 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 14, No. 4, July 15, 2011 TECHNICAL NOTE Convenient genotyping of nine bovine K-casein variants Víctor I. Pacheco Contreras1 · Diana L. Lourenco Jaramillo1 · Gaspar M. Parra Bracamonte1 · Juan C. Martínez González2 · Ana M. Sifuentes Rincón*1 1Laboratorio
de Biotecnología Animal, Centro de Biotecnología Genómica, Instituto Politécnico
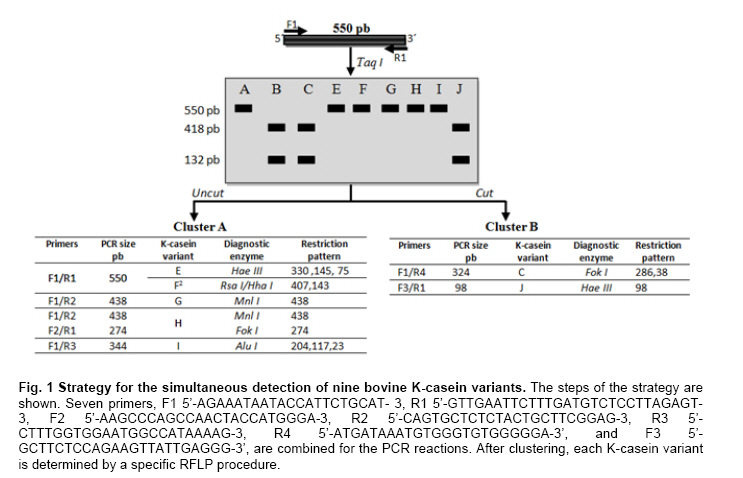
Nacional, Reynosa, Tamaulipas, México Financial support: Consejo Nacional de Ciencia y Tecnología (CONACYT), financiamiento del proyecto FORDECYT 116152, PIFI, SANTANDER. Code Number: ej11049 AbstractK-casein gene polymorphisms are of major importance in the dairy industry due to their association with different quality and productive traits (i.e., milk protein). Several methods for genotyping this gene have been proposed; however, none are focused on the simultaneous discrimination of nine K-casein variants. A strategy based on PCR-RFLP was designed to characterize nine K-casein variants (A, B, C, E, F2, G, H, I and J) and used to genotype three cattle populations: Gyrholando (3/8 Gyr x 5/8 Holstein), Charolais and Carora. The B variant was the most frequent in the Charolais and Carora breeds, with allelic frequencies of 0.60 and 0.59, respectively. In the Gyrholando breed, four variants were found, with the A variant being the most frequent and E and H the least. The genotyping strategy was effective in detection and differentiation of K-casein variants, and it is proposed for use in laboratories with minimal molecular biology equipment for genotyping and evaluation of the phenotypic effects of nine K-casein variants on milk production and quality. Keywords: milk proteins, milk quality, protein genes. IntroductionMilk is an essential source of nutrients for mammals during the first period of their life (Rijnkels, 2002; Otaviano et al. 2005). Its role in nutrition is extended throughout life for a vast majority of human populations. Strategies based on genetics and biotechnology have been developed to increase productivity and improve the quality of milk, and these include the Marker Assisted Selection (MAS) strategy (Rachagani and Gupta, 2008; Dogru and Ozdemir, 2009). Caseins are the most important proteins in mammal milk. They have been grouped into αs1-casein, αs2-casein, β-casein, K-casein and γ-casein derived from β-casein degradation. In cattle, the entire cluster is located on chromosome 6 (6q31) with a length of 200 to 250 kb (Sulimova et al. 2007; Azevedo et al. 2008; Rachagani and Gupta, 2008). Numerous genetic variants have been identified for caseins; their influence on milk composition has been demonstrated, and they have been used as an additional selection criterion in cattle breeding (Caroli et al. 2009). K-casein constitutes approximately 12% of the total casein, and due to its role in the regulation of micelle size and milk properties, it has been characterized across different species of the Bos genus, especially in different cattle breeds. Many studies on the k-casein gene have indicated that certain milk protein variants may be associated with milk production, milk composition and cheese production (Comin et al. 2008; Caroli et al. 2009). The B variant is associated with thermal resistance, shorter coagulation time, better curdles and micelle size while the A variant is associated with higher milk yield. The milk of cows with the BB genotype gives a 10% higher yield of cheese compared with the milk of cows with the AA genotype (Azevedo et al. 2008). This has led to the genotyping of cattle populations to confirm the reported effects. The K-casein gene contains five exons and four introns, with a length close to 13 kb. A 550-bp region in exon four contains all the polymorphic sites that produce the different variants described to date for this gene in cattle: A, A1, B, C, E, F1, F2, G, H, I and J (Chen et al. 2008; Prinzenberg et al. 2008; Caroli et al. 2009). With the exception of the A1 variant, the differences are based on non-synonymous nucleotide changes. Chen et al. 2008, resequenced this polymorphic region and found additional variation, especially in the produced haplotypes of the gene, and the authors suggested that resequencing should be done in local cattle breeds to estimate the real genetic variability and evolution of the k-casein gene (Chen et al. 2008; Prinzenberg et al. 2008). In cattle, the k-casein gene has been studied using both screening and scoring technologies, which include techniques based on PCR-RFLP, PCR-SSCP, sequencing and microarrays. Some assays have been developed and used for typing most of k-casein variants, including those whose productive effect has been determined or which have been reported to be most frequent in cattle breeds. However, none of these have focused on the simultaneous detection of up to six variants (Farrel et al. 2004; Chessa et al. 2007; Caroli et al. 2009). We