations (mM) of HMs found in shrimp pond of 0.0067 Cd2+, 0.54 Cu2+, 0.30 Pb2+, 0.89 Zn2+ and 3% NaCl under both incubating conditions exopolymeric substances (EPS) produced by both strains showed a greater removal of all HMs (average percentages; 90.52-97.29) than their cells (average percentages; 14.02-75.03). Keywords: contamination, exopolymeric substances, heavy metals, photosynthetic bacteria, shrimp farming. IntroductionThe rapid increases in industrialization have not been accompanied by increases in the treatment and disposal of the hazardous wastes they produce and this has resulted in huge environmental problems. The bulk of the hazardous wastes generated have been dumped into the ocean, rivers, canals, any drainage systems and landfills (Zarcinas et al. 2004). In addition, many chemicals used as additives to benefit agricultural activities such as lime, fertilizers, manures, herbicides, fungicides and irrigation waters are also sources of heavy metals (HMs) (Haq et al. 2003). In Thailand shrimp farming is a major economic aquaculture business that can cause severe ecological problems including introducing toxic substances like HMs to the water sources used for shrimp cultivation (Cheung and Wong, 2006; Panwichian et al. 2010) and a wide variety of chemicals and biological products (Gräslund and Bengtsson, 2001). Among these contaminants, HMs can cause the most serious environmental problems for wildlife, and in particular, human health. The HMs can accumulate in the shrimp body and finally affect humans as the highest tropic level of the food chain. Therefore, standard guidelines for aquaculture products have been established for food safety around the world (Petroczi and Naughton, 2009). This means that if the concentrations of the HMs such as Cd (cadmium), Cu (copper), Pb (lead), Zn (zinc) and etc. in shrimp exceed the standard safety levels, they might produce adverse effects on people who consume them and thus have an impact on shrimp exports. Moreover, since there are no strict regulations by governmental agencies for the discharge of effluent from shrimp ponds into the environments, their wastewater is discharged into canals and can flow directly into cultivated areas. This has had a serious polluting effect on soil and agricultural water, especially for the rice-fields and adjacent aquaculture areas. The accumulation of HMs in agricultural soils is therefore of increasing concern due to food safety issues and potential health risk as well as its detrimental effects on the soil ecosystem. Several studies have found that HMs influence microorganisms by harmfully affecting their growth, morphology, and biochemical activities, resulting in a decrease of biomass and diversity (Gad El-Rab et al. 2006). However, microbes living in contaminated environments often become adapted to survive in the presence of existing contaminants using a variety of mechanisms that enables them to tolerate the presence of HMs. For example, extracellular polymeric substances or exopolymeric substances (EPS) secreted by microbes have been recommended as surface active agents (Iyer et al. 2005; Pandi et al. 2009). Because microbes have important roles in the management and sustenance of environments it is important to characterize bacteria present in areas contaminated with HMs and to explore their roles in resistance to HMs that could include their removal (Mengoni et al. 2001). For instance, purple non sulfur bacteria (PNSB) have been reported to resist various HMs (Moore and Kaplan, 1992; Panwichian et al. 2010) and they are normally found in water habitats including shrimp ponds subject to a good light source. Hence, PNSB also have been extensively studied for their bioremediation potential (Watanabe et al. 2003; Feng et al. 2007; Bai et al. 2008). In our previous work we isolated two PNSB strains from HMs contaminated shrimp ponds that were resistant to Cd, Cu, Pb and Zn in salt (NaCl) concentrations of up to 3% (Panwichian et al. 2010). It therefore seemed useful to further explore their HMs resistances and establish if they could be considered for use in bioremediation of HMs in shrimp ponds. Hence, the aims of this study were to investigate the degree of resistance to HMs of both selected PNSB strains and accumulation of HMs in their cells. Moreover, biosorption by EPS and biomass of these PNSB were also investigated for consideration of their use for bioremediation in shrimp ponds. Materials and MethodsHeavy metal