 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 4, July 15, 2011 RESEARCH ARTICLE Detecting adventitious transgenic events in a maize center of diversity Luis Fernando Rimachi Gamarra1 · Jorge Alcántara Delgado1 · Yeny Aquino Villasante1 · Rodomiro Ortiz*2 1Instituto
Nacional de Innovación Agraria, Lima, Perú Code Number: ej11046 AbstractBackground: The
genetic diversity of maize in Peru includes several landraces (within race
clusters) and modern open pollinated and hybrid cultivars that are grown
by farmers across various regions, thereby making this country a secondary
center of diversity for this crop. A main topic of controversy in recent
years refers to the unintended presence of transgenic events in locally grown
cultivars at main centers of crop diversity. Peru does not yet have biosafety
regulations to control or permit the growing of genetically modified crops.
Hence, the aim of this research was to undertake a survey in the valley of
Barranca, where there were recent claims of authorized transgenic maize grown
in farmers fields as well as in samples taken from feed storage and grain
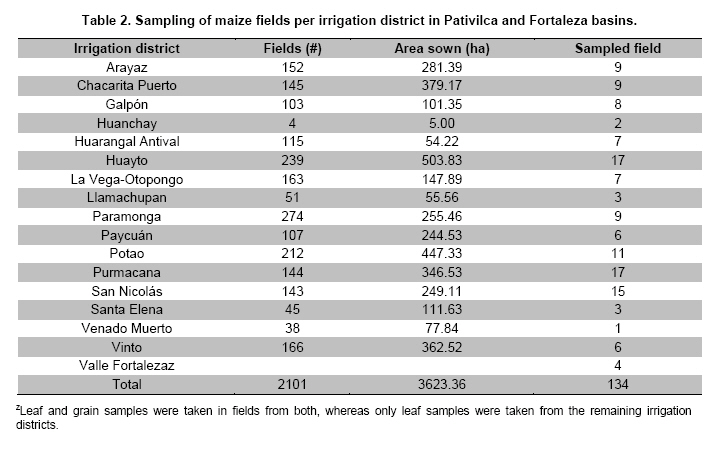
or seed trade centers. Keywords: biosafety, corn, sampling, transgenes, Zea mays. IntroductionA lot of maize genetic variation occurs in Peru (Grobman et al. 1961), which may be regarded as an important center of diversity for this crop. Sevilla (2005) indicates that there are about 55 Peruvian races of maize that played an important role in the development of modern maize cultivars, particularly in the highlands Table 1. Maize races have been extensively studied and classified using specific ear and kernel traits (Grobman et al. 1961). This maize germplasm clustering was further confirmed with modern numerical taxonomy methods (Ortiz et al. 2008a; Ortiz et al. 2008b). Highland farmers grow distinct races in their maize fields that led to cultivar mixtures due among other causes, to gene flow through pollen, close cropping of diverse landraces or formation of seed banks. Maize races are, however, easily distinguished by farmers, particularly when “foreign genes” are brought from modern hybrids. Gene flow is not something peculiar to transgenic plants. It happens at any time one organism breeds with a related species, thus passing along their combined DNA to the offspring. A main issue, which led to strong debates elsewhere, is the adventitious presence of transgenic events in main centers of crop diversity (Quist and Chapela, 2001; Christou, 2002; Kaplinsky et al. 2002; Metz and Fütterer, 2002; Quist and Chapela, 2002; Celis et al. 2004; Ortiz-García et al. 2005; Raven, 2005; Mercer and Wainwright, 2008; Piñeyro-Nelson et al. 2009; Schoel and Fagan, 2009). For example, the potential genetic and ecological impacts of gene flow from transgenic cultivars to landraces, weedy relatives and wild species are mainly related to the genetic integrity of landraces and crop wild relatives, and to developing plants with enhanced