describe a procedure for genotyping nine of the k-casein variants using PCR coupled to restriction fragment length polymorphism (PCR-RFLP) and its variants such as Artificially Created Restriction Site (PCR-ACRS). Materials and MethodsThe strategy for the detection of nine variants of the k-casein gene is based on their amino acid differences (Chen et al. 2008; Prinzenberg et al. 2008). Variant A differs at only one aa from the E, F2, G, H and I variants, and variant B is different from C and J at positions 97 and 155, respectively. Thus, they can be grouped as A-related variants (comprising E, F2, G, H and I variants) and/or B-related variants (C and J variants). As shown in Figure 1, the procedure begins with the genotyping of the samples with PCR-RFLP using the Taq I enzyme, which allows the distinction of A or B-related variants. Then each group of samples can be simultaneously genotyped using the primers and enzymes specific for determination of the final variant group, as shown in Figure 1. The procedure requires a minimum of three and a maximum of five additional PCR-RFLP reactions per each sample to determine the specific variant group of each k-casein variant. The strategy uses only seven different primers and seven enzymes, making the strategy cost and time effective for genotyping (Figure 1). It is important to note that for three of the variants, an Artificial Created Restriction Site strategy was applied using only one primer (forward or reverse). The primer design for those cases was based on the published Bos taurus K-casein gene sequence (X14908) using WatCut software. The strategy was tested in animals from three cattle breeds. Whole blood samples were collected from 41 Gyrholando animals and 105 Charolais animals. The sampled ranches are located in Sonora and Tamaulipas, Mexico, and their cattle are characterized as purebred cattle. DNA Isolation from blood samples was conducted as is recommended by the Wizard® Genomic DNA Purification Kit (Promega, Inc.). We also include thirty-five Carora breed hair samples; these samples were obtained from a Cattle Improvement Program in Venezuela, and DNA was isolated using the GeneEluteTM Mammalian Genomic DNA Miniprep Kit (Sigma-Aldrich, USA.). No additional data in terms of productive traits and or origin were collected for the sampled animals All PCR reactions used 50-100 ng genomic DNA, 0.5 µM of each primer, 0.4 mM of dNTPs and 1.5 mM of MgCl2 (for C and H variants, a 2.5 mM MgCl2 was used). The amplifications were performed using touchdown PCR with an annealing temperature of 62ºC for the first cycle, which was then decreased by 2ºC every cycle until 55ºC. Amplification products were resolved by electrophoresis in 1.5% agarose gels stained with Syber Gold 1X and visualized under UV light. The PCR products were digested using standard protocols under the conditions suggested by Promega and incubated at 37ºC for 2 hrs. The digested fragments were electrophoresed in 3% standard agarose and/or 4.5% NuSieve gels stained with Syber Gold 1X and visualized under UV light. Determination of each sample’s genotype was achieved by visual analysis of the gels of the nine variants detected by PCR-RFLP. Estimates of the allele frequencies of K-casein variants in the three populations were determined by direct count, using the following equation:
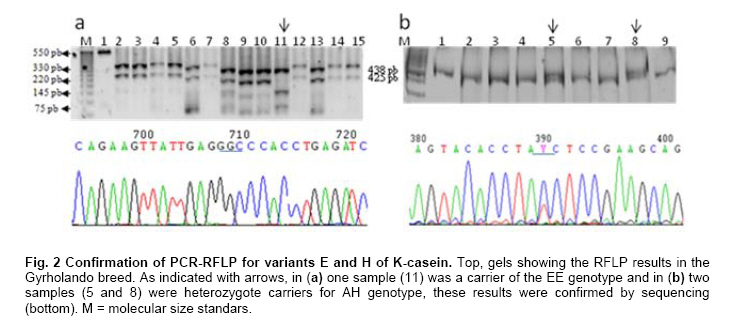
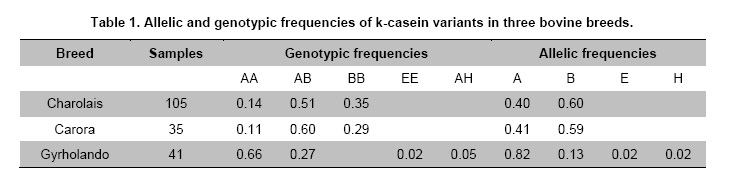
where j ≠ i and n is the number of variants in locus K-casein (Hedrick, 2005). When it was necessary, selected samples were sequenced using the BigDye® Terminator procedure and the DNA sequencer ABI PRISM 3100 Genetic Analyzer (Perkin-Elmer, U.S.A.). For sequence-based genotype determination, sequences were aligned with ClustalX 2.0.8 using the reported Bos taurus K-casein gene sequence (X14908) (Larkin et al. 2007). Results and DiscussionK-casein is of major importance among milk proteins as its allelic variants have been associated with different quality and production traits in the dairy industry. Typing studies of bovine K-casein have mainly been focused on the detection of variants A and B. Because of this, they are considered the most frequent variants in cattle, but this could be due to the lack of evaluation of other variants in these studies. Thus, there is information missing about the real frequencies and effects on productive traits of the K-casein variants in cattle. Several methods for genotyping have been used; however, none have allowed the simultaneous detection of up to six of the K-casein variants. We designed a simple strategy to genotype nine of the variants using PCR-RFLP. No technical modifications to the strategy were made during the genotyping process, and genotype determination was achieved in all animals of the tested population. The results of the first PCR-RFLP (Figure 1) allowed us to cluster 49 samples as A-related variants and 132 samples as B-related