resistant purple nonsulfur bacteria PNSB strains; Rhodomium marinum NW16 and Rhodobacter sphaeroides KMS24, were previously isolated from samples of water and sediment collected from HMs contaminated shrimp ponds in Nakhon Si Thammarat and Songkhla Provinces, Thailand, respectively (our unpublished data). Both strains were selected on the basis of their resistance to the HMs tested at the highest concentrations that had been detected in the sediment of shrimp ponds in mM; 0.0067 Cd2+, 0.54 Cu2+, 0.30 Pb2+, and 0.89 Zn2+, and including NaCl up to 3% under both microaerobic-light and aerobic-dark conditions (Panwichian et al. 2010). Determination of minimum inhibitory concentration (MIC) The HMs tested for their minimum inhibitory concentration (MIC) values were Cd, Pb, Cu, and Zn that were prepared from CdCl2, PbCl2, CuCl2 and ZnCl2, respectively. Stocks of the HMs were prepared in deionized water (DI) and sterilized by a filter membrane with a pore size of 0.22 µm and stored at 4ºC until used. Each HM solution was added into GM broth containing 3% NaCl at different concentrations ranging from 0-150 mg/L (1.33 mM Cd, 2.36 mM Cu, 0.72 mM Pb and 2.29 mM Zn). GM medium consists of 3.8 g sodium L-glutamic acid, 2.7 g DL-malic acid, 2.0 g yeast extract, 0.5 g KH2PO4, 0.5 g K2HPO4, 0.8 g (NH4)2HPO4, 0.2 g MgSO4.7H2O, 0.053 g CaCl2.2H2O, 0.001 g nicotinic acid, 0.001 g thiamine hydrochloride, 0.01 g biotin, 0.012 g MnSO4.5H2O, 0.025 g ferric citrate, 0.95 g CoCl2.6H2O, and deionized water up to 1000 ml with a final pH of 6.8. Both bacterial strains were separately grown in GM medium with either microaerobic-light (3000 lux) or aerobic-dark conditions at 30ºC for 48 hrs (log phase) according to the methods previously described by Panwichian et al. (2010). Both incubating conditions were utilized because both PNSB strains will be expected to treat water in normal shrimp ponds after harvesting. Each culture broth was adjusted to an optical density of 0.5 at a wavelength of 660 nm (OD660nm) and 10% of each culture was inoculated into GM broth containing each HM and 3% NaCl as previously described. All culture test tubes were incubated at 30ºC with either microaerobic-light or aerobic-dark conditions as mentioned above for 48 hrs. Bacterial growth was measured as turbidity using a spectrophotometer at a wavelength of 660 nm and the lowest concentration of each HM that prevented growth of each strain is reported as the MIC. Effect of cations (Cu2+ and Zn2+) on the cellular morphology of PNSB In order to observe if the uptake of HMs by cells would alter their cell morphology and to confirm their content of HMs in the biomass, SEM-EDX analysis was performed. Among the HMs (Cd, Cu, Pb, and Zn) that were present as contaminants in shrimp ponds, but only Cu2+ and Zn2+were considered to study their effect on the cellular morphology of selected PNSB strains due to our finding that the amounts of both these HMs (Cu2+ and Zn2+) in the water of shrimp ponds were present in amounts that exceeded the standard guidelines for marine aquatic animal cultivation (Panwichian et al. 2010). The stock cultures of strains NW16 and KMS24 were subcultured twice to obtain active cultures and adjusted to 0.5 OD660nm. Then 10% of each diluted culture was inoculated into GM broth containing 3% NaCl in the presence and absence (control) of 0.57 mM for Cu2+or 0.89 mM for Zn2+(the maximum amounts of both ions detected in sediments of shrimp ponds, CuCl2 and ZnCl2 were used) (Panwichian et al. 2010). All culture tubes were incubated at 30ºC for 48 hrs with microaerobic-light conditions because in these conditions these bacteria were more sensitive to HMs when compared with aerobic-dark conditions (Table 1). After that each culture broth was centrifuged at 8000 rpm for 15 min and the cell pellet was washed twice with 0.1% peptone water prior to use for studying cell morphology by scanning electron microscope (SEM) (JSM-5800LV, JEOL) that was attached to an energy dispersive X-ray spectrometer (EDS) (Oxford ISIS 300). The bacterial fixation and preparation technique used for SEM and EDS were those described in the instruction manuals for the instruments. In brief, bacterial cells were fixed with 2% glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.0) for 1 hr and then washed with the same buffer for 3 times. Cells were then treated with 1% osmium tetraoxide for 90 min followed by washing with 0.1M sodium phosphate buffer and finally dehydration in