invasiveness or weediness in ecosystems (Cleveland et al. 2005; Engels et al. 2006; Scurrah et al. 2008; Warwick et al. 2009; Sahoo et al. 2010). Farmers' behavior and crop husbandry may significantly influence transgene spread in native germplasm. However, the perceptions of farmers and consumers that the transgenes are “polluting” and that landraces or local cultivars containing transgenes are “contaminants” could cause that these landraces or local cultivars may be rejected, which would mean a direct loss of agro-biodiversity (Bellon and Berthaud, 2006). The global spread of transgenic crops has also significant implications for organizations involved in germplasm conservation and genetic enhancement. In this regard, Mezzalama et al. (2010) describes a protocol used for monitoring unintentional transgene flow in maize gene bank and breeding plots. Their protocol is based on polymerase chain reaction (PCR) markers for detecting specific recombinant DNA sequences in bulked samples collected from sentinel plots. Peru does not yet have biosafety regulations to control or permit the growing of genetically modified crops, and their introduction is a source of lively debate in the Peruvian media (Laursen, 2011). Very recently, Gutiérrez-Rosati et al. (2008) indicated that 1/3 of 42 samples of yellow maize grains from the valley of Barranca (north of Lima, Peru) were positive for transgenic events SYN-BTØ11-1 (BT11) and MON-ØØ6Ø3-6(NK603), which provide host plant resistance to insect and tolerance to glyphosate herbicide, respectively. Their reports refer to both grains from harvests in this valley as well as from stores of animal feed. The 1999 Peru’s Law 27104 (Prevention of risks from the use of biotechnology) and the 2002 Supreme Decree No. 108-2002 (regulating this law) empowers the Instituto Nacional de Innovación Agraria (INIA) as the sectoral body in agriculture to enforce provisions under national and international policy, to regulate, manage and control risks arising from the contained use and environmental release of living (LMO) or genetically (GMO) modified organisms. INIA formally asked for more information to Gutierrez-Rosati on the location of the fields where the samples were obtained, and the submission of the respective counter samples to validate their claim. Unfortunately, further details or the respective counter samples were not provided. The main goal of our research was therefore to assess qualitatively the presence of promoter P35S and sequence of Tnos terminator, to detect 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) and Cry1Ab delta-endotoxin from NK603 and BT11 respectively, using immune-assays, as well as six commercial transgenic events, namely BT11, NK603,ACS-ZMØØ3-2(T25), SYN-EV176-9 (176), DAS-Ø15Ø7-1 (TC1507) and MON-ØØ81Ø-6 (MON810), with the aid of PCR amplification using event specific primers in maize samples taken from farmers fields, local markets, seed trade centers or barns of poultry farms in the valley of