 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 4, July 15, 2011 RESEARCH ARTICLE Codon optimization of 1,3-propanediol oxidoreductase expression in Escherichia coli and enzymatic properties Wei Li2 · I-Son Ng1 · Baishan Fang*1 · Jincong Yu2 · Guangya Zhang3 1Department
of Chemical and Biochemical Engineering, College of Chemistry and Chemical
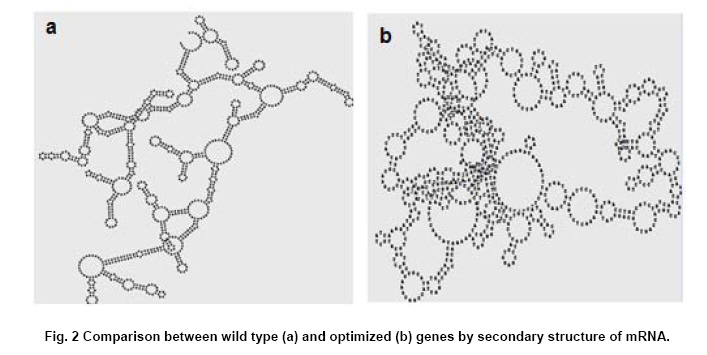
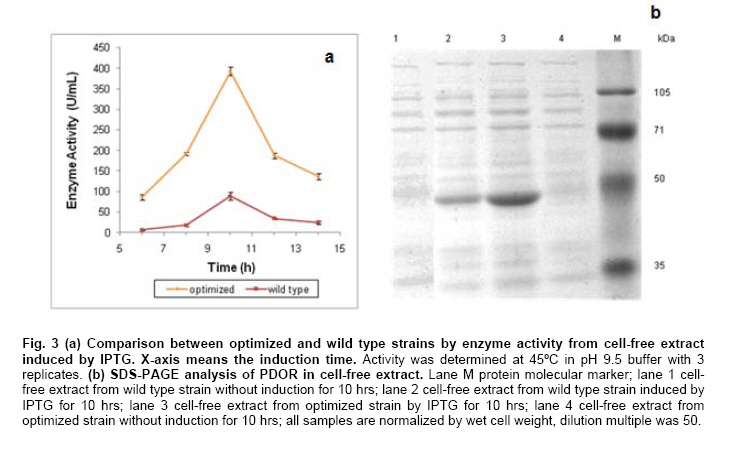
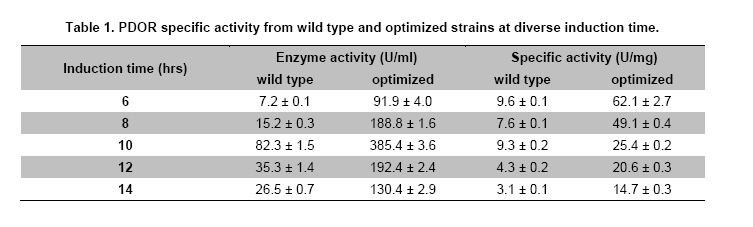
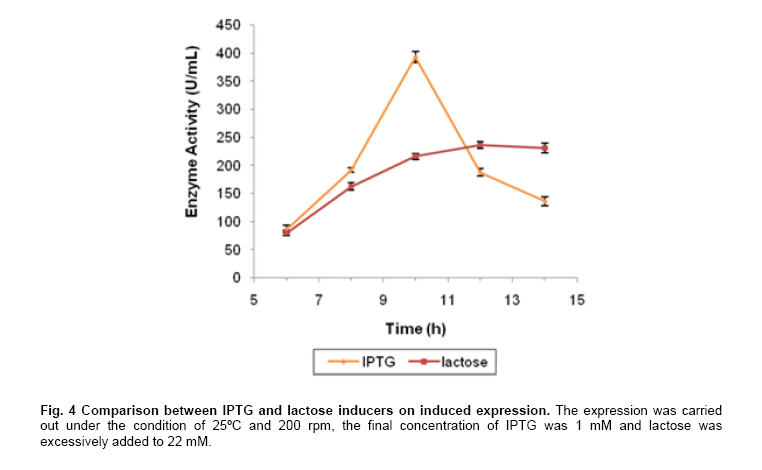
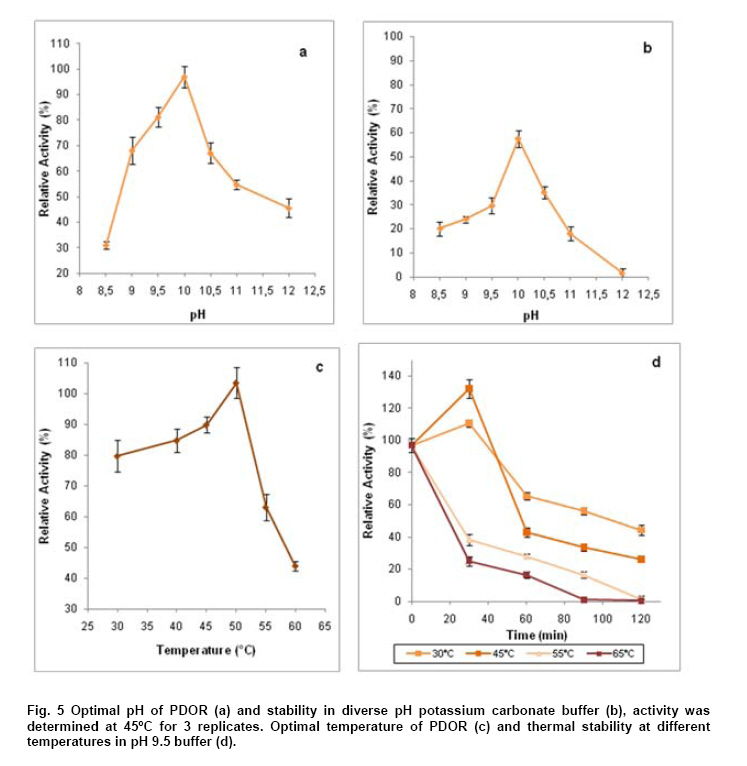
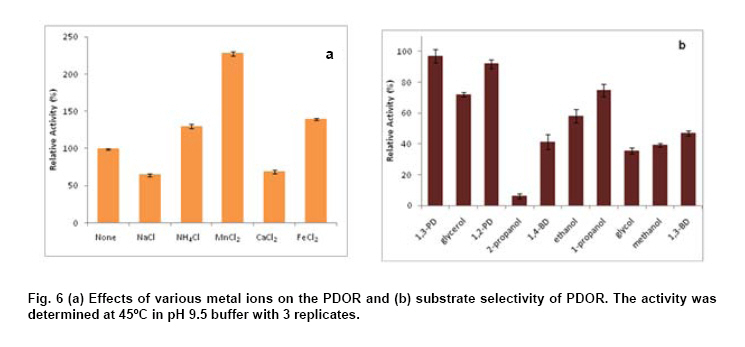
Engineering, Xiamen University, Xiamen, P. R. of China Financial support: This work was supported by Chinese 863 Program (No. 2006AA020103) and National Natural Science Foundation of China (No. 30770059 and 21076172). Code Number: ej11044 AbstractThe gene dhaT from Klebsiella pneumoniae encoding 1,3-propanediol oxidoreductase (PDOR) was de novo synthesized by splicing overlap extension polymerase chain reaction (SOE-PCR) primarily according to Escherichia coli’s codon usage, as well as mRNA secondary structure. After optimization, Codon Adaptation Index (CAI) value was improved from 0.75 to 0.83, meanwhile energy of mRNA secondary structure was increased from -400.1 to -86.8 kcal/mol. This synthetic DNA was under control by phage T7 promoter in the expression vector pET-15b and transformed into the E. coli BL21 (DE3) strain. Inducers such as isopropyl β-D-thiogalactoside (IPTG) and lactose were compared by activity at different inducing time. The activity of PDOR after codon optimized was 385.4 ± 3.6 U/mL, which was almost 5-fold higher than wild type (82.3 ± 1.5 U/ml) under the flask culture at 25ºC for 10 hrs. Then his-tagged enzyme was separated by using Ni-IDA column. The favorite environment for enzyme activity was at 5°C and pH 10.0, PDOR showed a certainly stability in potassium carbonate buffer for 2 hrs at diverse temperatures, enzyme activity was significantly improved by Mn2+. Keywords: 1,3-propanediol oxidoreductase, codon optimization, enzymatic properties, Escherichia coli, overlap extension PCR. Introduction1,3-Propanediol (1,3-PD), a bulk chemical, can be formulated into a variety of industrial products especially in the synthesis of polytrimethylene terephthalate and other polyester fibers (Yuanyuan et al. 2004). It can be produced either by chemical synthesis or by microbial fermentation. In the microbial part, many kinds of microorganisms are proved to be capable of producing 1,3-PD such as Klebsiella pneumoniae (Yuanyuan et al. 2004), Citrobacter freundii (Daniel et al. 1995) and Clostridium butyricum (Raynaud et al. 2003). In the model organism, Klebsiella pneumonia, the glycerol metabolism primarily involves two branch pathways: the reductive branch and the oxidative branch. 1,3-propanediol oxidoreductase (PDOR, E.C. 1.1.1.202) encoded by dhaT gene is the key enzyme in oxidative procedure, which catalyzes 3-hydroxypropionaldehyde (3-HPA), production of reductive branch into 1,3-PD under the consumption of reducing power NADH2 (Fenghuan et al. 2005). The crystallographic structure of PDOR, a type III Fe-NAD-dependent alcohol dehydrogenase, has been determined to be a decameric structure with exactly the same monomer (Marçal et al. 2009). 