 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 4, July 15, 2011 SHORT COMMUNICATION Stability inside hen eggs of a Salmonella enterica serovar Enteritidis bacteriophage James Robeson*1 · Martina Valencia1 · Julio Retamales1 · Consuelo Borie2 1Instituto
de Biología, Facultad de Ciencias, Pontificia Universidad Católica de Valparaíso,
Campus Curauma, Placilla, Valparaíso, Chile Financial support: We thank CONICYT, Chile, for financial support through FONDECYT Grant nº 1080291. J. Retamales is the recipient of a CONICYT Doctoral Scholarship. Code Number: ej11048 AbstractBackground: Bacteriophages
are viruses that infect bacteria and therefore are widespread in nature.
Those that lyse the pathogen Salmonella enterica serovar Enteritidis (SE) should be expected in niches in which this bacterium
thrives, among others the avian egg. Furthermore, bacteriophages could remain
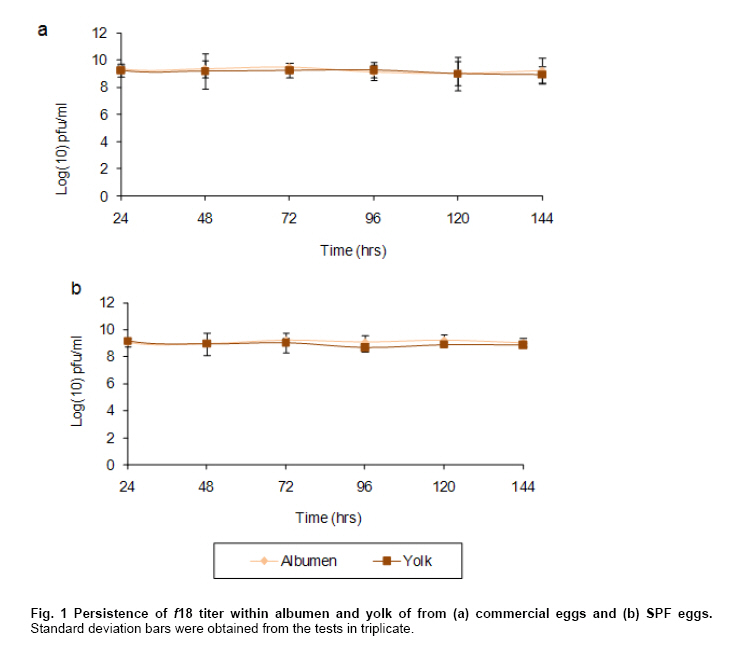
viable in this milieu. Keywords: bacteriophage, egg, Enteritidis, Salmonella. IntroductionBacteriophages or phages are viruses that infect bacteria. These biological entities are ubiquitous in nature; they are present in the oceans, soil, sewage, foodstuffs and practically every habitat where bacteria thrive since they are their natural predators and therefore play a crucial role in the control of natural bacterial populations. Lytic bacteriophages are particularly important in fulfilling such role because the outcome of their infective cycle is the lysis of the bacterial host cell together with the liberation of new phage progeny- or virions- able to infect other sensitive bacterial cells (Deresinski, 2009). Within this context, several lytic phages that infect Salmonella enterica serovar Enteritidis (SE) have been described. These have been isolated from poultry excreta and effluents from poultry farms and poultry-processing facilities (Atterbury et al. 2007), municipal wastewaters (Carey-Smith et al. 2006; Atterbury et al. 2007; Bielke et al. 2007) and faeces of free-range layers and other bird species (Fiorentin et al. 2004). Another niche where SE phage could be present is the hen egg. It is a fact that SE is equiped to infect the egg which acts as a vector for the transmission of this pathogen (Gantois et al. 2009). Therefore, at least a fraction of SE-infected eggs should be expected to carry SE bacteriophages. However this situation has not been reported, probably due to the relatively low frequency of infected eggs, normally one in a thousand (Alexandre et al. 2000), which are not routinely examined for phage. Alternatively, phage might not be detected because of inactivation by egg defences, since it is known that egg components such as ovotransferrine-derived peptides, ovoinhibitor, ovomucin and lysozyme inhibit viruses (Réhault et al. 2007). Furthermore, egg-white lysozyme has been shown to inhibit the replication of phage M13 by 97-99% in a plaque assay (Sitohy et al. 2006). Considering these latter data we sought to test the hypothesis of phage inactivation by experimentally infecting both SPF and commercial eggs with the SE phage f18 (Robeson et al. 2008) and testing its stability in the egg milieu. We found that phage injected in the albumen is later found in the yolk in practically the same concentration as in the albumen. Furthermore, there was no significant decrease in phage titer, both in yolk and albumen, after 144 hrs post injection. Phage and bacterial strain. A purified preparation of phage f18 obtained according to Sambrook et al. (1989) was used throughout