Barranca and neighboring locations following proper sampling and screening methods. Materials and Methods The sampling area of maize fields was mainly along the Pativilca River Basin Figure 1, which is the main river of the valley of Barranca. Four sites from the neighboring Fortaleza River Basin were also added to this field sampling. There were additional grain samples from one local maize collecting center, eight poultry farm barns, four private seed dealers and 15 local markets. Each maize field was regarded as an experimental unit, thereby estimating the sample size (n) for a categorical variable (presence or absence) with a finite population size as follows (Cochran, 1977):
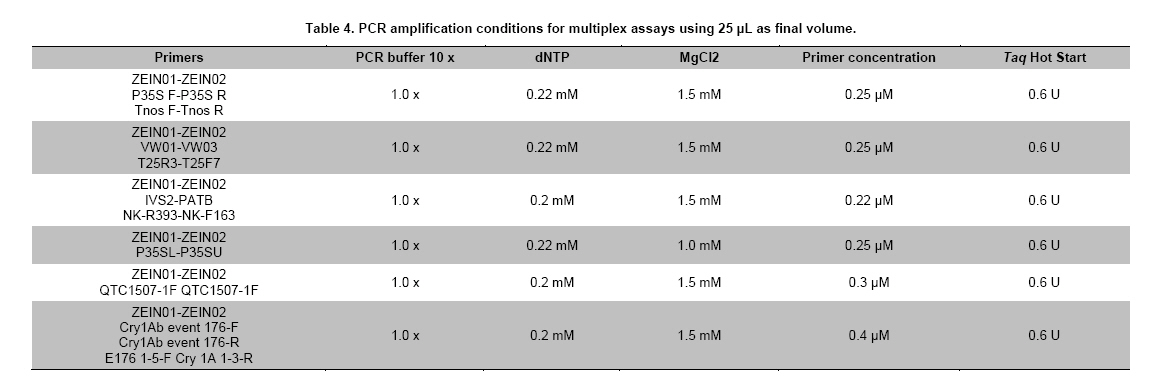
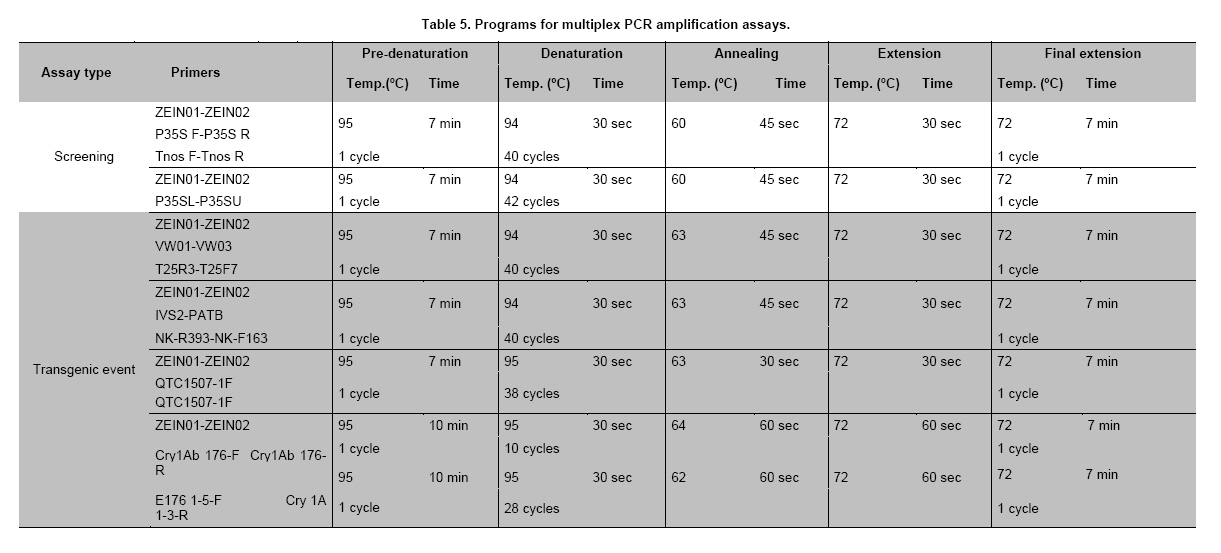
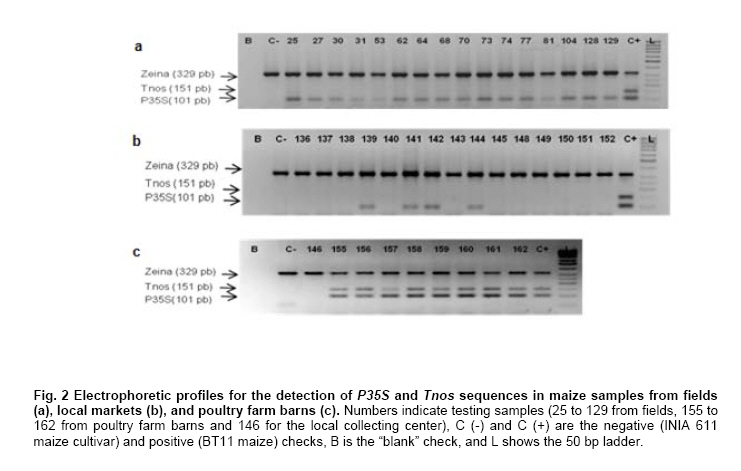
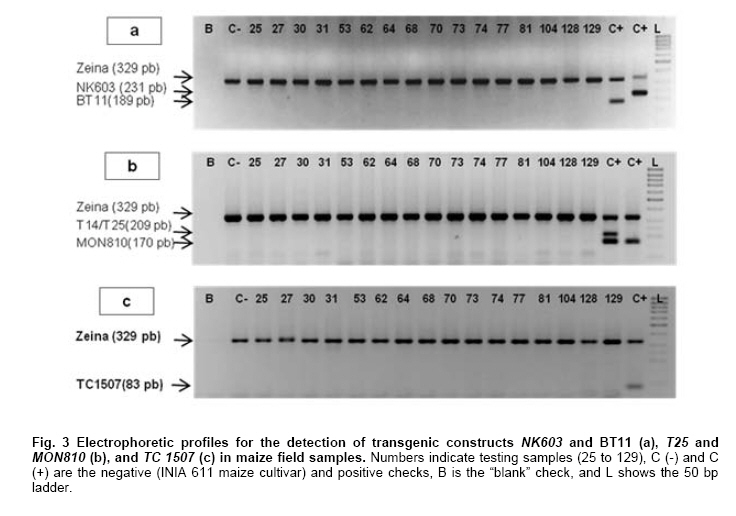
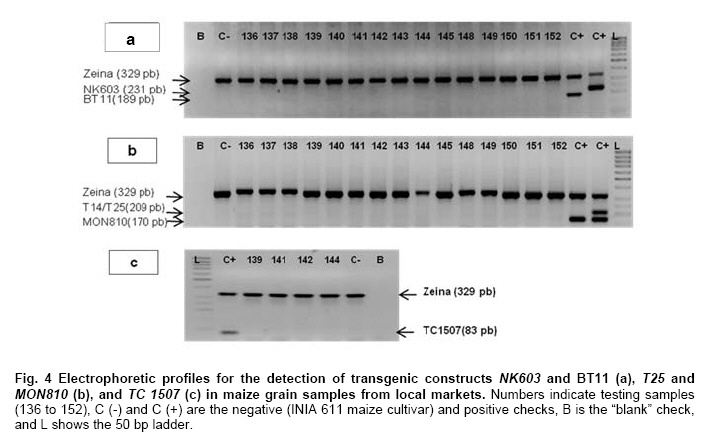
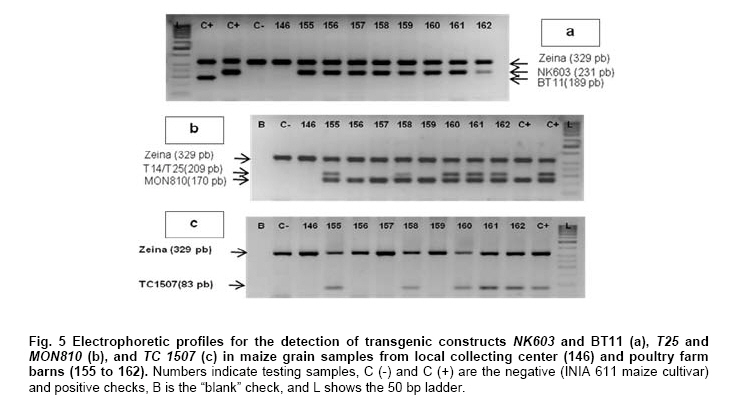
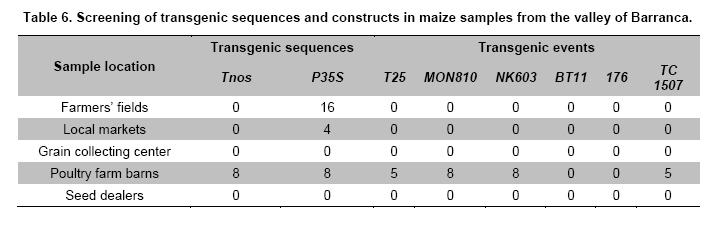
Where N is the population size; i.e., the total number of maize fields (2100), p the prevalence (0.1), q equals 1 – p (0.9), d is the precision, α is the significance level (0.05), 1- α is the confidence level, and Z1- α a pre-established value. The sample size used was 130 maize fields taking into account the above sampling equation and the putative transgene frequency (33.3 to 62%) in the valley of Barranca (Gutiérrez-Rosati et al. 2008). The sub-sampling within each location used the probability of detection (Pd) as follows (Remund et al. 2001; Lockwood et al. 2007): Pd = 1 – (1 – pGM)m.s Where pGM is the uniform frequency of a genetically modified organism (GMO), m is the number of fields or seed lots sampled, and s is the number of individuals or 2n alleles sampled per field or seed lot. This protocol could allow, with a 95% probability, detecting transgenic events with a frequency equal or greater than 0.05% ensuing from unauthorized GM-seed imports when sampling at least 50 plants in each location. The probability of detection of GMO with a frequency equal or greater to 0.05% (using a PCR assay) will be 96.13% following above equations used for this two-step sampling approach of 130 maize fields and 50 plants per field. The number of maize fields sampled was determined according to their relative number in each irrigation district Table 2. A zigzag walk was used for leaf sampling in a minimum of 1 ha and taking one leaf per plant from at least 50 plants. Some farmers maize fields included in the field survey were larger than 1 ha and due to logistics only 1 ha was taken randomly for sampling. The leaf samples had between 5 to 10 cm in length and preferably from the middle part of healthy tender leaves. Two thousand grains were collected from each of the maize fields that were at harvest time; i.e., 100 randomly selected ears were selected from each field, and 20 grains were obtained from two rows per each ear. This grain sampling provides a 99% certainty to detect the adventitious presence