3-HPA is an inhibitory intermediary metabolite, rapid conversion of 3-HPA to 1,3-PD is one of the ways to improve the yield of 1,3-PD from glycerol and to avoid 3-HPA accumulation, which is tightly associated with the PDOR amount and activity. In order to decrease 3-HPA accumulation during 1,3-PD production, Hao et al. 2008 over expressed dhaT gene in K. pneumoniae TUAC01 and alleviated the toxicity of 3-HPA in fermentation. In the fed-batch fermentation of the recombinant K. pneumoniae strain, over-expressing PDOR individually enhanced the molar yield significantly and reduced the concentration of by-products, it did not affect the concentration of 1,3-PD (Zhao et al. 2009). The genetic code uses 60 nucleotide triplets (codons) to encode 20 amino acids and three codons to terminate translation. Each amino acid is therefore encoded by between one (Met and Trp) and six (Arg, Leu and Ser) synonymous codons. These codons are ‘read’ in the ribosome by complementary tRNAs that have been charged with the appropriate amino acid, high frequency usage codons usually own a larger number of corresponding tRNAs than low frequency usage ones. The frequencies of different codons vary significantly between different organisms, even between proteins expressed at high or low levels within the same organism, and sometimes even within the same operon (Gustafsson et al. 2004). Codon distribution responds to genome GC content and the changes in codon usage are at least partly explained by a mutation-selection-equilibrium among different synonymous codons in each organism (Knight et al. 2001). Thus it is definitely necessary to optimize codons when expressing functional proteins in heterologous hosts. Many typical increasement in expression for mammalian proteins from E. coli are between 5- and 15-fold (Sinclair and Choy, 2002; Slimko and Lester, 2003). However, the codon optimization of dhaT gene from K. pneumoniae for protein expression in E. coli has not been reported yet. The PDOR expression level needs to be enhanced to perform in vitro coupled with enzymatic reaction. Moreover, we can further reform this optimized strain by recombining with other key enzymes for the 1,3-PD in vivo fermentation. In this paper, the PDOR gene dhaT from K. pneumoniae was de novo synthesized by splicing overlap extension polymerase chain reaction (SOE-PCR). The comparison of different inducers for expression in E. coli and characterization of the recombinant enzyme were discussed. Materials and MethodsK. pneumoniae DSM2026 was obtained from the German Collection of Microorganisms and Cell Culture (presented by Dr. An-Ping Zeng, GBF, Biochemical Engineering Division, Germany). E. coli BL21 (DE3) and DH5α are conserved in our laboratory. Plasmid pUC57 was purchased from Sangon (Shanghai, China). Plasmid pET-15b was purchased from Novagen (Darmstadt, Germany). NAD was bought from Roche (Shanghai, China). Restriction enzymes were bought from TaKaRa (Dalian, China). All other chemicals used in this study were of analytical grade commercially available. Codon optimization, gene design and de novo synthesis According to the codon usage in E. coli (Hénaut and Danchin, 1996) at gene highly and continuously expression period, we