a series of ethanol (70-100%) for 15 min in each step. The samples were coated with gold and observed with an SEM equipped with the EDS. Accumulation and distribution of cations (Cu2+ and Zn2+) in cells The HMs tested for accumulation in cells was Cu and Zn ions at their previously stated concentrations of 0.57 and 0.89 mM, respectively. The HMs uptake was tested by culturing each isolate in 1000 ml GM medium containing Cu2+and Zn2+with a shaker speed of 150 rpm at 30ºC under microaerobic-light and aerobic-dark conditions for 48 hrs and the conditions set as previously mentioned. Each culture broth was centrifuged at 8,000 rpm for 15 min to obtain the cell pellet and culture supernatant. Each culture supernatant was analyzed for the remaining amount of HM in the solution using Inductively coupled plasma-optical emission spectroscopy (ICP-OES) and all the cell pellet of each organism was analyzed for the distribution of accumulated Cu2+ and Zn2+ within its cells using the method according to Al-Momani et al. (2007) as follows: Cell wall. The accumulation of Cu and Zn ions at the cell wall surface was determined by washing the cell pellet of each strain with 10 ml of 0.1 M sodium citrate for 10 min, 3 times to release the cations from the cell wall and the wash solution of each organism was analyzed for Cu2+ and Zn2+ using ICP-OES. Membrane. After washing, the cell pellets from the previous section they were separately incubated at 30ºC for 1 hr in a solution of 4 ml lysozyme (1 mg/ml) and 6 ml of 0.01 M sodium phosphate, pH 7.0 to hydrolyze the cell walls under the hypertonic conditions of 10.3% sucrose for protecting the protoplast from rupture. After incubating, each suspension was centrifuged at 8000 rpm for 10 min and the concentrations of Cu2+ and Zn2+ in the supernatant were analyzed by ICP-OES. Cytoplasm. Intracellular accumulation (cytoplasm) was determined by using 10 ml of nitric acid (35% HNO3: DI = 1:1) for opening the protoplast and then Cu2+ and Zn2+ were measured by ICP-OES. The percentage accumulation or relative amount (RA) in each cellular part was calculated based on the initial loss of HMs from the culture medium supernatant (accumulated amount in the bacterial cells). Removal of heavy metals by exopolymeric substances and cells of PNSB PNSB strains; NW16 and KMS24 were grown in 1000 ml GM medium with microaerobic-light conditions until the cells reached stationary phase (72 hrs). This incubating condition was used because it allowed for a better growth than with aerobic-dark conditions (Panwichian et al. 2010). Each culture was centrifuged at 8,000 rpm for 15 min to obtain the culture supernatant for detecting EPS while the cell pellets (biomass) were separated to use for measurements for removal of HMs. To obtain the EPS, two volumes of cold ethanol (4ºC) was added to each culture supernatant and incubated at 4ºC, for 24 hrs to precipitate the EPS. The suspension was then centrifuged at 8,000 rpm for 15 min to separate a wet sediment (EPS) and a portion of EPS was used to determine the dry weight (DW). For the biosorption experiment, an amount of wet EPS equivalent to 2.5 mg DW/ml was added to the HMs aqueous solution (0.0067 mM Cd2+; 0.54 mM Cu2+, 0.30 mM Pb2+; and 0.89 mM Zn2+) and the biosorption test conditions were (pH 6, 30ºC, 30 min with microaerobic-light or aerobic-dark conditions) as described in our previous paper (Panwichian et al. 2010). The both incubating conditions were tested because the majority of extracellular polymeric substances (EPS) consist of exopolysaccharides and these compounds are also important in transparent exopolymeric particles (TEP) (Passow, 2002). It is well recognized that both EPS and TEP is secreted by aquatic or marine microbes and the compounds, particularly TEP are degraded by solar radiation especially in the UVB range (Ortega-Retuerta et al. 2009; Hassler et al. 2011). Besides, one of the factors that affect iron binding involves the redox potentials (Rose et al. 2005) thereby there would be competition with other HMs such as Cu and Zn in the shrimp ponds. After biosorption any EPS-metal complex was separated by adding one volume of cold ethanol to the solution (modified from Prado Acosta et. al. 2005) and centrifuging at 8,000 rpm for 15 min. Residual HM levels in the supernatant were determined by ICP-OES and the loss of HMs was interpreted as the amount that had formed a complex with EPS. In addition, the results of the EPS biosorption