our experiments. The bacterial strain used to determine f18 titers was Salmonella enterica serovar Enteritidis PT4, obtained from Dr. Roy Curtiss III, The Biodesign Institute, Arizona State University. A rifampicin (Rif) resistant mutant isolated according to Miller (1972) was employed. Bacteria were routinely grown at 37ºC in LB agar (1.5%) or broth (Miller, 1972). Determination of phage f18 stability in eggs. Commercial eggs were purshased at a local supermarket and specific pathogen-free (SPF) eggs were obtained from facilities at the Universidad Austral de Chile. To test stability of the bacteriophage titer in commercial and SPF eggs over time, we inoculated directly into the albumen of intact eggs 1 x 1011 plaque forming units (pfu) of f18 phage using sterile syringes and puncturing through the air chamber of each inoculated egg. Eggs were then incubated at 25ºC. Phage were allowed to diffuse for 24 hrs and subsequently, every 24 hrs and up to 144 hrs the eggs were processed to determine phage titers in yolk and albumen, separately. Yolks were sampled cauterizing the yolk membrane to prevent cross-contamination with phage in albumen, according to the method described by Gast et al. (2007). All determinations were performed in triplicate. Bacteriophages were titered performing suitable dilutions in phosphate buffer supplemented with 1 mM MgS04 and 0.01% gelatin (Robeson et al. 2008); 0.1 mL of each phage dilution was mixed with 0.1 mL of an exponentially growing culture of the indicator bacterial strain in 3 mL of molten (45ºC) soft LB agar (0.7%) which was overlayed on LB agar plates supplemented with Rif (100 ug/mL). After incubation for 18-24 hrs at 37ºC bacteriophage plaques that developed were counted. Statistical analysis Slopes corresponding to data of both commercial and SPF eggs groups (albumen and yolk) were compared using one way ANOVA (p < 0,05) by means of the program Statistica Application (Statsoft , Version 6.0). Results and DiscussionWe found that phage f18 is able to remain within commercial and SPF eggs keeping its ability to lyse SE without an appreciable loss in titer (Figure 1). Furthermore, no significant differences were found between the slopes of the titer curves for both yolk and albumen from commercial and SPF eggs. This is indicative that phage f18 is able to withstand the physical and chemical conditions of the egg's inner milieu. This contrasts with the results found for phage M13 (Sitohy et al. 2006). However, this might be due to the widely contrasting assay conditions or to inherent differences between M13 and f18 virions. This remains to be tested. Furthermore, we should point out that it is unlikely that titers of phage f18 could be boosted by the presence of SE bacteria in commercial eggs because we found no significant differences in phage titers between commercial and SPF eggs. Moreover, we tested randomly-picked commercial eggs (n = 20) for the presence of Salmonella bacteria and did not find any. As was pointed out before bacteriophages are biological entities that are widespread in nature (Deresinski, 2009) but there is no reference to the presence of phage in avian eggs. However, our results indicate the feasibility that eggs might be a potential niche for bacteriophage associated to bacteria naturally hosted by hens or phage that may accompany bacterial pathogens infecting such hosts and therefore a possible scenario for the bacteria-bacteriophage interaction. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11048f1.jpg] |
| |||||||||

{kind=link}