
|
Electronic Journal of Biotechnology
Universidad Católica de Valparaíso
ISSN: 0717-3458
Vol. 1, Num. 3, 1998, pp. 103-117
|
Electronic Journal of Biotechnology, Vol.
1, No. 3, December, 1998
REVIEW ARTICLE
Medium-term and long-term in
vitro conservation and safe international exchange of yam (Dioscorea
spp.) germplasm
Bernard Malaurie*1, Marie-France
Trouslot2, Julien Berthaud3,
Mustapha Bousalem4, Agnès Pinel5
and Jean Dubern6
1GeneTrop – Unité de Génétique
et d’Amélioration des Plantes - Centre ORSTOM** – 911 avenue Agropolis
- BP. 5045 -F. 34032. Montpellier Cedex 1, France. Tel: Bureau
(33)-- 4 67 41 62 44.Standard (33)-- 4 67 41 61 00 Fax: (33)-- 4 67 54 78 00
E-mail : Bernard.Malaurie@mpl.orstom.fr
2GeneTrop – Unité
de Génétique et d’Amélioration des Plantes - Centre ORSTOM**
– 911 avenue Agropolis - BP. 5045 -F. 34032. Montpellier Cedex 1, France.
3GeneTrop – Unité de Génétique
et d’Amélioration des Plantes - Centre ORSTOM** – 911 avenue Agropolis
- BP. 5045 -F. 34032. Montpellier Cedex 1, France.
4Laboratoire de Phytovirologie des
Régions Chaudes (LPRC) – CIRAD-ORSTOM** – avenue du val de Montferrand
- BP. 5035 -F. 34032 - Montpellier Cedex 1, France.
5Laboratoire de Phytovirologie des
Régions Chaudes (LPRC) – CIRAD-ORSTOM** – avenue du val de Montferrand
- BP. 5035 -F. 34032 - Montpellier Cedex 1, France.
6Laboratoire de Phytovirologie des
Régions Chaudes (LPRC) – CIRAD-ORSTOM** – avenue du val de Montferrand
- BP. 5035 -F. 34032 - Montpellier Cedex 1, France. E-mail : dubern@melusine.mpl.orstom.fr
http://www.mpl.orstom.fr
*Corresponding author
**ORSTOM is now called IRD, Institut de Recherche pour le Developpement.
Code Number: ej98015
Abstract
Yam edible tubers feed million of peoples
in the intertropical area, where they represent 12% of human feeding. However,
as a vegetatively propagated crop, yam is seriously affected by an accumulation
of pathogens. Establishing in vitro germplasm collection is a process
that cleans the plants from all diseases but viruses. It gives a good control
on the preservation of the yam genetic resources and facilitates international
exchanges of healthy plant material.
Two kinds of in vitro germplasm preservation
were considered : slow growth condition culture for mid-term preservation,
and cryopreservation using the encapsulation/dehydration technique for long-term
preservation. Virus eradication was approached by meristem culture and chemo
and thermotherapy. Production of virus-free plants was controlled by ELISA.
We succeeded in the introduction and maintenance
of 20 yam species, under slow growth conditions. Cryopreservation was applied
successfully on two edible yam species, Dioscorea. alata L and
D. bulbifera L. Virus-free plants were obtained by meristem culture in
D. cayenensis-D. rotundata complex and D. praehensilis. Indexation
allowed the detection of different virus (poty-, potex-, badna- and cucumovirus),
where the most important potyvirus was YMV.
Mid-term conservation of yam germplasm is
used routinely, and from these conditions a direct acclimatization is possible.
On the cryopreservation aspect, experiments are under way to apply the optimized
protocol to genotypes which are more representative of the diversity, to insure
a routinely use. More work can be conducted now on virus eradication, based
on knowledge accumulated on potyvirus diversity, on several tests available
for yam indexing (ELISA, rt/PCR, monoclonal antibodies) and on new sanitation
techniques.
Keywords: Active and base in vitro
genebanks’, Chemotherapy, Cryopreservation, Disease-free techniques, Indexation
techniques, Slow growth condition culture, Virus eradication, Yam viruses.
Article
Yam belongs to
Dioscorea genus which has more than 600 species (Coursey,
1967) most of them distributed in the intertropical humid area. We will
distinguish two types of yam: 1) medicinal yams, 2) edible yams and relatives.
Medicinal yams concern about fifty species caracterized by their sapogenin content,
which are steroidal components. For the edible yams and relatives, we will observe
two groups: 1) domesticated species, 2) and wild species.
For domesticated species we consider that
forty to fifty species are occasionally used (Martin and Degras,
1978). From these only eleven are cultivated (Table 1).
From the 11 cultivated, 6 represent an important part of feeding (D. alata,
D. cayenensis-D. rotundata complex, D. bulbifera, D. dumetorum, D. esculenta,
D. trifida), 3 are scattered in all the intertropical humid area (D.
alata, D. bulbifera, D. esculenta), and yam belonging to the D. cayenensis-D.
rotundata complex take place most of them in West Africa and some in the
Caribbean area. The other yams are cultivated in their origin area (Degras,
1986).
Table 1 .
Main edible species of yam
|
Species 1
|
Zone of origin
|
Zone of culture
|
| |
|
|
|
Enantiophyllum Section
|
|
|
|
D. alata L.
|
South East Asia
|
Inter-tropical humid
|
|
D. cayenensis Lamk.
D. rotundata Poir.
complex 2
|
West Africa
|
West and Central Africa, and Caribbean
|
|
D. nummularia Lamk.
|
Indonesia, Oceania
|
Indonesia, Oceania and
Micronesia
|
|
D. opposita Thunb.
D. japonica Thunb.
complex 3
|
Temperate area from:
China, Corea, Taiwan
Japan
|
Temperate area from:
China, Corea, Taiwan
Japan
|
|
D. transversa Br.
|
South Pacific
|
South Pacific
|
|
Lasiophyton Section
|
|
|
|
D. dumetorum (Kunth) Pax.
|
West Africa
|
West Africa
|
|
D. hispida Dennst.
|
India, South-China, New Guinea
|
India, South-China, New Guinea
|
|
D. pentaphylla L.
|
Himalaya and Oceania
|
Himalaya and Oceania
|
|
Combilium Section
|
|
|
|
D. esculenta (Lour.) Burk.
|
South East Asia
|
Inter-tropical humid
|
|
Opsophyton Section
|
|
|
|
D. bulbifera L.
|
South East Asia and Africa
|
Inter-tropical humid
|
|
Macrogynodium Section
|
|
|
|
D. trifida L.
|
Guyana, Amazonian basin
|
Caribbean
|
Sources: Malaurie et al. (1998a)
1 Species have been regrouped in Section
by Knuth (1924), completed by Burkill
(1960)
2 Grouping together species of D. cayenensis and
D. rotundata in a Complex has been proposed by Ayensu
and Coursey (1972), Martin and Rhodes (1978),
Miège (1982)
3 Grouping together species of D. opposita and D.
japonica in a Complex has been proposed by Tanaka
(1977).
Main edible species of yams are: 1) native
of a continent, 2) and cultivated in the same continent, 3) or/and cultivated
in an other. This observation implies very strong links to exchange problems.
In Table 2, 36 countries have been observed by IBPGR
in 1986 with Dioscorea germplasm. These countries are supposed to be
concerned by an international exchange of yam germplasm. Some of them, have,
in our knowledge, already developed in vitro germplasm collection.
Table 2 .
