 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 1, No. 2, August, 1998 Invited Review Article Golgins: coiled-coil-rich proteins associated with the Golgi Complex Edward K.L. Chan*1 and Marvin J. Fritzler2
1W.M. Keck Autoimmune Disease
Center, Department of Molecular and Experimental Medicine, The Scripps Research
Institute, 10550 North Torrey Pines Road, La Jolla, California, 92037 Tel: 619-784-8250
Fax: 619-784-2131 E-mail:echan@scripps.edu
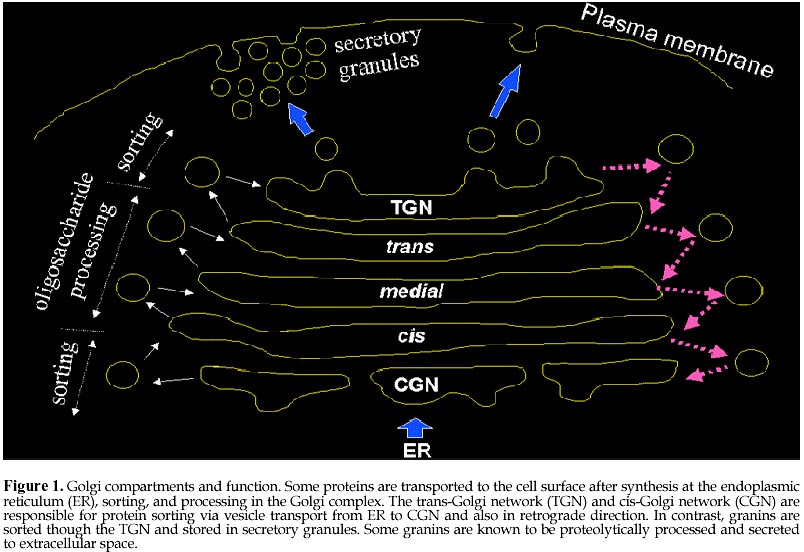
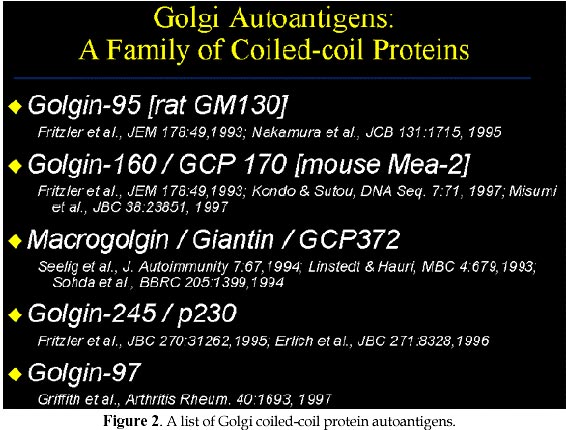
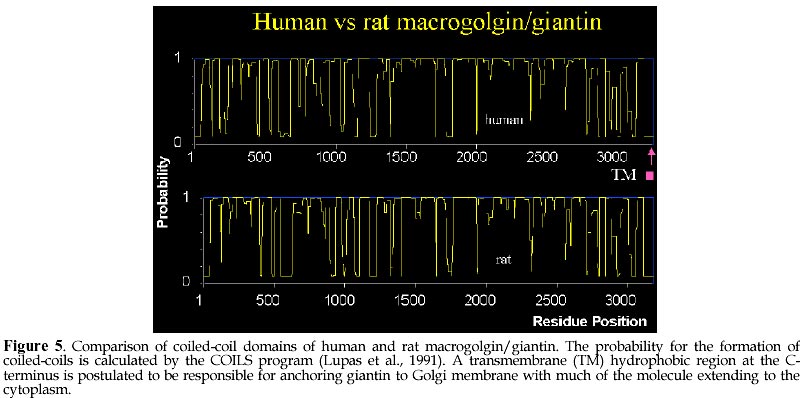
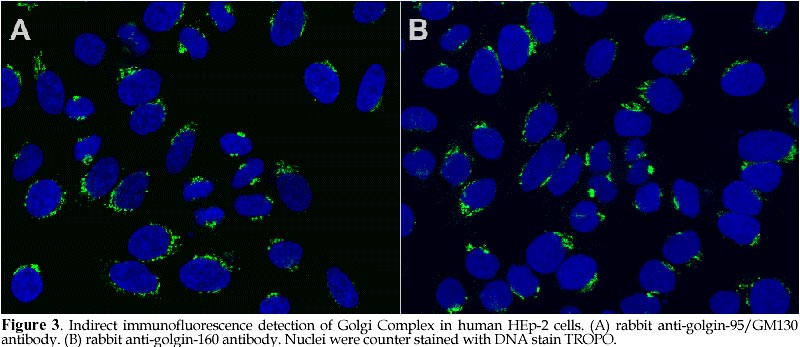
Code Number: ej98008 Autoantibodies directed against self-antigens are characteristic features of a number of human diseases. In systemic rheumatic diseases, these include antibodies that react with extracellular molecules; the cell membrane; nuclear and cytoplasmic components. Our laboratories have been interested in understanding autoimmune responses in systemic lupus erythematosus and related disease states and using human autoantibody as molecular probes in studying the cell biology of target antigens. This review summarizes our research efforts in dissecting the autoimmune response to the Golgi complex via expression cloning using human autoantibodies. We have cloned 4 of 5 known Golgi autoantigens that are referred to as golgin-95/GM130, golgin-97, golgin-160/GCP170, and golgin-245, respectively based on their molecular weights. The fifth is giantin, also known as macrogolgin or GCP372. Interestingly, the amino acid sequences deduced for all Golgi autoantigens indicate that they are proteins with predominantly coiled-coil domains and non-alpha helical domains at the N- and C-termini. The current postulate is that these coiled-coil rich proteins like golgin-95/GM130 form intermolecular complexes with a docking protein p115 that has been shown to be important for Golgi vesicular traffic. An alternative nonexclusive hypothesis is that these coiled-coil rich proteins are the components of the "strings" that are important in linking and guiding vesicles in intercisternae traffic. How this family of coiled-coil proteins and their complexes become autoimmune targets remains to be determined. Keywords: Autoimmunity, Autoantibodies, Antigens Patients with certain autoimmune diseases are characterized by the spontaneous occurrence of autoantibodies directed against a variety of tissue antigens (Tan, 1989). These include cell membrane components such as hormones and hormone receptors, immunoglobulins, proteins involved in fibrinolysis, complement proteins, and intracellular cytoplasmic and nuclear components. Some of the target antigens are tissue specific, while others are variably expressed in differentiated and undifferentiated tissues. The studies of many intracellular autoantigens illustrate a number of important features concerning the clinical importance of human autoantibodies. First, certain autoantibodies seem to be restricted in one or a few diseases and, therefore, have useful diagnostic and prognostic implications. Second, patterns of end organ involvement are sometimes related to the presence of certain autoantibodies suggesting that they may have pathogenic significance. An equally important outcome of studies of autoantigens has been the tremendous increase in our understanding of their molecular and cell biology (Tan, 1989; Tan et al., 1988). First, the majority of autoantibodies studied have been shown to bind to highly conserved determinants on ubiquitous cellular proteins. Second, most of the autoantibodies associated with systemic rheumatic diseases are often directed to functional macromolecules rather than to structural components. These include histones, DNA and HMG of the nucleosome, several proteins of the small nuclear ribonucleoprotein (snRNP) complex, various components of the mitotic