defined the least preferred codon by usage frequency below 10% and all those codons were modified into the most preferred ones without changing amino acid sequence. More preferred codons should be used at N-terminal to promote translation quickly, while less preferred ones were introduced to enhance the termination of protein translation at C-terminal. As for other codons at different frequencies, they were changed on the basis of codon usage table of E. coli. Meanwhile, GC content was adjusted to be consonant with that in E. coli and decreased as low as possible to prevent stable secondary structure of mRNA, as simple and loose mRNA secondary structure was favourable for translation (Wu et al. 2004). When the same residue repeated in sequence, the identical codon was avoided otherwise corresponding tRNAs would be scant during that time. On the premise of identical amino acid sequence (Accession No. CP000647.1), 29 pairs of primers were designed with 25 nucleotides overlap for each pair. SOE-PCR was performed with primers using Pfu DNA polymerase (BBI, Can). These primers were self-complemented through 30 cycles of denaturizing (94ºC for 30 sec), annealing (55ºC for 30 sec), and extension (72ºC for 2 min). Then, the production of former step was used as template to amplify full-length dhaT sequences in the second cycle of PCR under the same conditions. DNA fragments were purified with EZ Spin Column DNA Gel Extraction Kit (BBI, Can) and ligated into pUC57 vector by restriction enzyme SmaI. The resulting plasmid (pUC57-opt) was confirmed by EcoRI and SalI double digestion, then transformed into E. coli DH5α. The restriction enzyme sites NdeI and BamHI were designed on both ends. Double digested pUC57-opt with NdeI and BamHI to construct the expression vector pET-15b-opt. After being transformed into E. coli BL21 (DE3), the positive transformants were selected on Safranin O selection medium (10 g/L NaCl, 6 g/L tryptone, 3 g/L yeast extract, 18.4 g/L glycerol, IPTG 1mM, safranin o 0.1 mM and 20 g/L agar). Then vectors were extracted by EZ Spin Column Plasmid Mini-Preps Kit (BBI, Can) and confirmed by double digestion. PDOR expression and activity assay The transformed single colony was inoculated in 5 ml LB medium (10 g/L NaCl, 10 g/L tryptone, 5 g/L yeast extract) and cultured for 16 hrs at 37ºC. Then cell culture was inoculated to 50 ml fermentation medium (12 g/L tryptone, 24 g/L yeast extract, 3 g/L NH4Cl, 4.5 mL/L glycerol, 1.7 mM KH2PO4, 7.2 mM K2HPO4) for 2 hrs at 30ºC. We added inducers (IPTG or lactose) to the medium at certain concentration to promote expression at 25ºC. Induced E. coli were collected by centrifugation and broken up by sonication at 300 W for 2 sec operation and 2 secinterval with 60 cycles in ice bath, then centrifuged at 15,000 g for 15 min to collect the supernatant. Ni-IDA resin was used to purify PDOR by affinity chromatography. Proteins in the supernatant and purified ones were detected by 10% SDS-PAGE and stained with Coomassie Brilliant Blue R-250. The activity of PDOR was determined as described by Ahrens et al. 1998 with modification according to Chen (Hongwen et al. 2005). Both cell-free extract and purified protein were analyzed by dynamic absorbance at 340 nm in potassium carbonate