of transgenic events with P ≥ 0.005. Grain samples (500 g ≈ 2100 grains) from local markets were purchased from the main wholesalers and retailers. They were grouped into three subsamples of approximately 700 seeds each. Based on the binomial probability, if the 3 sub-samples showed negative results in the PCR analysis, there would be a 95% certainty that the transgenic event frequency was below 1%. Similar approach was used for grain samples (of same weight) from the local maize collecting center and private seed dealers. Seed samples of hybrid maize cultivars Agroceres 003 and Agroceres 1596 were kindly provided by a local dealer. Four samples (≈ 2 kg) were taken from two grain lots in each of the poultry farm barns. The analysis of all field samples were carried out in two stages, the first called scanning or screening used qualitative PCR detection for P35S and Tnos sequences, which are present in most transgenic maize events. The second stage involved the identification of specific transgenic events that were indicated as grown in Peru by previous reports (Gutiérrez-Rosati et al. 2008). Three of them have P35S and Tnos sequences (BT11, NK603 and MON863) and one (T25) only has P35S. DNA was extracted from each leaf sample following a modified CTAB method (Doyle and Doyle, 1990). The DNA from grains was taken according to the manual of the EU for detecting GMO in food samples (Querci et al. 2006). DNA extraction was from 1 cm2 each in groups of 10 leaves due to the number of samples; i.e. a total of five DNA sub-samples every field with its own duplicate. DNA was quantified automatically on the Nanodrop 2000, by the standard spectrophotometric relations to 260 nm, 260/280 nm and 260/230 nm. The quality of DNA was visualized by electrophoresis on 0.8% agarose gels (Sambrook and Russell, 2001). All samples were standardized at a concentration of 10 ng ml-1 before mixing the five sub-samples for their later use in PCR amplification. Protocols and programs for qualitative PCR amplification were standardized following known protocols Table 3. New multiplex PCR assays were standardized for the analysis of two or three primers per reaction Table 4 and Table 5 with the aim of reducing costs and time. The primers were synthesized by Invitrogen (São Paulo, Brazil) and IDT (Coral Ville, Iowa, USA) whereas other reagents used in PCR amplification (10 x PCR Buffer, dNTP, MgCl2, Taq polymerase) were from QIAGEN (Hamburg, Germany). The controls for the PCR amplification assays and for the analysis of amplification products by electrophoresis were BT11, NK603, MON810, TC1507, 176 and T25 [positive checks provided by the Instituto Nacional de Tecnología Agropecuaria of Argentina (INTA)], a 329 bp zein gene (DNA amplification check), DNA from hard yellow maize cultivar INIA 611 (negative check), and a PCR master mix without DNA (“blank” check). Amplified products were separated by electrophoresis on 2% agarose gels (120 volts x 80 min) and visualized by ethidium bromide