experiments were compared with the data from the cell pellets (biomass) which had produced the EPS for removal of the HMs. Conditions for removal of the HMs by the biomass were the same as those used for the EPS experiment in order to compare the results directly. The biosorption capacity (mmol HM uptake/g DW) of EPS or biomass was also determined as described by Panwichian et al. 2010. In addition, the yield of EPS per biomass (Yp/b) based on the DW was calculated. Statistical analysis All experiments in this work were carried out in triplicate unless otherwise stated. Data are presented as a mean with a standard deviation from three determinations. One way ANOVA was used to analyze statistical differences at a P-value < 0.05 and mean comparisons were performed by the Duncan's multiple range test. ResultsDetermination of MIC values The testing of the HM tolerance of NW16 and KMS24 strains in the presence of 3% NaCl was carried out with both incubating conditions; microaerobic-light and aerobic-dark (Table 1). With both incubating conditions both strains were resistant to Cu2+ > Zn2+ > Cd2+. The degree of resistance for strain NW16 to Cu2+ and strain KMS24 to Zn2+ with aerobic-dark conditions was higher than with microaerobic-light conditions. Moreover, strain NW16 under both incubating conditions had a significantly higher resistance to Cd2+ (0.40-0.43 mM) than strain KMS24 (0.30-0.33 mM). In contrast, under microaerobic-light conditions strain KMS24 had a significantly higher resistance to Cu2+ (1.99 mM) than that found for strain NW16 (1.71 mM). Overall, the strain KMS24 under both incubating conditions was more resistant to Cu2+ and Zn2+ than the strain NW16. The MIC value for Pb2+ could not be determined in this study due to the precipitation of lead ions in the GM medium when the concentration of Pb2+ was 0.34 mM. However, both selected strains under both incubating conditions did grow in the GM medium containing 0.31 mM Pb2+. SEM was used to examine the morphology of the PNSB cells after growing in GM medium containing 3% NaCl with and without 0.57 mM Cu2+ or 0.89 mM Zn2+ under microaerobic-light conditions for 48 hrs (Figure 1). Comparisons between the cell morphology in the controls (no HM) (Figure 1a and Figure 1d) and cells grown with either Cu or Zn, clearly show marked differences. In Figure 1b and Figure 1c strain NW16 was now elongated or even filamentous in the presence ions of Cu or Zn. Strain KMS24 also showed filamentous cells in the presence of Cu2+ (Figure 1e), but with Zn2+ they were dumbbell shaped (Figure 1f). Moreover, analysis for EDS confirmed that it was present after growth with Cu2+ or Zn2+ but not in the control set. From the ten areas of treatment sets that were selected for EDS analysis, the averages amounts of Cu and Zn in the NW16 cells grown in their presence were 0.44% and 0.38% of the total element components found in the cells, respectively. In contrast, strain KMS24 had even higher levels of Cu and Zn with 1.23% and 2.82% of the total element components, respectively (Table 2). Accumulation and distribution of cations (Cu2+ and Zn2+) in cells Table 3 shows that Zn and Cu ions were accumulated by strains NW16 and KMS24 in three cellular fractions; cell wall, plasmamembrane (cell membrane) and cytoplasm. Over the 48 hrs of cultivation in GM medium, the removal percentages of HMs in 3% NaCl under microaerobic-light and aerobic-dark conditions by the strain NW16 were 37.25 and 46.54% for Cu2+ and 12.63% and 13.88% for Zn2+, respectively. The amount of each HM removed, calculated as a percentage of the initial HM available, was assumed to have been accumulated within the cells. The amount of HMs detected in the various cell fractions was then calculated as a percentage of the amount thought to be associated with the cell (relative amount, RA). For both strains the RA associated with each of the three cell fractions was similar in the order of cell wall > cytoplasm > cell membrane. For instance, uptake of Cu by strain NW16 as a percent RA in the fractions of cell wall, cytoplasm and cell membrane was 8.70, 8.26 and 1.92% (see details for Zn and strain KMS24 in Table 3). Furthermore, the amount of HMs not detected in any fraction of the biomass was extremely high and similar for both strains: undetected Cu (78.61-81.70%) and undetected Zn (93.55-95.69%). Removal of heavy metals by exopolymeric substances and cells of PNSB Comparisons between the ability of the biomass and EPS of