buffer of diverse pH and temperature values. Enzymatic properties were studied by purified PDOR. For optimal temperature and thermostability, enzymatic activity was analyzed in 100 mM potassium carbonate buffer (pH 9.5), incubated at 30, 40, 45, 50, 55 or 60ºC. Thermostability was first investigated by incubating the enzyme at 30, 45, 55 or 65ºC for 2 hrs. The residual enzyme activity was determined at pH 9.5 and 45ºC. For optimal pH and pH stability, enzyme was analyzed variable pHs, from pH 8.5 to pH 12. In metal ions and substrate specificity effects, 1 mM various metal ions or different substrate on PDOR activity were detected at 45ºC in pH 9.5 buffer. The standard deviation was calculated by three repeated experiments. Results and DiscussionSequence modification and improvement of secondary structure of mRNA E. coli possesses advantages in many fields such as high growth rate and clearly known metabolic network, thus it proves to be the most suitable host to express heterologous proteins from procaryotes. According to codon usage table (Hénaut and Danchin, 1996), we revised nucleotide sequence in the whole gene range. Compared with wild type, the optimized sequence shared 83.76% identity, however, 189 nucleotides which accounted for 16.24% were replaced (Figure 1). GC content decreased from 58.7% to 54.9%, and it was consistent with the genomic GC content of E. coli. Moreover, energy of mRNA secondary structure was increased from -400.1 to -86.8 kcal/mol (Figure 2). The decreased GC content would alleviate the formation of stable secondary structure, which was beneficial for translation (Wu et al. 2004). Judged by the CAI value (Henry and Sharp, 2007), it was increased from 0.75 to 0.81, which indicated the optimized gene was closer to the highly expressed genes in E. coli. PDOR expression We performed double digestion on pUC57-opt by EcoRI and SalI. As for pET-15b-opt recombinant, XbaI and BamHI were used to testify the positive clone. Our target gene was at the length of about 1.2 kb. The transformants were cultured in LB medium overnight and transferred to fermentation medium for PDOR expression. The enzyme activities (Figure 3a) and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) of cell-free extract were both analyzed (Figure 3b). As shown in Figure 3a, the enzyme activity of optimized strain was much higher than wild type in the whole process of induced expression. The activity increased and reached to the maximum point: 385.4 ± 3.6 U/mL, which was almost 5-fold higher than wild type (82.3 ± 1.5 U/mL) at 10 hrs, and the PDOR accumulation rate was significantly faster. Then it decreased quickly due to the toxicity of IPTG for E. coli, even over-produced protein could be toxic and result in host cells death (Miroux and Walker, 1996). Long-standing IPTG would cause cells to autolysis and decompose proteins (Kweon et al. 2001), so the effect on optimized strain was much heavier. The specific activity for optimized and wild type strains in Table 1 were 25.4 ± 0.2 and 9.3 ± 0.2 U/mg correspond to maximum enzyme activity 385.4 ± 3.6 and 82.3 ± 1.5 U/mL, respectively at 10 hrs. The maximum specific activity for optimized and wild type strains in Table 1 were 62.1 ± 2.7 and 9.6 ± 0.1 U/mg, respectively