Countries¹ and geographic zones where yam collections have been observed
|
Europe
|
West Indies
|
America
|
Pacific
|
Asia
|
Africa
|
| |
|
|
|
|
|
|
France 2
|
Barbados *
|
Brazil 2
|
Cook Islands
|
Bengladesh
|
Bénin
|
| |
|
|
|
|
|
|
United Kingdom 2
|
Cuba
|
Colombia
|
Fiji
|
India
|
Burkina Faso
|
| |
|
|
|
|
|
| |
Guadeloupe 2
|
Costa Rica
|
Niue Islands
|
Indonesia
|
Cameroun
|
| |
|
|
|
|
|
| |
Jamaïque
|
Guatemala
|
Nouvelle Calédonie 2
|
Japan 2
|
Côte d’Ivoire 2
|
| |
|
|
|
|
|
| |
Saint-Domingue
|
Mexico
|
Papua NewGuinea
|
Malaisia
|
Ghana
|
| |
|
|
|
|
|
| |
Trinidad
y Tobago
|
Panama
|
Salomon Islands
|
Nepal
|
Nigeria 2
|
| |
|
|
|
|
|
| |
|
USA
|
Tonga
|
Philippina
|
South Africa
|
| |
|
|
|
|
|
| |
|
|
Vanuatu
|
Sri Lanka
|
Togo
|
| |
|
|
|
|
|
| |
|
|
Western Samoa 2
|
Thailand
|
Uganda
|
| |
|
|
|
|
|
| |
|
|
|
Viet Nam
|
|
(Sources : IBPGR
1986, FAO 1996, Malaurie et al 1998a)
* in vitro maintenance for production purpose
1 This country listing is not exhaustive, and take into account only
sources in our possession
2 Countries with in vitro collection (according to sources
in our possession)
Different genebank preservation levels exist.
A first group concerns non aseptic germplasm conservation with in field genebank
and seed genebank, where important disadvantages and heavy constraint of quarantine
measures explain the choice of in vitro germplasm conservation (Hanson,
1986; Malaurie et al., 1998a) (Table
3).
Table 3. Non
aseptic Germplasm Conservation
| |
Non aseptic Genebanks
|
| |
In field Genebanks
|
Seed Genebanks
|
|
Disadvantages
|
|
|
|
Genetic erosion
|
+++
|
|
|
Expensive
|
+++
|
|
|
Hard to manage
|
+++
|
|
|
Do not bread true
|
|
+++
|
|
Tuber shape
|
|
+++
|
|
Dormancy
|
|
+++
|
In vitro
conservation
Three levels of in vitro genebank
preservation levels could be considered: 1) short term conservation: this conservation
under normal growth conditions is suitable for temporary storage of germplasm
collections, and for international distribution, 2) medium term conservation,
which could be considered as an active in vitro genebank, 3) long term
conservation, considered also as a base in vitro genebank.
These in vitro genebanks have been
previously introduced in vitro from tuber or seed. These introductions
have to be linked to an obligatory phytosanitary control from mother plants
and from in vitro material after introduction. Medium term conservation,
which correspond to in vitro culture under slow growth conditions, could
be obtained by several ways: 1) physiological stage of the explant, 2) addition
of osmotic agents and growth moderators, 3) low storage temperature, 4) low
mineral or sucrose concentrations, 5) low oxygen pressure, 6) encapsulation
in alginate (Charrier et al., 1991; Withers,
1991; Engelmann, 1991; Malaurie et al.,
1998a).
Medium-term conservation
At ORSTOM**, we choose to maintain the in
vitro yam collection in a medium with low mineral nutrient and a low sucrose
concentration. We succeeded in the introduction and maintenance of 14 species
of yam (Malaurie et al., 1993). Since this time, this collection
is continuously enriched by new genotypes and comprises 20 species (Table
4).
For yam, this simple solution of slow growth
is used routinely and from these culture conditions a direct acclimatization
is possible. This mode of conservation allows an international distribution
of the material and corresponds to an active genebank (Malaurie
et al. 1993,1998c, Malaurie and Trouslot
1995c).
This in vitro germplasm collection
of yam is maintained in test tubes, at ORSTOM** (Montpellier, France), with
a total of 6 test tubes by accession, with two different places of storage for
the replicates ; the minimal growth conditions allow to maintain most of
the accessions up to 2 years. Technical constraints in the collection management
lead to subculture the accessions every 6-8 months (Malaurie et
al., 1998c).
Table 4 .
Listing of different species of yam maintained in an in vitro collection,
under slow growth culture condition *
(GeneTrop, GAP unit, ORSTOM**, Montpellier,
France)
|
Species
|
Number of accessions
|
|
D. abyssinica Hochst.
Ex Kunth
|
6
|
|
D. alata L.
|
91
|
|
D. bulbifera L.
|
8
|
|
D. cayenensis Lamk.
D. rotundata Poir.
complex
|
63 (+ 17)
|
|
D. burkilliana J. Miège
|
11
|
|
D. dumetorum (Kunth) Pax.
|
2
|
|
D. esculenta (Lour.) Burk.
|
10
|
|
D. hirtiflora Benth.
|
1
|
|
D. mangenotiana J. Miège
|
15
|
|
D. minutiflora Engl.
|
2
|
|
D. opposita Thunb.
D. japonica Thunb.
complex
|
1
|
|
D. praehensilis Benth.
|
17
|
|
D. preussii Pax
|
1
|
|
D. sansibarensis Pax
|
1
|
|
D. schimperana Hochst.
Ex Kunth
|
1
|
|
D. smilacifolia De Wild
|
2
|
|
D. togoensis Knuth
|
8
|
|
D. transversa Br.
|
1
|
|
D. trifida L.
|
2 (+ 1)
|
|
Interspecific Hybrids: D.
cayenensis-D. rotundata complex cv. ‘Krengle’ X D. praehensilis
|
14
|
|
so-called ‘Igname de Pilimpikou’
|
(+ 9)
|
* (+ ): Accessions recently
introduced
Different species maintained in the in
vitro collection, such as D. cayenensis-D. rotundata complex are
going to be enriched by cultivars from Burkina Faso for a sanitation program,
and from Benin for a genetic program linkeds to virologic aspect. Others species
supposed to be links to D. cayenensis-D. rotundata complex, such as D.
mangenotiana, D. praehensilis, D. minutiflora, or D. abyssinica,
D. praehensilis, are going to be enriched by further introduction.
Orstom** virologists are interested by D.
trifida because of its strong sensibility to virus, which provoked in Guadeloupe,
French West-Indies, its quite disappearance. Serological and molecular works
are developed to explain this virus sensibility.
Long-term conservation
Long term conservation correspond to cryopreservation
in liquid nitrogen, at –196 °C. Plant cryobiology, which begun in 1971 by Latta
works on carrot cell suspension, benefited from results on animals cell by Polge
et al in 1949. Since these dates, different techniques have been set up:
1) on one hand, the so-called conventional techniques, using two steps of slow
freezing, with the addition of cryo-protector (Sakai, 1984),
2) and on the other hand, new techniques, characterized by a very rapid freezing,
about 1000°C/ min, by direct immersion in liquid nitrogen (Table
5) (Dereuddre et al., 1990, 1991; Tannoury
et al., 1991; Uragami, 1993). The aim of these techniques
is to try to control water flow and ice formation, and tend to a vitrificated
state, avoiding crystal formation during thawing, and to protect the cell from
thermic shocks.