staining (0.3 mg ml-1) and photo-registered with ChemiDocTM XR. The amplified product size of the samples analyzed for each of the primers and positive controls (at a 5% weight/weight in the working GM/non-GM samples provided by INTA) were compared with the ladder of DNA fragments of 100 bp (Invitrogen: 1500 to 100 bp) and 50 pairs bases (Fermentas/Gen Lab del Peru S.A.C., Lima, Peru): 1031 to 50 bp). Eye scoring for absence or presence of transgenic constructs was used for recording into a database. Immunoassay for detecting GMO using lateral flow strips was conducted in farmers' fields. The kits for the detection of Cry1Ab and CP4 EPSPS proteins of transgenic events Bt11 and NK603, respectively, were purchased from Estrategic Diagnostic (Newark, Delaware, USA). Other kits to verify the detection of the same proteins from transgenic events Bt11 and NK603 were also kindly provided by AGDIA (Elkhart, Indiana, USA). The methodology for using both sets of kits was described in the respective company manuals. Results There were 127 maize leaf DNA samples from the sampled fields (94.77%), whose concentrations ranged from 20 to 150 ng ml-1. The DNA obtained from grain samples had a concentration of 40 to 130 ng ml-1. The seven samples with non-amplifiable DNA for PCR analysis were collected from fields at harvest time. The degree of leaf deterioration did not allow to obtain quality DNA and to get appropriate concentrations. Hence, the probability of detecting a GMO with a frequency greater or equal to 0.05% was adjusted to 95.82%. Sixteen out of 127 field samples that amplified the endogenous maize gene region were positive thrice for P35S (101 bp), but none of these samples was positive for Tnos Figure 2a. Four of the 15 samples from local markets were positive thrice for P35S but they were negative for Tnos Figure 2b. The eight grain samples from the poultry farm barns amplified for both P35S and Tnos sequences Figure 2c, whereas the grain sample from the local maize collection center or the local seed dealers did not amplify for either. The 16 field samples and four samples from local markets showed faint bands for P35S, compared to well-defined bands from samples of the poultry farm barns. None of the 127 field samples, including the 16 samples that amplified the P35S sequence, showed positive results for the presence of transgenic constructs BT11, NK603, T25, 176, TC1507 and MON810 in the three repetitions used Figure 3. There were no positive results for the presence of the same transgenic constructs in the three repetitions for tests on 15 grain samples from local markets, including the four samples that amplified the sequence P35S Figure 4. Five of the eight grain samples from the poultry farm barns amplified the transgenic construct T25, whereas eight samples amplified from the transgenic constructs NK603 and MON810 Figure 5. The transgenic constructs 176 and BT11 were not found in any of the grain samples. The grain sample from the local collecting