at 6 hrs. The former was nearly 7-fold higher than the latter due to the higher content ratio of PDOR in the cell-free extract. The specific activity of purified PDOR by affinity chromatography was 50.5 U/mg. From the analysis of SPS-PAGE (Figure 3b), we found our target protein was at the position around 48 kDa. The pure enzyme was 42 kDa and Histidine-tag was 6 kDa. Through densitometry analysis to the target bands by Tanon GIS (Shanghai, China), the protein amount from optimized strain (lane 3) was approximately 4-fold of the wild type (lane 2). Thus, significant higher expression level resulted in the enhancement of enzyme activity. Lactose, analog to IPTG and IPTG are both considered as inducers for T7 promoter, however, their inductive ability are quite different. In Figure 4, IPTG was more powerful in expression level and efficiency. PDOR expression induced by IPTG was much faster than by lactose, it reached the highest activity after 10 hrs induction. The expression level for lactose induction was close to IPTG at 6 hrs to 8 hrs. But it was consumed as a kind of carbon source and concentration kept declining. The expression was heavily affected when lactose was consumed by a large amount but kept carrying on at a low pace till it was finally used up at 12 hrs. Since the cells were in good condition unlike cells induced by IPTG, the enzyme activity maintained at the maximum point and sharp decrease didn't take place. Recently, a study revealed the possible reasons for IPTG effect on enzymes expressed in E. coli. The loss of specific activity and stability were correlated to incomplete attachment of cofactors onto enzyme (Zheng et al. 2011), so synthesis speed should be controlled by reduction of IPTG amount. Enzymatic properties study The optimal pH for PDOR was pH10.0 (Figure 5a). Its activity maintained at high level between pH 9.0 and 11.0, which indicated it was an alkaliphilic enzyme. Below pH 9.0, the activity decreased dramatically, but when pH exceeded 10.0, it decreased slowly to 50% at pH 12.0, so the enzyme preferred alkaline environment. The stability of PDOR in diverse pH environments were determined (Figure 5b). PDOR was pre-treated in potassium carbonate buffer of diverse pH values for 2 hrs at 45ºC. The most stable environment for PDOR was at the optimal pH 10.0, where the enzyme remained approximate 60% activity. The optimal temperature of PDOR was 50ºC (Figure 5c) while activity remained more than 80% in the range of 30 to 50ºC. However, it quickly declined when the temperature was higher than 50ºC. Only 45% activity remained when the temperature came to 60ºC (all the experiments were tested in pH 9.5 buffer). In other words, this enzyme favoured the mild thermal condition. The thermal stability of PDOR was carried out in pH 9.5 buffer at diverse temperatures for 2 hrs. In Figure 5d, 30ºC and 45ºC had activation effect on enzyme which was conserved at freezing before reaction, the activity went through a sharp down from 30 to 60 min, drop tended to slow down since then. The relative activity was 50% and 30% at 30 and 45ºC respectively. At higher temperatures, the activity decreased quickly at the first 30 min and dropped continuously as time went by. The enzyme lost activity after 120 min