Table 5 .
Long term conservation : cryopreservation in liquid nitrogen, -196°C
|
Steps
|
Conventional techniques
|
|
New techniques
|
|
| |
|
Air-drying
|
Vitrification
|
Encapsulation
/ Dehydration
|
|
Encapsulation
|
|
|
|
+
|
|
Sucrose pretreatment
|
+/-
|
+
(+ABA)
|
|
+
|
|
Cryoprotector
|
+
|
|
++++
|
|
|
Desiccation
|
|
+
|
|
+
|
|
Slow-freezing
|
+
0°C to -40°C
(0.3 to 1°C/min)
|
|
|
|
|
Rapid-freezing
|
+
-40°C to -196°C
(200°C/min)
|
+
+25°C to -196°C
(720°C/min)
|
+
+25°C to -196°C
(400 to 1100°C/min)
|
+
+25°C to -196°C
(720°C/min)
|
|
Thawing
|
500°C/min
|
120°C/min
|
120°C/min
|
120°C/min
|
Sources : Uragami (1993),
Malaurie et al. (1998a).
Most of the results about cryopreservation
have been obtained from conventional techniques on suspension cells of medicinal
yam, D. deltoidea being the most used ( Butenko et al.,
1984; Popov and Fedorovskii, 1992; Popov
and Volkova, 1994). More recent works have been done on rapid cryopreservation
of callus (Chulafich et al., 1994), by direct immersion in
liquid nitrogen, of two other medicinal yams (D. balcanica, D. caucasica).
Since 1996, new results have been obtained
by two different research teams, using encapsulation/dehydration of shoot apices.
On the one hand, Mandal et al. (1996) compared the
survival capacities of apices after the osmotic and thermic stress of the technique
of four species of yam - three edible (D. alata, D. bulbifera, D. wallichii),
and one medicinal (D. floribunda). Four of them have survived after immersion
in liquid nitrogen, with 26 to 71%, depending on the species. Meanwhile, only
two of them (D. alata, D. wallichii) allowed the recovery into
shoots after immersion in liquid nitrogen, with 21 and 37%, respectively.
On the other hand, ORSTOM** ability in different
aspects of the long term conservation on tropical plants (Engelmann,
1991), and on encapsulation/dehydration technique applied on coffee, cassava,
oil palm...etc, permitted to apply the process on apical shoot-tips of in
vitro plantlets of yam (Malaurie and Trouslot 1996). Malaurie
et al. (1998b) obtained survival rates over 50% for the two species (D.
alata, D. bulbifera), and recovery to rooted leafy shoots after immersion
in liquid nitrogen of at least 60% for D. bulbifera and 20% for D.
alata, three months culture after thawing.
Comparatively to previous works on cryopreservation
using encapsulation/dehydration technique, Malaurie et al. (1998b)
have used higher sucrose concentration (0.9, 1.0 and 1.1M), a wider range of
dehydration duration, up to 23h and a new and more accurate method for measuring
of dry weight.
The new and more accurate method for measuring
of dry weight used in our experiments consisted of desiccating alginate beads
for 30d in airtight boxes containing dry silica gel, to avoid mass loss due
to caramelization of sugar when drying at a temperature higher than 100°C. We
obtained a strong linear correlation between dry mass (DW30) and
sucrose molarity for sucrose-pretreated alginate beads. During the whole experiment,
we used DW30 values estimated by linear regression (Table
6).
Table 6. Dry mass
and water content of sucrose-pretreated alginate beads, determined
after 30d of drying with silica gel in airtight boxes at room temperature (1)
|
Sucrose concentration
|
DW30 (% FW) estimated
by linear regression (2)
|
Water content before dehydration
(g.g-1 DW)
|
|
0.75M
|
28.8
|
2.47
|
|
0.9M
|
33.3
|
2.00
|
|
1.0M
|
36.3
|
1.76
|
|
1.1M
|
39.3
|
1.54
|
(1)Source: Malaurie et al.
(1998b)
(2)From mean values over 13 to 15 replicates for each of the
four sucrose concentrations
(y= 6.4319 + 29.872x; N= 4; r= 0.999). Similar results were
obtained from replicate data
(y= 6.4177 + 29.883x; N= 55; r= 0.960). Data not shown.
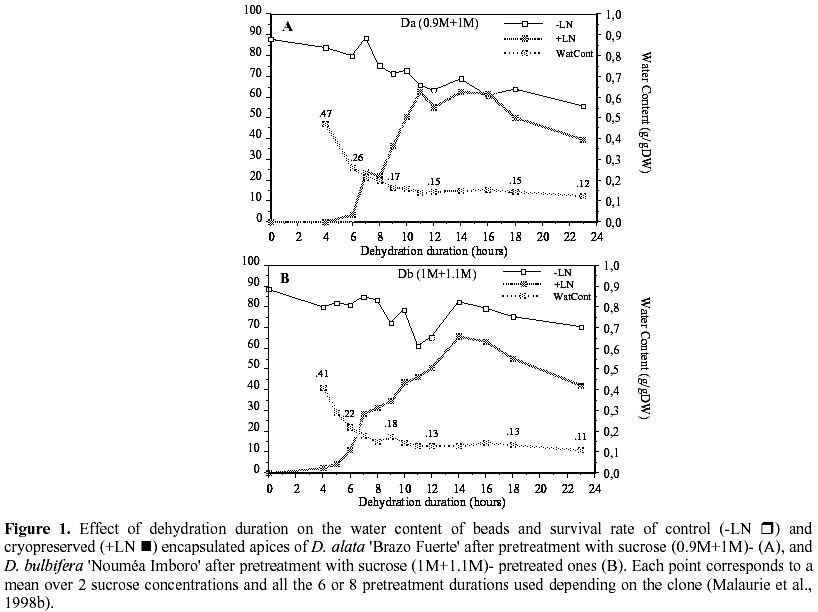
For
the best sucrose pretreatments depending species, Figure
1(source: Malaurie et al., 1998b) shows that D. bulbifera
still has high survival with high sucrose concentration and after long duration
dehydration (up to 23h). For the two species, the water content of encapsulated
apices had to be decreased down to 0.15g H2O g-1 DW in order to obtain high
survival after freezing. The percentage of water loss was of 67, 62, 58 and
55% FW (± 1%) for 0.75, 0.9, 1 and 1.1M sucrose pretreatments, respectively.
Our results demonstrated that, in most cases, survival increased when dehydration
was extended to a defined threshold, around 0.13-0.15g. H2O g-1 DW, which was
obtained after desiccation periods from 10 to 18h. It seemed that, with this
soft dessication process, we could rub out differences in residual water-free,
which certainly exist between apices from a same plot.
Indexation and
disease-free germplasm production
Indexation
In vitro germplasm conservation presents
different advantages such as: 1) to be free from genetic erosion, 2) to have
the possibility for the establishment of core collection with long term genebanks,
3) to be free from fungis and bacteria, 4) to be not expensive, when in vitro
facilities are already present, 5) easy and convenient for international distribution.
But International exchanges need more for safe international exchange. We need
to know the plant material on genetic level, and over all on the phytosanitary
level. On the phytosanitary level, various viruses have been described on edible
and medicinal yams on their production area. Different works, depending virus
and virus group, are reported (Table 7). Indexing techniques
allow to highlight a certain number of viruses on yam: Poty, potex, badna and
cucumo-viruses, where yam mosaic virus (YMV) provokes the most important loss.