facility did not amplify any of these six transgenic constructs The immunoassays using lateral flow trips for Cry1Ab-delta endotoxin and EPSPS with field samples were negative. Samples from positive (NK603 and Bt11) and negative (INIA 611) cultivar checks were used to validate the functionality of these lateral flow strips. Discussion The Barranca Valley is an agricultural area that primarily grows yellow maize, particularly commercial hybrid cultivars (93.3%) from private seed suppliers (Agricola, Agroceres, Dekalb, Pioneer HiBred, Hortus and Inti). Those commercial maize hybrid cultivars that do no longer produce good grain yields are used for fodder (locally known as "chala") and account for 4.5% of the field samples. Only three fields, of the 134 randomly selected for sampling, had landraces or local cultivars (2.2%), including two for green maize (or "choclo" as per its vernacular name), and one purple maize (for producing the local drink "chicha morada" or desserts such as "mazamorra morada"), whose seeds can be purchased in local markets or are kept by farmers for re-seeding at every planting. The initial screening for adventitious transgenic events was only to assess the presence of BT11 and NK603, which are widely distributed worldwide and were reported to be in maize samples from Barranca (Gutiérrez-Rosati et al. 2008). We decided to screen further other transgenic constructs (MON810, T25, TC1507 and 176), which possess P35S but lack Tnos, after being unable to detect BT11 and NK603 in the samples analyzed. The screening results from field samples were also negative Table 6. Transgenic events GA21 and MON863 were not included for subsequent analysis because both have the Tnos sequence, which was negative in the previous screenings. The finding of P35S on 16 field samples could be false positives due to the presence of the Cauliflower Mosaic Caulimovirus (CaMV) in these samples, as was also indicated by research elsewhere (Wolf et al. 2000; Holden et al. 2010). Another possible explanation would be a slight contamination in the laboratory. However, the negative checks for PCR amplification did not yield positive results in any test conducted, thereby ruling out this possibility. The positive results for transgenic events in grain samples from poultry farm barns could be attributed to the high demand for yellow maize by the poultry industry. Peru imports about 1.5 million t (in excess of 50% of the national demand) of maize grains mainly for animal feed every year from Argentina (75% of total import of maize grains) and USA (21%), where GM-maize seeds are widely grown by their farmers and traded in export markets. It is very important to use sound sampling protocols, analytical methods (Anklam et al. 2002) and probability models (Hernández-Suárez et al. 2008) for detecting adventitious transgenic events. We can conclude, based on our screening results with a 95% confidence level and a 95.82% probability of detecting adventitious transgenic events