the two PNSB strains (NW16 and KMS24) for removal of each HM from the mixed HMs solution (Cd2+, Cu2+, Pb2+ and Zn2+) in 3% NaCl are shown in Figure 2. The EPS from both strains exhibited a significantly higher ability to removal HMs (P < 0.05) when compared to their biomass. The efficiency to remove HMs ions by the EPS produced by two PNSB strains under both incubating conditions had the following order of removal of average percentages: Pb > Zn ≈ Cd ≈ Cu (97.29, 91.84, 90.79 and 90.52%, respectively), while the biomass had an order of removal average percentage of Pb > Cu > Zn > Cd (75.03, 40.81, 25.02 and 14.02%). Results show that the removal average percentages of Cd, Zn, Cu and Pb ions by EPS were increased by about 76.76, 66.81, 49.71 and 22.26%, respectively. There were no significant differences found for the removal efficiency of HMs ions by EPS collected from either strain NW16 or strain KMS24 under both incubating conditions (Figure 2). Therefore removal percentages of HMs ions by EPS under both incubating conditions are presented as average percentages for each HM ion (Table 4a). In contrast, the efficiency of removal of HMs by the biomass of both strains in microaerobic-light conditions was significantly higher than with the aerobic-dark conditions for the case of Cu2+ by the strain NW16 and for both Cu2+ and Pb2+ by strain KMS24 (Table 4b). However, the biosorption capacity of the amount of EPS per liter of supernatant produced by the strain NW16 was higher than for the strain KMS24 but not for the biosorption capacity based on mmol/g DW (Table 4a). Hence, the overall biosorptive capacity was higher for strain NW16 than that found for KMS24 and this is mainly due to the yield (Yp/b) of EPS produced by the strains NW16 and KMS24 that were 0.859 (1.630/1.898) and 0.512 (0.922/1.7996), respectively. Moreover, based on the removal percentages, the biosorption capacity (mmol/g dried cell weight (DCW) or mmol/L) by biomass of the strain NW16 was significantly better for removing all HMs ions with the exception for Zn than that found for the strain KMS24 (Figure 2 and Table 4b). DiscussionResistance and accumulation of heavy metals by PNSB strains The aim of this study was quite different from other previous studies on PNSB strains as the MIC values for HMs of isolates NW16 and KMS24 were investigated in a medium containing 3% NaCl due to the expectation that they might be used for bioremediation of contaminated shrimp ponds. It was of interest that the MIC value for each HM under the both incubating conditions tested was significantly higher than the maximum concentrations of the HM levels found in the shrimp ponds (Table 1 and Panwichian et al. 2010). Hence, these organisms could be used as inoculants for removal of HMs in shrimp farm ponds based on metal immobilization to PNSB cells and/or binding with biofilms that occurred with both PNSB strains and after that biomass/biofilms with HMs will be separated from the water. It was not surprising that the selected strains were the most sensitive to Cd, followed by Zn and Cu (Table 1) and these results are consistent with those previously obtained (Panwichian et al. 2010). Cu is an active component of enzymes such as cytochrome c oxidase including other oxygenases and thus bacteria can tolerate a higher concentration than for the other HMs tested. Zn is also an essential trace element for organisms and is a co-factor of some enzymes and forms complexes with enzymes like RNA and DNA polymerases (Nies, 1999). Therefore, both HMs at higher concentrations were less toxic than Cd. Cd is a toxic element and there have been no reports of its possible biological function. Consequently, it may not be surprising that Cd was the most toxic element to both organisms. Results indicate that Cd was more toxic than Pb for the organisms tested because they could grow at 0.31 mM Pb2+ while the MIC values for Cd2+ for both organisms were between 0.30-0.43 mM under both incubating conditions. Based on results in Table 1, the strain KMS24 seemed to have a higher resistance to Cu and Zn ions than strain NW16 and this was supported by finding higher levels of metal ions either Cu or Zn in cells of the former organism (Table 2). On the other hand, a lower accumulation of Cu or Zn in cells of the strain NW16 might cause by a higher adsorption of metal ions to EPS that was released by the culture. This was due to the EPS produced by the strain NW16 which was higher than that the