Table 7. Viruses
of yam: group and type viruses, yam species affected and reference works
|
Virus Group
|
Virus
|
Yam species affected
|
Geographic spreading
|
Disease importance
|
Authors
|
|
Cucumovirus
|
CMV
|
D. alata,
D. cayenensis-D. rotundata
complex, D. trifida
|
Caribbean and West-Africa
|
-
|
Migliori, 1977;
Fauquet and Thouvenel, 1987
|
| |
|
|
|
|
|
|
‘Carlavirus’ cf.
|
ChYNMV
(Chinese yam necrotic mosaic virus)
|
D. batatas
|
Japan
|
-
|
Fukomoto and Tochihara,
1978; Shirako and Ehara, 1986
|
| |
|
|
|
|
|
|
Badnavirus
|
DBV
(Dioscorea bacilliform
badnavirus)
|
D. alata
|
Barbade
|
+/-
|
Mantell and Haque,
1978
|
| |
DaBV
(D. alata bacilliform virus)
|
|
|
-
|
Degras, 1986
|
| |
DbBV
(D. bulbifera bacilliform
virus)
|
|
|
-
|
Degras, 1986
|
| |
|
|
|
|
|
|
Potexvirus
|
DLV
(Dioscorea latent virus)
|
D. floribunda
D. composita
|
Puerto Rico
|
-
|
Hearon et al., 1978;
Phillips and Brunt 1988; Watterworth
et al., 1974
|
| |
PVX
(Potato virus X)
|
|
In vitro collection
|
-
|
Urbino et al.,1998
|
| |
|
|
|
|
|
|
Potyvirus
|
YMV
(yam mosaic virus)
|
D. alata, D. cayenensis-D.
rotundata complex,
|
All the intertropical area
|
+++
|
Thouvenel and Fauquet,
1979;Goudou-Urbino,1995;Goudou-Urbino
et al. 1996 a,b)
|
| |
YMMV 1
(yam mild mosaic virus)
|
D. alata, D. cayenensis-D.
rotundata complex,
|
West-Africa
|
+++
|
Mumford and Seal,
1997
|
| |
D. trifida virus 2
|
D. trifida
|
Guadeloupe
|
+++
|
Migliori, 1977
|
| |
DGBMV 2
(Dioscorea green banding
mosaic virus)
|
|
Togo
|
++
|
Porth and Nienhaus,
1983
|
| |
DaRMV 3
(D. alata ring mottle virus)
|
D. alata
|
Togo
|
++
|
Porth and Nienhaus,
1983
|
| |
|
|
|
|
|
| |
DaV 4
(D. alata virus)
|
D. alata
D. rotundata
|
Togo
|
+
|
Reckhaus and Nienhaus,
1981
|
| |
D. dumetorum potyvirus
|
|
South-Pacific
|
-
|
Mumford and Seal,
1997
|
| |
D. esculenta potyvirus
|
|
South-Pacific
|
-
|
Mumford and Seal,
1997
|
| |
DGBV (Dioscorea greenbanding
potyvirus)
|
D. composita
D. floribunda
|
Puerto Rico
|
-
|
Hearon et al., 1978;
Phillips et al.,
1986
|
| |
PVY
(Potato virus Y)
|
|
In vitro collection
|
|
Urbino et al., 1998
|
1 YMMV: is it a new potyvirus
or a strain of the YMV ?
2 D. trifida virus and DGBMV have been shown as YMV strains
(Porth et al.,1987)
3 DaRMV should be a ‘yam strain’ of the beet mosaic potyvirus transmissible
on N. benthamiana (Porth et al.,1987)
4 DaV is serologically links toYMV but differ by is non-transmissibility
(Porth et al.,1987)
During the establishment of the yam in
vitro germplasm collection, in the biotechnology laboratory of ORSTOM**
(Malaurie et al., 1993), afterwards IIRSDA - Adiopodoumé
research station, near Abidjan, Ivory Coast - clones were systematically indexed
by ELISA directed to YMV, when introduced in vitro (Malaurie
and Thouvenel, 1988; Malaurie et al., 1988a,b;
Charrier and Hamon, 1991).
Later on, one indexation was carried out
by a virologist team on the duplicate of the yam in vitro germplasm collection
enriched by introduction of new genotypes. 92 samples, belonging to several
yam species, were used for the indexation : D. alata, D. bulbifera, D. cayenensis-
rotundata complex, D. dumetorum, D. esculenta, D. mangenotiana, D. praehensilis,
D. shimperiana, D. togoensis, D. trifida. These samples were originating
from various geographic areas: Africa, Caribbean, South America and Asia. Four
viruses were fetched by ELISA technique: PVX (potato virus X, potexvirus) PVY
(potato virus Y, potyvirus), CMV (cucumber mosaic cucumovirus), and YMV (Urbino
et al. 1998). Results are presented in Table 8.
Table 8. Indexation
of the ORSTOM** in vitro yam collection
|
ELISA results
|
PVX
|
PVY
|
CMV
|
YMV
|
|
% Positives
|
5,5
|
6,3
|
2,1
|
7,7
|
|
% Negatives
|
94,5
|
93,7
|
97,9
|
92,3
|
Source: Urbino et al. (1998)
This study allowed to show
that detection of viruses serologically linked to PVX and to PVY, in different
yam species, was possible, even with the same frequencies than with YMV. Further
works have to be done for a precise caracterization of these virus isolates,
and check their respective importance in natural environement. Experiments using
more sophisticated techniques for virus diagnostic (IC/rt/PCR) are developed
(Bousalem, 1995). Yam on molecular caracterization and molecular
diversity on potyvirus of the yam mosaic virus (YMV) have been developed on
the ILTAB/ORSTOM**-TSRI laboratories (Aleman, 1996; Aleman
et al 1996a,b) and from the LPRC laboratory (Bousalem,
1995; Urbino et al., 1998).
Virus eradication
techniques
The use of in vitro techniques allows
to be free from fungis, bacteria, and other pests. Only viruses could be present
on the plant and have to be eradicated. Different techniques exist and are already
applied on yam. There are meristem culture, thermotherapy and/ or chemotherapy
(36°C during 1 to 2 weeks on in vivo or in vitro plants, use of
chemicals such as vidarabine, ribavirin and 2-thiouracil). They could be used
alone or associated (Table 9).
Table 9 . Yam
disease eradication techniques
| |
Eradication techniques
|
Species
|
Authors
|
Type of use*
|
Virus eradication
|
|
(A)
|
meristem culture
|
D. cayenensis-rotundata,
D. japonica, D. opposita,
D. praehensilis,
D. rotundata, D. trifida, Dioscorea
spp.
|
Cortes Monllor et al.,
1982; Kobayashi 1991; Malaurie et
al 1988a,b, 1992, 1995a,b;
Malaurie and Thouvenel, 1988;
Matsubaru and Ishira, 1988; Mikami ,1984; Saleil
et al., 1990;
|
E
|
+ / -
|
| |
|
|
|
|
|
|
(B)
|
Thermotherapy in vivo + meristems
culture
|
D. alata
|
Mantell et al 1980
|
E
|
+
|
|
(C)
|
Nodal microcutting or apices + Thermotherapy
|
D. alata, D. trifida
|
Balagne 1985; Salazar
and Fernanadez, 1988
|
E, R (+ / -)
|
+
|
| |
|
|
|
|
|
|
(D)
|
Nodal microcutting + Chemotherapy
|
D. alata
|
Mantell ,1993
|
E, R (+ / -)
|
+
|
|
(E)
|
Nodal microcutting + Thermotherapy
&/or Chemotherapy
|
D. praehensilis
|
Malaurie, unpublished results
|
E
|
+ / -
|
|
(F)
|
Meristem culture + Thermotherapy
&/or Chemotherapy
|
D. cayenensis-rotundata,
D. praehensilis
|
Malaurie, unpublished results
|
E
|
+ / -
|
* E: experimental use; R: routine use
Success in meristem culture depends on the
size and location of the explant excised, and on the growth regulator ratio.