apparatus including centrosomes, kinetochore and centromere (CENPs) components, components of the nucleolus and the nuclear membrane. Third, where systems are amenable to testing, it has been shown that the autoantibodies are able to inhibit the cellular functions served by the antigens. Examples include the inhibition of aminoacylation of tRNAs; the relaxation of supercoiled DNA; inhibition of precursor mRNA splicing; and the transcription of ribosomal RNA. Taken together, these observations suggest that the conserved epitopes recognized by human autoantibodies are often the functional or active sites of these intracellular proteins. The study of human autoantibodies and their use as probes of cell structure and function has had an important impact on the disciplines of molecular and cell biology (Tan, 1991). For immunologists, one of the interesting objectives is the identification of macromolecules and organelles, such as the Golgi complex, that are targets of the autoimmune response. For cell biologists, attempts to unravel cellular events can be enhanced by the availability of specific autoantibody probes. The Golgi compartmentIn 1898, exactly 100 years ago, Camillio Golgi described a novel intracellular network which now bears his name (Golgi, 1898). The Golgi complex is an elaborate cytoplasmic organelle that has a prominent function in the processing, transporting, and sorting of intracellular proteins (reviewed in Gonatas, 1994; Mellman, 1995; Nilsson and Warren, 1994). Structurally, the Golgi complex is localized in the perinuclear region of most mammalian cells and is characterized by stacks of membrane-bound cisternae as well as a functionally distinct trans- and cis-Golgi networks (TGN, CGN, Figure 1). It is proposed that the sorting functions of the Golgi complex are performed in TGN and CGN while the processing functions take place in the cis-, medial-, and trans-compartments (Mellman and Simons, 1992). The intracellular transport of newly synthesized proteins requires directed movement from the endoplasmic reticulum (ER), via transport vesicles to the cis-, medial- and trans-compartments of the Golgi complex, and in some cases, to the plasma membrane (Banfield et al., 1994; Farquhar and Palade, 1981; Griffiths et al., 1989; Mellman, 1995; Nilsson and Warren, 1994; Rothman and Orci, 1992). Coatomer proteins COPI-coated vesicles are currently understood to mediate this anterograde transport across the intervening cisternae (Rothman, 1994; Schekman and Orci, 1996). Protein transport through the Golgi complex is mediated by small vesicles budding from a donor membrane and are targeted to, and fused with, an acceptor membrane (Rothman and Orci, 1992). Transport vesicles are known to move towards the TGN and are also hypothesized to move in the 'retrograde' direction to the CGN via the coat protein complex (coatomer proteins, e.g. beta-COPs, ref. (Banfield et al., 1994; Barlowe et al., 1994; Duden et al., 1991; Orci et al., 1997; Pelham, 1994; Seaman and Robinson, 1994; Serafini et al., 1991; Waters et al., 1991). The pathways for the vesicular transport are shown schematically in Figure 1 and are thought to be important for the recycling of these membranous structures. The signals that control the vesicular traffic are poorly understood although it is known that intracellular microtubules are important components (Kreis, 1990; Mizuno and Singer, 1994). Other proteins of the Golgi complex believed to play a role include families of proteins such as the adaptins (Pearse and Robinson, 1990), GTP-binding proteins (Jena et al., 1994; Martinez et al., 1994; Nuoffer et al., 1994; Oka and Nakano, 1994; Pfeffer, 1994), ADP ribosylation factors (ARFs) (Stearns et al., 1990), and resident enzymes (reviewed in (Farquhar, 1985; Nilsson and Warren, 1994). A rat peripheral membrane protein p115/TAP has been identified as a