with a frequency equal or greater than 0.05%, that farmers do not grow transgenic maize cultivars in the valley of Barranca. Previous research about the presence of transgenes in maize samples from this valley (Gutiérrez-Rosati et al. 2008) did not indicate if they were found in native maize cultivars. Hence, there is a lack of evidence for a possible hybridization between the landraces and GM cultivars of maize, and it seems very unlikely that such possible introgression of transgenes occurs in Peruvian maize landraces. Pollen flow from maize hybrids to local cultivars often occurs in farmers fields of the Peruvian coast. However, when selecting their seeds for planting, farmers retain the varietal purity of their landraces and local cultivars because the grains have special uses in drinks such as "chicha", or are freshly eaten as "choclo" (Sevilla, 2005). The seeds ensuing from the fertilization with pollen from hybrid yellow maize cultivars are easily distinguishable by the xenia effect and therefore dismissed as seed for planting by the local farmers. Furthermore, Palaudelmàs et al. (2009) found that transgenic maize volunteers had low plant vigour, rarely had cobs and produced pollen that cross-fertilized neighbour plants only at low levels. Transgene flow raises a new set of ecological and economic issues for scientists and policymakers to consider for transgene containment (Dyer et al. 2009). Local farmer knowledge will be useful to avoid transgene flow and maintain distinct cultivars for the markets (Ortiz and Smale, 2007). Appropriate measurements should be also taken in Peru when transgenic and conventional crops of the same species will coexist in the future in the same locations if some farmers will wish to grow crops for GMO-free markets. Such regulations will also benefit from recognition of the practices farmers use to maintain the genetic integrity of their cultivars in their fields. Acknowledgments The authors thanks Dr. Felipe de Mendiburu for his advice on sampling and statistical analysis, Prof. Ricardo Sevilla for reviewing the spatial distribution of local maize landraces and cultivars, colleagues from the Agencia Agraria de Barranca, Gerencia Regional de Recursos Naturales y Gestión del Medio Ambiente from Gobierno Regional de Lima and INIA for the assistance in collecting maize samples, and to the Junta de Usuarios del Valle Pativilca and her farmers for both assisting in collecting samples and allowing to take them from their fields. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile -->ISSN: 0717-3458 Contact: edbiotec@ucv.cl Pontificia Universidad Católica de Valparaíso The following images related to this document are available:Photo images[ej11046t5.jpg] [ej11046t4.jpg] [ej11046f4.jpg] [ej11046t6.jpg] [ej11046t3.jpg] [ej11046f3.jpg] [ej11046f1.jpg] [ej11046t1.jpg] [ej11046t2.jpg] [ej11046f2.jpg] [ej11046f5.jpg] |
| |||||||||

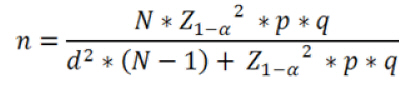
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}