‘Meristem culture’, on Table 9, concerns works using meristem-tips
(0.2-0.5 mm long) as well as shoot-tips (0.6-2.5 mm long). Experiments on viability
and in vitro morphological development of meristem-tips of two sizes,
‘small’ (0.3-0.5 mm) and ‘large’ (0.6-0.8 mm), have shown that it was better
to use large meristem size to increase the shoot elongation percentage. The
use of axillary or apical meristems did not induce difference and should allow
an important yield in micropropagating such material from excised meristem-tips.
Eleven months after meristem excision, production of plantlets was observed
with a rate of 82% and 39% from the survivors, for a clone of D. cayenensis-D.
rotundata complex and D. praehensilis genotype, respectively (Malaurie
et al., 1995a,b).
Meristem cultures have been done on 8 clones
of 5 Dioscorea species belonging to the in vitro germplasm collection.
Morphological development has been observed and data were recorded 60 days after
meristem inoculation. In our case, the production of rooted leafy shoots, 60
days after meristem inoculation, occurred in five clones out of eight, with
percentage shoot leaf production of 5 to 26 %, depending on the clone. Six months
later, the excised meristems of all clones developed into rooted leafy shoots,
where D. bulbifera, and D. dumetorum was not, to our knowledge,
mentioned in the literature (Table 10).
Table 10. Genotypic
effect on morphogenetic orientation
2 months after meristem excision of Dioscorea
spp *
| |
Total meristems observed
|
Necrosis 1)
%
|
|
Organogenesis 2)
%
|
|
Regeneration 3)
%
|
|
|
D. alata
|
67
|
27
|
|
55
|
|
18
|
|
|
D. bulbifera
|
252
|
46
|
|
37
|
|
18
|
|
|
D. bulbifera
|
146
|
52
|
|
48
|
|
0
|
|
|
D. cayenensis-D. rotundata complex
|
23
|
65
|
|
35
|
|
0
|
|
|
D. cayenensis-D. rotundata complex
|
81
|
24
|
|
51
|
|
26
|
|
|
D. cayenensis-D. rotundata complex
|
81
|
26
|
|
54
|
|
20
|
|
|
D. dumetorum
|
24
|
83
|
|
17
|
|
0
|
|
|
D. praehensilis
|
117
|
41
|
|
54
|
|
5
|
|
1) Necrosis. 2) Organogenesis: callusing,
rooting, swelling were added together, 3) Regeneration: meristem development
into rooted leafy shoots and axillary bud development or bud neoformation.
*(Malaurie, unpublished results)
Works about production of virus-free in
vitro plants of yam through yam meristem culture alone are very rare. Saleil
et al. (1990) on D. trifida obtained YMV-free plants, after ELISA
indexation with a 27% rate through the total indexed plants. Nevertheless, other
unpublished results on 2 genotypes YMV-infected of 2 species (D. cayenensis-
D. rotundata complex, D. praehensilis) showed that meristem culture
allowed the production of virus-free plants with 76% and 17% plants indexed,
respectively (Malaurie, unpublished results).
Production of virus-free in vitro
plants of yam has been attempted through thermotherapy, chemotherapy associated
or not, from in vivo mother plants, nodal cuttings or apices (Balagne,
1985; Mantell, 1993; Mantell et al., 1980;
Salazar and Fernandez, 1988). None of them described clearly
the percentage of virus-free plants obtained through these techniques. Meanwhile,
the production of plantlets free from virus is described by Mantell
(1993) on D. alata cv. Kinabayo, after the action of antiviral agents
(vidarabin, ribavirin) on nodal microcuttings infected by a potyvirus. The production
of virus-free plants have been obtained 210 days after in vitro inoculation,
after 3 subcultures of 60, 120 and 30 days on a liquid/solid biphasic cuture
system with 10-5 M of antiviral agent.
Other available techniques could be electrotherapy
used on potato, with 60 to 100% success as compared to 25-40% with thermotherapy
(Lozoya-Saldaña et al., 1996; Bernal et
al., 1998), or apex micrografting, used on Lemon tree or vine, routinely.
If different works have already been done
on yam sanitation, only a few of them conducted to an eradication of virus with
more or less importance.
Safe international
exchange
Exchange and distribution of plant material
could be done by two ways: 1) with non aseptic plant material (tubers, aerial
tubers, seeds, nodal cuttings from the vine), 2) with plant material in aseptic
conditions (micro-nodal cuttings, microtubers, aerial microtubers, apices, zygotic
or somatic embryos, callus and cells suspension).
Exchange in non-aseptic conditions was used
in the past, but required severe quarantine measures. Since 1989, with the FAO/IBPGR
technical guidelines for the safe movement of yam germplasm, recommendation
has been given to use in vitro conditions for exchange and distribution.
For that, safe movement of yam germplasm could be done easily by three ways:
1) micro-nodal cuttings, 2) micro-tubers, 3) or encapsulated apices.
Safe movement of yam germplasm by micro-nodal
cuttings is the most common way and has been frequently used (Malaurie
et al., 1998a). In Table 11, the use of laboratories
with in vitro and quarantine facilities allowed the indexation, in
vitro introduction and micropropagation for a safe diffusion of various
genotypes from different geographical origin.
Table 11. Enrichment of the genetic diversity
of a country by transfer and introduction
of in vitro yam genotypes from
different geographic origins*
|
Species
|
Number of accession
|
Sending countries
|
Receiving countries
|
|
D. alata
|
6
|
Côte d’Ivoire
|
Nouvelle Calédonie
|
|
D. alata
|
5
|
West Indies
|
Nouvelle Calédonie
|
|
D. alata
|
1
|
Brazil
|
Nouvelle Calédonie
|
|
D. alata
|
5
|
West Indies
|
Côte d’Ivoire
|
|
D. alata
|
3
|
Nouvelle Calédonie
|
Côte d’Ivoire
|
|
D. alata
|
3
|
Brazil
|
Côte d’Ivoire
|
|
D. bulbifera
|
1
|
Nouvelle Calédonie
|
Côte d’Ivoire
|
|
D. cayenensis-D. rotundata complex
|
4
|
Côte d’Ivoire
|
Nouvelle Calédonie
|
|
D. cayenensis-D. rotundata complex
|
1
|
Brazil
|
Côte d’Ivoire
|
* All plant material from the sending countries
were, at first, tubers sent to laboratories with quarantine and in vitro
culture facilities (1988-89: Orstom** & Iirsda, Adiopodoumé, Côte
d’Ivoire; 1992-95: Orstom**, LRGAPT, Montpellier) for their in vitro
introduction and micropropagation, preliminary to all safe international exchange.
Tuber potentiality shown by a great number
of in vitro yams (aerial and basal micro-tubers) could be also used for
a safe transfer of yam germplasm. They could increase the percentage of success
during their acclimatation in field (John et al.,1993; Malaurie
et al., 1993; Mantell, 1993; Ng, 1988;
Ng and Mantell, 1997). These tubers developed in vitro
are dormant at maturity and they still keep their dormancy from 2 to 5 months,
as tubers developed in vivo.