at 55ºC, while 90 min at 65ºC. Effects of various metal ions (1 mM) on PDOR activity were detected at 45ºC in pH 9.5 buffer (Figure 6a). Compared by the standard activity of PDOR with none metal ion added, the enzyme was inhibited by some metal ions, such as Na+ and Ca2+, its activity had 35% and 30% decrease. While, PDOR was activated by NH4+, Fe2+ and especially Mn2+, which was increased by 30%, 40% and 130%, respectively. Generally, divalent cations showed a positive activation function and Mn2+ was the best one in our results. In the reaction with metal Mn2+ which showed significant improvement on PDOR activity was validated by Daniel (Daniel et al. 1995). Moreover, PDOR was a kind of Fe-dependent dehydrogenase (Marçal et al. 2009), Fe ion played an important role in enzymatic catalytic domain. Substrate selectivity was studied from a series of monohydric alcohols and diols at 45ºC in pH 9.5 buffer (all these substrates were catalyzed in oxidation reactions). According to Figure 6b, the optimized PDOR showed a universal applicability to alcohols, except for 2-propanol. The catalytic efficiency to 1,3-PD and 1,2-propanediol (1,2-PD) were both over 90%, while the activity of PDOR to glycerol and 1-propanol were at ~70%. PDOR was also active to substrates like 1,4-butanediol (1,4-BD) and 1,3-butanediol (1,3-BD) except for the toxic 2-propanol. Concluding RemarksAs heterologous genes might contain codons that are rarely used in the expression host, come from organisms that use non-canonical code or contain expression-limiting regulatory elements within their coding sequence; proteins are often difficult to express outside their original context (Gustafsson et al. 2004). In this paper, the codon-optimized PDOR gene was efficiently expressed in E. coli with the activity of 385.4 ± 3.6 U/mL, which was the highest activity ever reported. It was almost 5-fold higher than wild type (82.3 ± 1.5 U/mL) in the cell-free extract, the corresponding specific activity was 25.4 ± 0.2 U/mg. Our result was much better than those expressions of dhaTgene from K. pneumoniae in the heterologous host. Cao et al. 2006 expressed PDOR in Escherichia coli using the methods of uniform design with activity of 9.25 U/mL; Ma (Ma et al. 2010) expressed dhaT gene in Escherichia coli and Saccharomyces cerevisiae with the specific activities are 0.9 and 12.3 U/mg, respectively. Meanwhile, the specific activity was also much higher than the reported 1.51 U/mg (Zhao et al. 2009) and 1.56 U/mg (Hao et al. 2008) that were directly expressed in K. pneumoniae. Thus, we achieved a fruitful result on protein expression in E. coli by codon optimization. The optimal condition for enzyme activity was at 50ºC and pH 10.0. It was also stable in alkaline buffer and meso-thermal condition. The metal Mn2+ with significant improvement on PDOR activity was in consistency of the study by Daniel et al. 1995. AcknowledgmentsThe authors would like to thank the anonymous reviewer for their valuable comments and insights that helped to greatly improve this manuscript. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11044f2.jpg] [ej11044f1.jpg] [ej11044f6.jpg] [ej11044f4.jpg] [ej11044f3.jpg] [ej11044f5.jpg] [ej11044t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}