component necessary for intra-Golgi transport (Barroso et al., 1995; Waters et al., 1992) and is proposed to be the mammalian homolog of Uso1 a protein required for endoplasmic reticulum to Golgi vesicular transport in Saccharomyces cerevisiae (Sapperstein et al., 1995). Autoantigens of the Golgi compartmentAutoantibodies directed against the Golgi complex were first identified in the serum of a Sjögren's syndrome (SS) patient with lymphoma (Rodriguez et al., 1982). Several isolated reports have described anti-Golgi antibodies in systemic rheumatic diseases including SS and systemic lupus erythematosus patients (Blaschek et al., 1988; Fritzler et al., 1984; Hong et al., 1992; Kooy et al., 1994; Kooy et al., 1992; Renier et al., 1994; Rodriguez et al., 1982; Rossie et al., 1992), and in a variety of other disease conditions including idiopathic Raynaud's phenomenon (Gentric et al., 1990), idiopathic cerebellar ataxia (Fritzler et al., 1993; Gaspar et al., 1988), active Wegener's granulomatosis (Mayet et al., 1991), paraneoplastic cerebellar degeneration (Greenlee et al., 1988; Hida et al., 1994), stiff man syndrome (Butler et al., 1993), and viral infections including the Epstein Barr virus (EBV) (Huidbuchel et al., 1991), hepatitis B (Funaki et al., 1996), and the human immunodeficiency virus (HIV) (Gentric et al., 1991). Immunoblotting and immunoprecipitation studies have shown that there are heterogeneity of reactivities among anti-Golgi sera with at least 14 different Golgi complex autoantigens ranging from 35 to 260 kDa (Kooy et al., 1994; Renier et al., 1994). Immunofluorescence studies suggest that, like many other autoantigens, Golgi autoantigens are evolutionarily conserved and autoantibodies are able to recognize homologous proteins across species (Fritzler et al., 1993). Within the last several years, our laboratories and others have cloned and identified five novel Golgi autoantigens (Table 1, Figure 2). This has been achieved primarily by expression cloning using human autoantibody probes. These are referred to as golgin-95 (GM130), golgin-97, golgin-160, golgin-245 (p230), and giantin/macrogolgin/GCP372 respectively based on their molecular weights estimated from SDS-PAGE under denaturing condition. Interestingly, the amino acid sequences deduced for all golgins show that these proteins are composed of predominantly alpha-helical coiled-coils except for the N- and C-terminal domains (Figure 4 and 5). The potential implication is that these Golgi proteins may have common biochemical characteristics that are responsible for inducing autoimmune response in certain disease states. Golgin-95/GM130 and golgin-160/GCP170The first two Golgi complex autoantigens were cloned in our laboratory using autoantibodies from a systemic lupus erythematosus patient (Table 1, Figure 3). These Golgi complex autoantigen cDNAs encode proteins of 95 and 160kDa, and thus termed golgin-95 and golgin-160 respectively (Fritzler et al., 1993). Sequence analysis showed that golgin-95 and golgin-160 are composed of alpha-helical coiled-coil domains sharing 43% overall sequence similarity and, therefore, may be functionally related proteins (Figure 4). Subsequent work from the isolation of rat liver Golgi matrix proteins, Nakamura et al (Nakamura et al., 1995) have shown that a 130kDa protein (GM130) may be the rat homolog of golgin-95. Our recent data confirmed that a ~130kDa protein is detected in HeLa cells using antibody to the previously observed 95kDa protein; thus our new data is consistent with the mol wt reported for GM130 and the 95kDa protein was most likely a degradation product of GM130. A recent report showed that the N-terminal 73 amino acid residues of GM130 interacts with the vesicle docking protein p115 and therefore may play a role in vesicular traffic between ER and Golgi stacks (Nakamura et al., 1997). In addition, the