Recently a new method, experimented over
three yam species (D. alata, D. opposita, D. rotundata),
has been proposed by Hasan and Takagi (1995). They use encapsulation
technique, with the embeddment of nodal cuttings in alginate beads, for a concept
of a material transfer. This process allow to maintain in the dark for at least
2 weeks. These 2 weeks in the dark allow to envisage a safe and easy international
exchange of genetic resources.
Concluding remarks
This paper tries to describe different studies
done and to be done on yam in vitro germplasm conservation and its safe
international exchange. Yam in vitro culture contributes to the safeguard
of the biodiversity of the genus Dioscorea. An application of the results
obtained on cryopreservation to more species should allow a transfer of technology.
The use of new techniques, in a one hand, for pathogen eradication (electrotherapy,
micrografting), in addition to the existent ones, and in the other hand, for
the obtention of plants resistant to somes viruses (transformation), should
guarantee to yam a state of virus-free plant and allow international exchanges,
and in long term, distribution to the farmer of cultivar free from virus.
To conclude we can say that we are already
able to manage routinely yam in vitro genebanks in slow growth culture,
to index for more viruses, and to produce some virus-free in vitro plantlets.
For an efficient distribution - transfer
- utilisation of yam germplasm, we should develop: 1) virus-free germplasm,
2) restricted size collection, with large diversity, so-called core-collections.
For that, in vitro conservation under slow growth condition and cryopreservation,
have to be applied routinely to more genotypes; virus-indexing has to be done
with more precise techniques (rt/PCR); therapy has to be done with several combined
techniques to become genotype independant.
But, we should not forget, as previously
said by Hanson (1986), that, for a better security of germplasm
conservation, different methods of conservation have to be combined (in situ
- Field Genebanks - , ex situ - Seed Genebanks, in vitro Genebanks).
References
- Aleman, M.E. (1996). Caractérisation
moléculaire, diversité génétique et contrôle
du virus de la mosaïque de l'igname (YMV). These, Doctorat en sciences
agronomiques. ENSAM, Montpellier, France, 138p.
- Aleman, M.E., Goudou-Urbino, C., Dubern,
J. and Fauquet, C. (1996a). Analysis of sequence variations in the P1, HC,
P3, NIb and CP regions of Yam Mosaic Potyvirus isolates: implications for
potyvirus intra-species molecular diversity. Journal of General Virology 78:
1253-1264.
- Aleman, M.E., Marcos, J.F., Brugidou,
C., Fux, C.I., Goudou-Urbino, C. and Dubern, J. (1996b). Genomic Organization,
Molecular Diversity and Control of Yam (Dioscorea) Mosaic Virus. ILTAB
Review 1991-1996, p. 81-82.
- Ayensu, E.S. and Coursey, D.G. (1972).
Guinea yams. The botany, ethnobotany, use and possible future of yams in West
Africa. Economic Botany 26: 301-318.
- Balagne, M. (1985). Le microbouturage
in vitro de l'igname cousse couche Dioscorea trifida en vue
de son application pour la guérison des variétés atteintes
de viroses. Thèse de 3ème cycle, USTL, Montpellier, 144 p.
- Bernal, F.J.M., Hernández, R.P.,
Noa, J.C., Pichardo, T.P., Igarza, Y.C., Sarria, Z. and Agromonte, D.P. (1998).
Técnicas de saneamiento para la obtención de papa (Solanum
tuberosum L.) var. Desirée libre del virus del enrollamiento de
la hoja. REDBIO’98-3rd Latin-American Meeting on Plant Biotechnology,
June 1-5, La Habana, Cuba, p. 137.
- Bousalem, M. (1995). Mise au point d'outils
immunologiques et moléculaires pour la détection et la caractérisation
des potyvirus infectant l'igname- Caractérisation moléculaire
préliminaire de l'isolat BFC 56 du Yam Mosaic Virus. Laboratoire de
Phytovirologie des Régions Chaudes (LPRC), Montpellier, Cirad-Orstom**,
23p. (document interne).
- Burkill, I.H. (1960). The organography
and the evolution of Dioscoreaceae, the family of the yams. J. Linn. Soc.
(Bot.) 56: 319-412.
- Butenko, R.G., Popov, A.S., Volkova,
L.A., Chernyak, N.D. and Nosov, A.M. (1984). Recovery of cell cultures and
their biosynthetic capacity after storage of Dioscorea deltoïdea
and Panax ginseng cells in liquid nitrogen. Plant Science Letters 33:
285-292.
- Charrier, A., Dereuddre, J., Engelmann,
F. (1991). The implications of biotechnology in germplasm conservation and
utilization. Crop Genetic Resources of Africa. Vol II. N.Q. Ng, P. Perrino,
F. Attere, H. Zedan ed., Ibadan, Nigeria, IITA/IBPGR/UNEP/CNR, In: Proceedings
of an International Conference on Crop Genetic Resources of Africa. pp. 279-286.
- Charrier, A. and Hamon, S. (1991). Germplasm
collection, conservation and utilization activities of the Office de la Recherche
Scientifique et Technique d'Outre-Mer (ORSTOM**). Crop Genetic Resources of
Africa. Vol II. N.Q. Ng, P. Perrino, F. Attere, H. Zedan ed., Ibadan, Nigeria,
IITA/IBPGR/UNEP/CNR. In: Proceedings of an International Conference on Crop
Genetic Resources of Africa. pp. 41-52.
- Chulafich, L., Grubishich, D., Vuiichich,
R., Volkova, L.A. and Popov, A.S. (1994). Somatic embryo production in
vitro in Dioscorea caucasica Lipsky and Dioscorea balcanica
Kosanin and cryopreservation of their organogenic callus tissue. Russian Journal
of Plant Physiology 41: 821-826.
- Cortes Monllor, A., Liu, L.J. and Arroyo,
E. (1982). An improved medium for tissue culture of yam Dioscorea sp.
in vitro in Puerto Rico. Phytopathology 72 (1): 171.
- Coursey, D.G. (1967). Yams, London,
Longmans, 230 p.
- Degras, L. (1986). L'igname, Techniques
Agricoles et Productions Tropicales, Ed. G.P. Maisonneuve et Larose &
Agence de Coopération Culturelle et Technique 409 p.
- Dereuddre, J., Scottez, C., Arnaud,
Y. and Duron, M. (1990). Résistance d'apex caulinaires de vitroplants
de poirier (Pyrus communis L. cv Beurré Hardy), enrobés
dans l'alginate, à une deshydratation puis à une congélation
dans l'azote liquide: effet d'un endurcissement préalable au froid.
C. R. Acad. Sc. Paris, Série III, 310: 317-323.
- Dereuddre, J., Blandin, S. and Hassen,
N. (1991). Resistance of alginate-coated somatic embryos of carrot (Daucus
carota L.) to dessication and freezing in liquid nitrogen: 1- Effects
of preculture. Cryo-Letters 12:1125-134.
- Engelmann, F. (1991). In vitro
conservation of tropical plant germplasm - a review. Euphytica 57: 227-243.
- Fauquet, C. and Thouvenel, J.C. (1987).
Cucumber mosaic on sweet potato and yam. In: Plant Viral Diseases in
the Ivory Coast, Documentations Techniques n°. 46, Editions de l'Orstom**,
Paris, p. 29.