variants. Each group of samples was typed according to the strategy with five and two additional PCR-RFLP procedures per sample. Four variants, A, B, E and H, were detected in the studied populations. Due to the low frequency of the last two variants, samples carrying these genotypes were sequenced. The presence of a GGC codon encoding glycine at position 155 and the ATC codon encoding for isoleucine at position 135 confirmed that these genotypes belong to the E and H variants, respectively, thus confirming the PCR-RFLP results (Figure 2). As shown in Table 1, the allelic and genotyping frequencies of the K-casein variants were determined and compared with those previously reported for the three studied cattle breeds. For the Carora breed, the previously reported frequencies were 0.261, 0.487 and 0.252 for genotypes AA, AB and BB, respectively (Rojas et al. 2009). The frequencies of the studied Carora population showed minimal difference from those previously reported; this could possibly be due to the fact that the samples analyzed are considered elite animals, and a high frequency of efficient milk production and high milk quality alleles could be expected. In the Gyrholando breed, we found similar frequencies of A, B and E alleles as previously reported, with genotypic frequencies close to the reported values of 0.64, 0.26 and 0.10 for genotypes AA, AB and AE, respectively (Azevedo et al. 2008). However, an important finding was the presence of variant H, which had not previously been reported in the Gyrholando breed, even when previously reported populations tested more individuals than those included in our study. Therefore, this confirms the novelty of our detection strategy as it allows the detection of those variants of the bovine K-casein gene that have not previously been genotyped. Due to the low frequency of the E and H variants, there are currently no reports about their effects on milk traits, but it is important to promote their inclusion in genotyping studies for the k-casein gene. K-casein variants have been studied mainly in dairy breeds, and to our knowledge, there is little information about the effects of k-casein variants in beef cattle. Lara et al. 2002 suggest that k-casein variants could have an important role in the growth and maternal effects in beef breeds. However, Biase et al. 2005 indicate that a kappa-casein polymorphism did not influence weight-gain in Brazilian Nellore herds, although they found differences in the frequency of the A and B alleles, with the A allele observed most frequently in this beef breed. Similar results were reported by Curi et al. 2005 in Nellore, Canchim and a crossbreed (Simmental x Nellore, Angus x Nellore) cattle. Here, we include animals from the Charolais breed, and we found frequencies similar to those reported by Jann et al. (2004). Indeed, as previously reported for breeds with Bos taurus background, the B variant was the most frequent in the Charolais breed. As the B variant has been associated in the Herford breed with low birth weight and weaning at 180 days, further studies should be performed to provide more information on the effects of k-casein polymorphisms in taurine beef cattle. Introduction of molecular methods to assist animal selection is a current goal in cattle production systems. K-casein is a strong candidate gene for MAS in dairy and beef cattle. Depending on the breed and population investigated, the B and A alleles have so far been found to affect both milk yield and the components of the milk, but studies must be performed to define how other gene variants contribute to milk traits. Most of the reported studies focused on the previously reported k-casein variants in different cattle breeds had used techniques developed to detect only two, four or six K-casein variants (Soria et al. 2003; Chessa et al. 2007; Naranjo et al. 2007; Azevedo et al. 2008). We designed and implemented a strategy based on PCR-RFLP to detect nine k-casein variants in cattle. These variants are characterized by the presence of non-synonymous aa. Amplification coupled to restriction fragment length polymorphism analysis and its variants (i.e., PCR-ACRS) is a good option for most laboratories equipped with basic molecular biology instruments. Although RFLP can, in some instances, result in significant false positive rates as a result of incomplete restriction enzyme digestion or the presence of other mutations close to the mutation of interest, when the procedure is well established, it is a highly sensitive, specific, cost-effective, and straightforward assay for SNP screening (Agarwal et al. 2008). AcknowledgmentsAuthors thanks to the different herds-owners for supporting the cattle sampling. References
Copyright 2011 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej11049t1.jpg] [ej11049f2.jpg] [ej11049f1.jpg] |
| |||||||||

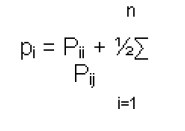
{kind=link}
{kind=link}
{kind=link}