same laboratory reported that GM130 also interact strongly with GRASP65, a Golgi reassembly and stacking protein of 65kDa involved in the reformation of Golgi Complex after mitotic cell division (Barr et al., 1997). A mouse testis protein known as Male-Enhanced Antigen-2 (Mea-2) originally identified with a monoclonal histocompatibility Y (H-Y) antibody was cloned and found to be highly homologous to human golgin-160 (Kondo and Sutou, 1997). Although conclusive data is not presented, some role for golgin-160 in spermatogenesis is postulated based on the high level of expression of this gene in testis. Recently, the laboratory of Ikehara (Misumi et al., 1997) reported the cloning of a full-length cDNA by extending from our earlier reported partial sequence of golgin-160. The group of Ikehara has named this protein GCP170 based on their estimated mol wt for this protein. It should be noted that the full-length protein contains an additional 160 proline-rich amino acids at the N-terminus compared to Mea-2. Based on these observations, Misumi et al (Misumi et al., 1997) postulated the full-length protein has an N-terminal globular head, a long stalk composed of coiled-coil domains, and a C-terminal short tail. It was also shown that GCP170 is localized to the cytoplasmic face of the Golgi membrane but not associated with beta-COP subunits of coatomers. Giantin/macrogolgin/GCP372The third autoantigen of the Golgi complex was identified by Seelig et al as macrogolgin (Seelig et al., 1994a). It was so named because of its large size (376kDa) and subsequent studies showed that macrogolgin is identical to a previously described large protein known as giantin (Linstedt and Hauri, 1993; Seelig et al., 1994b). In 1994 Ikehara and his colleagues reported a 372kDa Golgi complex autoantigen known as GCP372 (Sohda et al., 1994). When sequence alignments were performed, it was apparent that GCP372 has an extra 5 amino acids (QLSSM) "inserted" at residue 215 of the giantin amino acid sequence. Thus GCP372 could be derived from an alternatively spliced mRNA transcribed from the same gene as giantin/macrogolgin. This same laboratory (Toki et al., 1997) recently reported the cloning of the rat homolog GCP364 and, in agreement with other earlier studies, concluded that giantin is anchored to the Golgi membrane via C-terminal hydrophobic domain with the coiled-coil domain extending to the cytoplasm (Figure 5); it is also postulated that this molecule may be involved in the formation and/or maintenance of the Golgi structure. Recently, Sonnichsen et al (Sonnichsen et al., 1998) proposed that giantin interacts with p115 which in turn binds to GM130 on Golgi membranes to promote vesicle traffic. Golgin-245/p230Using a serum from a Sjögren's syndrome patient who had glomerulonephritis, the fourth golgin cloned was a 245kDa protein, named golgin-245 (Fritzler et al., 1995). This protein was also characterized by a dominant coiled-coil domain and a granin signature motif found in a family of proteins known as granins. Granins are acidic proteins present in the secretory granules of a wide variety of endocrine and neuroendocrine cells. About the same time, another group described the nucleotide sequence from a previously reported a 230kDa Golgi complex autoantigen (p230) which is now known to be identical to golgin-245 (Erlich et al., 1996; Kooy et al., 1992). The study of p230 described alternatively spliced products suggesting that there may be further heterogeneity of this Golgi complex antigen (Erlich et al., 1996). A third group of investigators has recently reported the cloning of the same protein using the serum from a patient with hepatitis B (Funaki et al., 1996). Golgin-97The fifth Golgi autoantigen cDNA cloned with the serum from another Sjögren's syndrome patient was shown to encode a protein of 97kDa (golgin-97). Like