- FAO/IBPGR (1989). Technical guidelines
for the safe movement of yam germplasm. A.A. Brunt, G.V.H. Jackson, E.A. Frison,
ed., Rome, pp. 1-20.
- FAO (1996). The State of the World’s
Plant Genetic Resources for Food and Agriculture. Background Documenation
prepared for the International Technical Conference on Plant Genetic Resources,
Leipzig, Germany, 17-23 June, 1996. FAO : Rome, 336 p.
- Fukomoto, F. and Tochihara, H. (1978).
Chinese yam necrotic virus. Annals of the Phytopathological Society of Japan
44: 1-5.
- Goudou-Urbino, C. (1995). La mosaïque
de l'igname: aspects épidémiologiques au Burkina-Faso et variabilité
du virus. Thèse de l'Université Montpellier II, 147 p.
- Goudou-Urbino, C., Givord, L., Quiot,
J.B., Boeglin, M., Konate, G. and Dubern, J. (1996a). Differentiation of yam
virus using symptomatoloy, western blot assay, and monoclonal antibodies.
Journal of Phytopathology 144 : 235-240.
- Goudou-Urbino, C., Konate, G., Quiot,
J.B. and Dubern, J. (1996b). Etiology and ecology of yam mosaic disease in
Burkina Faso. Tropical Sciences 36: 34-40.
- Hanson, J. (1986). Methods of storing
tropical root crop germplasm with special reference to yam. FAO/IBPGR Plant
Genetic Ressources Newsletter 64: 24-32.
- Hasan, S.M.Z. and Takagi, H. (1995).
Alginate-coated nodal segments of yam (Dioscorea spp.) for germplasm
exchange and distribution. Plant Genetic Resources Newsletter 103: 32-35.
- Hearon, S.S., Corbett, M.K., Lawson,
R.H., Gillespie, A.G. and Waterworth H.E. (1978). Two flexuous-rod viruses
in Dioscorea floribunda: symptoms, identification and ultrastructure.
Phytopathology, 68: 1137-1146.
- IBPGR (1986). 2. Root and Tuber Crops
(2nd Ed.), In Directory of Germplasm Collections. J. Toll, T. Lawrence,
D.H. Van Sloten Ed. International Board for Plant Genetic Resources, Rome,
pp. 130-155.
- John, J.L., Courtney, W.H. and Decoteau,
D.R. (1993). The influence of plant growth regulators and light on microtuber
induction and formation in Dioscorea alata L. cultures Plant Cell,
Tissue and Organ Culture 34: 245-252.
- Knuth, R. (1924). Dioscoreaceae. A.
Engler. Das Pflanzenreich, IV-43, 87. Heft 1-387.
- Kobayashi, N. (1991). Production of
virus-free plants and its mass propagation by tissue culture in Chinese yam,
Dioscorea opposita Thunb. Bulletin of the Saitama Horticultural Experiment
Station. N° 18, 81-99.
- Latta, R. (1971). Preservation of suspension
culture of plant cells by freezing. Canadian Journal of Botany 40: 1253-1254.
- Lozoya-Saldana, H, Abello, F.J. and
Garcia G de la R. (1996). Electrotherapy and shoot tip culture eliminate potato
virus X in potatoes. American Potato Journal 73: 149-154.
- Malaurie, B., Tardieu, F. and Thouvenel,
J.C. (1988a). Rapid production of disease-free germplasm of Dioscorea
spp. (Monocotyledonous). In: Proceedings of 8th International Biotechnology
Symposium, Paris, 17-22 July 1988, p. 248.
- Malaurie, B., Tardieu, F. and Thouvenel,
J.C. (1988b). Improving Yam (Dioscorea spp.), using Biotechnology.
1) Rapid production of disease free germplasm. In: Proceedings of the International
Society for Root Crops Symposium. Bangkok, 30/10-5/11/1988, p. 51.
- Malaurie, B. and Thouvenel, J.C. (1988).
Production de plants d'Igname, dépourvus de viroses, par culture de
méristèmes. 2° Salon International de la Coopération
et de l'Aide au Développement (SICAD). 7-11/12/1988, Montpellier, Poster
Session, 3 p.
- Malaurie, B., Pungu, O., Dubern, J.
and Thouvenel, J.C. (1992). Determination of the best conditions for the regeneration
of microplants and the elimination of YMV from excised meristems of yam nodal
cuttings (Dioscorea spp.). Association of Applied Biologists. Plant
Virology in the Tropics. University of York. 9-10 April 1992.
- Malaurie, B., Pungu, O., Dumont, R.
and Trouslot, M.F. (1993). The creation of an in vitro germplasm collection
of yam (Dioscorea spp.) for genetic resources preservation. Euphytica
65: 113-122. Corrigendum 66: 243
- Malaurie, B., Pungu, O. and Trouslot,
M.F. (1995a). Effect of growth regulators concentrations on morphological
development of meristem-tips in Dioscorea cayenensis-D. rotundata complex
and D. praehensilis. Plant Cell, Tissue and Organ Culture 41: 229-235.
- Malaurie, B.,Thouvenel, J.C. and Pungu,
O. (1995b). Influence of meristem-tip size and location on morphological development
in Dioscorea cayenensis-D. rotundata complex 'Grosse Caille' and one
genotype of D. praehensilis. Plant Cell, Tissue and Organ Culture 42:
215-218.
- Malaurie, B. and Trouslot, M.F. (1995c).
Les Ignames. In Biotechnologies végétales BV9D, CNED-AUPELF-UREF
ed., chapitre 10: 49-77.
- Malaurie, B. and Trouslot, M.F. (1996).
Cryopreservation of in vitro yam (Dioscorea sp.) apices by the
encapsulation-dehydration technique. Eucarpia, Meeting on Tropical Plants,
Marsh 11-15 1996, France, Abstract, p 252.
- Malaurie, B., Trouslot, M.F. and Berthaud,
J. (1998a). Conservation et échange de germoplasme chez les ignames
(Dioscorea spp.).‘L’igname : Plante Séculaire et Culture
d’Avenir’. eds. J. Berthaud, N. Bricas, J.-L. Marchand, Montpellier 3-6 Juin
1997, In : Acte du séminaire international Cirad-Inra-Orstom**-Coraf,
France. 135-161.
- Malaurie, B., Trouslot, M.F., Engelmann,
F. and Chabrillange, N. (1998b). Effect of pretreatment conditions on the
cryopreservation of in vitro-cultured yam (Dioscorea alata
'Brazo Fuerte' and D. bulbifera 'Nouméa Imboro') shoot apices
by encapsulation-dehydration. Cryo-Letters 19 : 15-26.
- Malaurie, B., Trouslot, M.F., Berthaud,
J., Chabrillange, N., Récalt, C. and Dussert, S. (1998c). The use of
slow growth condition culture and cryopreservation in liquid nitrogen for
medium and long term conservation and utilisation of in vitro yam (Dioscorea
spp.) germplasm. In: Proceedings of the Workshop on Conservation & Utilisation
of Cassava, Sweetpotato and Yam Germplasm in Sub-Saharan Africa. Nairobi,
November 11 to 13, 1997, (in press).
- Mandal, B.B., Chandel, K.P.S. and Dwivedi,
S. (1996). Cryopreservation of yam (Dioscorea spp.) shoot apices by
encapsulation-dehydration. Cryo-Letters 17: 165-174.
- Mantell, S.H. and Haque, S.Q. (1978).