strain KMS24 as previously stated. However, overall of the 2 strains studied the strain KMS24 is perhaps the best candidate to use for removal of HMs in shrimp ponds. For the reasons previously described we have studied only the accumulation of Cu2+ and Zn2+ and their distribution in cellular fractions. At least 2 steps are required for metal uptake first their absorption onto the cell surface (passive) using metal-binding groups such as carboxyl, phosphate, hydroxyl, etc. followed by active uptake using energy (Wang and Chen, 2006). In the latter step metal ions penetrate the cell membranes and enter into the cells. The present study reveals that the uptake of both metal ions by the selected strains consisted of both passive and active transport as the accumulated metal ions bound on the surface cell were eluted by 0.1 M sodium citrate but were also detected in the cell membrane and cytoplasm (Table 3). Under both incubating conditions tested most of the metal ions accumulated by the selected strains were observed in their cell wall followed by their cytoplasm and the least was found in the cell membrane (Table 3). However, the amount of both metal ions undetected was extremely high (Cu, 11-13.84 mg and Zn, 6.99-10.29 mg for 1 L of each culture broth and this may be because some were consumed by cells for their biological functions. This was possible when considering the composition of the bacterial growth media that normally contain mineral salts similar to those concentrations for supporting growth. Another reason should be possible that some loss of HMs may bind with EPS in the culture supernatant of each strain so they were not accumulated in cells as the evidence in the next experiment. Unfortunately, we did not investigate this point in this experiment. It is known that both Cu and Zn are essential trace elements for bacterial growth and can be detected and accumulated in various subcellular compartments of cells (Table 3). However, when grown at high concentrations of HMs such as those found in contaminated shrimp ponds (0.57 mM Cu2+ and 0.89 mM Zn2+) they produced adverse effects on bacterial cells as filamentous or dumbbell shapes were observed (Figure 1b, 1c, Figure 1e and Figure 1f). A similar result was previously reported by (Gad El-Rab et al. 2006) who showed that Cd altered the cellular morphology of Rhodobacter capsulatus B10 from rods to filamentous cells. As the excess metal ions in cells, results in alterations to cell morphology, selected strains might become resistant by trying to expel metal ions using efflux mechanisms. This may be one reason why the distribution of metal ions was in order of cell wall > cytoplasm > cell membrane. Biosorption of metal ions on the cell surface, including bound EPS might reflect intracellular uptake and/or their efflux mechanisms required for their HMs resistance mechanisms. In the present study results indicate that the main resistance mechanism of the 2 selected isolates involves biosorption of HMs via EPS (Table 4a). Removal of heavy metals by exopolymeric substances and PNSB cells It is well recognized that one of the important roles of EPS produced by microbes is to bind with HMs to allow for growth at high concentrations of HMs and thus EPS has been studied for use as a biosorbent for removing HMs (Watanabe et al. 2003; Iyer et al. 2005). There are two types of EPS: those attached to cells and those present in their culture fluid or non attached fractions (Xu et al. 2009). EPS is often only loosely bound to cells in the form of slimes and can be easily shed into the medium. In this study the EPS used was in the form of slime as it was collected from the culture supernatant using cold ethanol for precipitation. In general, most EPS consists of polysaccharides, protein, RNA and inorganic moieties such as sulfate or phosphate (Iyer et al. 2005; Xu et al. 2009). Based on the compositions of EPS, biosorption of HMs by EPS is non-metabolic, and no energy is required, as they bind with HMs using their negative charges (Watanabe et al. 2003). Hence, the different incubating conditions used, either microaerobic-light or aerobic-dark had no affect on the efficiency of HMs removal (Figure 2 and Table 4a). This means that light from the incandescent lamps had no effect on the degradation of EPS used when compared with sunlight that has UVB wavelengths as previously mentioned. Also there was no effect of the redox processes observed because DI water was used