golgin-245, golgin-97 has a predominant coiled-coil domain and a granin signature sequence that implicates its potential role associated with secretory granules. Among all the cloned Golgi complex autoantigens analyzed to date, golgin-97 appears to be the most common target associated with Sjögren's syndrome (Griffith et al., 1997). Although anti-Golgi autoantibodies are rare in Sjögren's syndrome patients, their detection may represent a subgroup of patients with interesting defects associated with Golgi function. Given that the sera used for the cloning of both golgin-245 and golgin-97 came from patients with SS and the existence of other reports of anti-Golgi antibodies detected in patients with SS, it is intriguing to consider that these autoantibody-autoantigen systems play a significant role in the secretory defects in Sjögren's exocrine glands (Table 1). Coiled-coil domains of golginsThe golgins are related as they have similar overall secondary structures as predicted by analysis programs such as COILS (Lupas et al., 1991) and PAIRCOIL (Berger et al., 1995). Figures 4 and 5 show the prediction of coiled-coil domains calculated by COILS for each of the five Golgi autoantigens. It is interesting that the golgins also show significant sequence similarity to several cytoskeleton-related proteins including kinesin (Bloom and Brashear, 1989), the 150 kD dynein-associated protein (Holzbaur et al., 1991), the myosin family proteins (heavy chain myosin, tropomyosin), and desmin. Kinesin, a microtubule-stimulated ATPase (Bloom et al., 1988; Kuznetsov and Gelfand, 1986), has been reported to be a motor for microtubule-mediated Golgi-to-ER membrane traffic (Lippincott-Schwartz et al., 1995). In support of this observation, it has been demonstrated that antibodies to kinesin and myosin bind to the Golgi complex (Fath and Burgess, 1993; Fath and Burgess, 1994; Marks et al., 1994). To extend the comparison, golgin-160 and macrogolgin have sequence similarity to the yeast cytoskeleton-related protein Uso1, a yeast protein which is required for protein transport from the endoplasmic reticulum to the Golgi apparatus (Nakajima et al., 1991). The human autoimmune response to golgins appears to be highly specific as many anti-Golgi sera react with only one of these autoantigens. It is therefore unlikely that the immune response is merely directed to cross-reactive the coiled-coil region in these molecules. In addition, coiled-coil domains have been noted in many other non-Golgi autoantigens including nuclear mitotic apparatus protein NuMA (Andrade et al., 1996; Price et al., 1984), lamin B (Chou and Reeves, 1992; Konstantinov et al., 1996; Pollard et al., 1990), myosin heavy chain (von Muhlen et al., 1995), 52 kDa SS-A/Ro (Buyon et al., 1994; Chan et al., 1991), and 70/80 kDa Ku antigens (Chou et al., 1992; Reeves et al., 1991). Granin signature in golgin-97 and golgin-245Granins are a family of acidic proteins present in the secretory granules of a wide variety of endocrine and neuroendocrine cells (Huttner et al., 1991; Simon and Aunis, 1989). Two consensus sequences have been reported. The granin 1 signature is located at the carboxyl terminus of the proteins and has been identified in all granin family proteins with the exception of murine secretogranin 2. The second consensus sequence, granin 2, has been described in chromogranin A and B and is characterized by two cysteine residues bound together near the amino terminus of the protein. The acidic charge of these proteins has been attributed to high content of glutamic and aspartic acid. The function of these proteins or these signatures has not been clearly defined but current evidence suggests that they function in modulator-processing or packaging of neuropeptides. In addition, granins such as chromogranins and secretogranins are precursors that are proteolytically processed to