Incidence of internal brown spot disease in White lisbon (D. alata)
during storage. Experimental Agriculture 14: 167-172.
- Mantell, S. H., Haque, S. Q. and Whitehall,
A. P. (1980). Apical meristem tip culture for eradication of flexous rod viruses
in yams (Dioscorea alata). Tropical Pest Management 26 : 170-179.
- Mantell, S.H. (1993). Integrated use
of micropropagation and conventional propagation techniques for production
of certified seed tubers of tropical yams (Dioscorea spp.). Adapted
propagation techniques for commercial crops of the tropics, IFS, Vietnam,
02/1993. In: Proceedings of the Southeast Asian Regional Workshop on Propagation
Techniques for Commercial Crops of the Tropics. 66-93.
- Martin, F.W. and Degras, L. (1978).
Tropical yams and their potential. Part 6. Minor cultivated Dioscorea species.
USDA agriculture handbook n° 538. 23 p.
- Martin, F.W. and Rhodes, A.M. (1978).
The relationship of Dioscorea cayenensis and D. rotundata. Tropical
Agriculture, Trin., 55: 193-206.
- Matsubara, S. and Ishihara, M. (1988).
Production and vegetative propagation of virus-free plants of Dioscorea
species. Scientific Reports of the Faculty of Agriculture, Okayama University,
N° 72, 19-26
- Miège, J. (1982). Etude chimiotaxonomique
de dix cultivars de Côte d’Ivoire relevant du complexe Dioscorea
cayenensis-D. rotundata. Yams-Ignames, J. Miège, S.N. Lyonga, Ed.,
Claredon Press Oxford, pp. 197-231.
- Migliori, A. (1977). Maladies à
virus de l’igname Dioscorea spp. Nouvelles agronomiques Antilles Guyane,
3: 428-435.
- Mikami, T. (1984). Virus-free plant
propagation through meristem-tip culture of Japanese yam and garlic. Research
Journal of Food and Agriculture 7 : 17-20
- Mumford, R.A. and Seal, S.E. (1997).
Detection and characterization of yam potyviruses. In : Yam viruses,
Annual Collaborators Meeting, IITA - JIC - NRI, Gatsby-funded Biotechnology
Projects, 7-9 February 1997, IITA, Ibadan, Nigeria, Extended Abstracts, pp.
14-15.
- Ng, S.Y.C. (1988). In vitro tuberization
in white yam (Dioscorea rotundata Poir.). Plant Cell, Tissue and Organ
Culture 14: 121-128.
- Ng, S.Y.C. and Mantell, S.H. (1997).
Technologies for Germplasm Conservation and Distribution of pathogen-free
Dioscorea yams to National Root Crop Research Programmes. ODA Project
R4886 (H) Final Report. Wye College University of London, UK, 12p.
- Phillips, S. and Brunt, A.A. (1988).
Dioscorea latent virus. AAB Descriptions of Plant Viruses, N°. 335.
Association of Applied Biologists, Wellesbourne.
- Phillips, S., Pigott, J. d’A. and Brunt,
A.A. (1986). Further evidence that Dioscorea latent virus is a potexvirus.
Annals of Applied Biology 109: 137-145.
- Polge, C.A., Smith, A.V. and Parkes,
A.S. (1949). Revival of spermatozoa after vitrification and dehydratation
at low temperature. Nature 164: 666
- Popov, A.S. and Volkova, L.A. (1994).
Cryopreservation and some characteristics of Dioscorea deltoïdea
cell cultures in the vitamin-free medium. Russian Journal of Plant Physiology
41 : 815-820.
- Popov, A.S. and Fedorovskii, D.N. (1992).
Injuries to the plasmalemma of Dioscorea cells cultured in vitro
incurred in the process of their cryopreservation. Soviet Plant Physiology
39 : 211-216.
- Porth, A., Lesemann, D.-E. and Vetten,
H.J. (1987). Characterization of potyvirus isolates from West Africa yams
(Dioscorea spp.). Journal of Phytopathology 120: 166-183.
- Porth, A. and Nienhaus, F. (1983). Dioscorea
alata ring mottle virus, a new potyvirus in yam in Togo. Zeitschrift fur
Pflanzenkrankheiten und Pflanzenschutz 90: 352-362.
- Reckhaus, P. and Nienhaus, F. (1981).
Etiology of a virus disease of white yam D. rotundata in Togo. Journal
of plant disease and protection 88 : 492-509.
- Sakai, A. (1984). Cryopreservation of
apical meristems. In Horticultural Reviews, J. JANICK Ed., Purdue University,
pp. 357-372.
- Salazar, S.S and Fernandez, R.Z.Z. (1988).
Thermotherapy, shoot tip culture, axillary bud proliferation and plant regeneration
in yam (Dioscorea trifida L.). In VIIth Symposium of the International
Society for Tropical Root Crops, Gosier (Guadeloupe), 1-6 July 1985, INRA
Ed., Paris, p. 439-445.
- Saleil, V., Degras, L. and Jonard, R.
(1990). Obtention de plantes indemnes du virus de la mosaïque de l'igname
(YMV) par culture in vitro des apex chez l'igname americaine Dioscorea
trifida L. Agronomie, 10: 605-615.
- Shirako, Y. and Ehara, Y. (1986). Rapid
diagnosis of chinese yam necrotic mosaic virus infection by electro-blot immunoassay.
Annals of the Phytopathological Society of Japan 52: 453-459.
- Tanaka, S. (1977). Chinese yam Dioscorea
opposita Thunb. in Japan. Tropical Root and Tuber Crops Newsletter n°10,
pp. 4-5.
- Tannoury, M., Ralambossa, J., Kaminske,
M. and Dereuddre, J. (1991). Cryoconservation par vitrification d'apex enrobés
d'oeillets (Dianthus caryophyllus L.) cultivés in vitro.
C. R. Acad. Sc. Paris, Série III, 313: 633-638.
- Thouvenel, J.-C. and Fauquet, C. (1979).
Yam mosaïc, a new potyvirus infecting Dioscorea cayenensis in
the Ivory Coast. Annals of Applied Biology 93: 279-283.
- Uragami, A. (1993). Cryopreservation
of cultured cells and organs of vegetables. In Reprint from JICA GRP REF N°6,
111-135.
- Urbino, C., Bousalem, M., Pinel, A.,
Fargette, D. and Dubern, J. (1998). Les Virus de l’igname : aspects épidémiologiques
et variabilité.‘L’igname : Plante Séculaire et Culture
d’Avenir’. eds. J. Berthaud, N. Bricas, J.-L. Marchand, Montpellier 3-6 Juin
1997, In : Acte du séminaire international Cirad-Inra-Orstom**-Coraf,
France. 205-211.
- Watterworth, H.E., Lawson, R.H. and
Kahn, R.P. (1974). Purification, electron microscopy, and serology of Dioscorea
latent virus. Journal of Agriculture of the University of Puerto Rico 58:
351-357.
- Withers, L.A. (1991). Crop strategies
for roots and tubers: potato- A model for refinement, Yam- a problem for development.
In Workshop on Conservation of Plant Genetic Resources. Becker B. Ed.,
Bonn. ATSAF/IBPGR, pp. 11-14.
Supported by UNESCO
/ MIRCEN
network
© 1998 by Universidad Católica de Valparaíso
-- Chile
The following images related to this document are available:
Photo images
[ej98015f1.jpg]
|

{kind=link}