in this experiment and also a washing step completely removed iron from the cell pellets. It is of interest that the EPS produced by both PNSB strains without optimizing their HMs removal conditions are very effective in removing all the HMs tested. Both strains gave no significant differences for biosorption capacity based on mmol/g DW; however, the strain NW16 gave a better biosorption capacity for HMs in mmol/L of culture supernatant (Table 4a) because it produced a higher yield than the strain KMS24 of roughly 1.68 times greater. It should be noted that EPS produced by both PNSB strains occurred at a higher efficiency for removing HMs when compared to the EPS produced by Enterobacter cloaceae, a marine bacterium that removed only 65% of Cd and 20% of Cu (Iyer et al. 2005). In addition, EPS produced by both PNSB strains was more efficient at adsorb all HMs tested in the presence of 3% NaCl when compared with other previous studies. These had shown that their EPS had a high affinity for Pb but a low affinity for Cd, Co, Cu, Ni and Zn (Iyer et al. 2005; Morillo Perez et al. 2008). This is most likely due to differences in their chemical composition, which has not been investigated. EPS produced by both organisms bound HMs in the order of Zn2+ > Cu2+ > Pb2+ and, Cd2+ (Table 4a). One possible reason to explain may be concerned with the different initial concentration of each HM ion and to the possibility that the uptake rate of the metal ion increases along with its increasing initial concentration when the amount of biosorbent is kept unchanged (Wang and Chen, 2006). Therefore, the highest biosorption of Zn2+ by EPS was due to its higher initial concentration. However, both strains produced a higher efficiency for removal of Pb2+ than Zn2+ and Cu2+ which had a higher initial concentration. A lower removal of the latter metal ions could be explained with the fact that the EPS had a limited number of active sites, which would have become saturated above a certain concentration for both HMs ions. Another factor that could have an influence on the amounts of different HMs bound is the possibility that the EPS had different affinities for different HMs. This means that Cd may have a higher affinity for EPS than did Cu and Zn and this was supported by Iyer et al. (2005) as previously mentioned. It will be of considerable interest in the future to compare the chemical composition of the EPS produced by the PNSB strains and other bacteria such as Enterobacter cloaceae. In contrast the efficiency of the biomass of cells from both strains (based on removal percentages) to remove HMs under both incubating conditions was in the order of Pb > Cu > Zn > Cd (Table 4b). This indicates that some functional groups (i.e. carboxyl, phosphate, hydroxyl, amino, etc.) present on the cell surface may have different binding abilities to those of EPS. These different affinities might also explain why Zn produced greater morphological changes to the organisms than did Cu (Figure 1 and Panwichian et al. 2010). In the present study, the removal efficiency of HMs by both strains was significantly higher than found in our previous work (Table 4b and Panwichian et al. 2010) probably because a higher biomass was used with an increase from 0.625 mg DCW/ml to 2.5 mg DCW/ml. However, the biomass of these organisms was less efficient than the EPS in removing HMs. Factors that can affect the efficiency of the biomass to remove HMs should be further investigated and optimized. Concluding Remarks Two selected strains, NW16 and KMS24, isolated from shrimp ponds contaminated with HMs grew well in medium containing HMs ions at concentrations that were much higher than those found in the contaminated shrimp ponds and their resistance to HMs probably includes biosorption, metal uptake and efflux. One benefit of using these organisms is that both produced significant amounts of EPS that was responsible for their major ability to accumulating HMs and protect themselves. Hence, the performance of both PNSB strains indicated that they would be potential candidates for bioremediation processes in shrimp ponds. Acknowledgments We would like to thank the Graduate School for partial support to conduct this research and Dr. Brian Hodgson for checking the English References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11039t1.jpg] [ej11039f2.jpg] [ej11039t2.jpg] [ej11039t4.jpg] [ej11039t3.jpg] [ej11039f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}