biologically active secretory peptides (Natori and Huttner, 1994). Decapeptides ESLALEELEL and ESLGKMEQEL corresponding to amino acids 388-397 in golgin-245 and amino acids 193-202 in golgin-97 were identified as a granin 1 'signature' which has the consensus pattern {DE}-{SN}-L-{SAN}-x(2){DE}xEL (Huttner et al., 1991; Simon and Aunis, 1989). The predicted isoelectric points (pI) for golgin-245 and golgin-97 are 5.15 and 5.09 respectively which are similar to those reported for granins (pI 4.9-5.6). The anionic pIs are consistent with the high content of glutamic and aspartic acid of these proteins. Golgin-245 has 17% and 4% while golgin-97 has 14% and 3% of glutamic and aspartic acid respectively. These features are consistent with those reported for granins. Recent review of available golgin-95/GM130 and golgin-160/GCP170 sequences showed no evidence for granin signature even when one mismatch is allowed. The significance of identifying granin signature may become apparent when immunohistocytochemical data is available for golgin-97 and golgin-245 at the electron microscopy level. Proposed function for golginsThe observation that all Golgi complex autoantigens cloned to date have predominantly central rod-like alfa-helical coiled-coil structures and non-helical termini may be a clue to their function. Recent publication by Orci, Perrelet and Rothman (Orci et al., 1998) proposes a "string theory" for the organization of Golgi vesicles primarily based on their electron microscopy and freeze-fracture analysis of Golgi fractions purified from Chinese hamster ovary cells. The interpretation is that transport vesicles remain locally within the Golgi stack during their lifetime, near their origin, favoring a processive pattern of transport in which vesicles transfers occur preferentially between adjacent cisternae in the stack (Orci et al., 1998). The proposed theory accounts for the processive transport by postulating that vesicles are linked by "strings" which will restrict and keep the vesicles in close proximity. It is highly interesting and, of course, entirely reasonable to propose that these strings are composed of macromolecules rich in coiled-coil domains like GM130 and giantin (Orci et al., 1998). The string theory postulates a predominate role for the coiled-coil rich Golgi proteins in holding transport vesicles in place. An additional function for this family of proteins may be in guiding vesicular traffic via specific interactions programmed into individual members of the coiled-coil rich protein family. Recent studies by Warren and his colleagues demonstrated the interaction of p115 with giantin and GM130 and thus p115 may serve as a bridge for COPI vesicles and Golgi membrane (Sonnichsen et al., 1998). In addition, the data from the same laboratory suggest that protein like golgin-95/GM130 are directly associated with GRASP65, which is important in the stacking of Golgi cisternae (Barr et al., 1997). In summary, based on molecular sequence data, several Golgi autoantigens are defined as proteins rich in alpha-helical coiled-coil domains with non-helical termini. These represent a subfamily of proteins that may play structural roles in supporting vesicles associated with the Golgi complex as well as stacking of Golgi cisternae. The rod domains are likely to participate in self-association as well as interaction among different family members. Their roles in vesicular traffic may soon become apparent as they are being actively investigated in many laboratories. References
Abbreviations: SLE=systemic lupus erythematosus. 1Diagnosis of patient whose antibody was used for screening cDNA library. Supported by UNESCO / MIRCEN network © 1998 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej98008f5.jpg] [ej98008f4.jpg] [ej98008f2.jpg] [ej98008f1.jpg] [ej98008f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}