 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 1, No. 3, December, 1998 In vitro fruit trees rooting by Agrobacterium rhizogenes wild type infection Carmine Damiano*1 and Simona Monticelli2 1Fruit
Trees Research Institute, 00040 Ciampino Aeroporto - Roma, Italia. Tel: + 39 06
7934811- Fax +39 06 79340158 E-mail: isf.propag@mclink.it Financial Support: National Plan for Plant Biotechnology, Ministry of Agriculture Received
24 July 1998 Code Number: ej98022 Agrobacterium rhizogenes infection at the base of microcuttings in vitro can improve the rooting of some fruit species. A study was carried out comparing rooting of almond, apple, plum, Pyrus pyraster and two hybrid rootstocks, when infected with A. rhizogenes strain 1855, with and without the addition of hormones. Three responses occurred: genotypes rooted without auxins; genotypes rooted only with auxins; genotypes rooted only after infection. All genotypes rooted after bacterial infection. In the first group the auxins increased rooting percentages. No substantial differences were found with and without infection in hormone free media, while the rooting percentages tended to decrease with the combination auxin/infection. In the second group, infection on hormone free media increased rooting; in addition there was a synergistic effect between auxins and infection in pears. In the third group only infection induced rooting. A random sample of roots obtained from infection was molecularly analysed. Amplification of the sequences of rolB and vir genes was done using PCR. Roots non-transgenic and confirmed as transgenic were, respectively, 67 % and 6.8 % respectively. In the remaining 26.2% both genes appeared, thus it was not possible to confirm their transgenic nature. Some microcuttings showed both transformed and non-transformed roots. Keywords:
Biotechnology, Propagation, rol gene, Transformation Abbreviations: BAP, 6-benzylaminopurine; IAA, indoleacetic acid; NAA, a naphthaleneacetic acid; IBA, indole-3-butyric acid; GA3, gibberellic acid; PCR, polymerase chain reaction; EtBr, ethidium bromide; YMB, yeast-extract mannitol broth; MS, Murashige and Skoog medium; SH, Schenk and Hildebrandt ; QL, Quoirin and Lepoivre. Many woody plants, economically important for timber and/or fruit production, are often difficult to root, both in conventional and in vitro propagation. In some cases, it was possible to improve in vitro rooting with hormone application, etiolation, or the use of polyamines (Damiano et al., 1991; Rugini et al., 1993). However, the difficulty of rooting is still one of the major obstacles to successful micropropagation. Recently, many attempts to overcome this problem have been carried out on fruit trees and woody species using Agrobacterium rhizogenes. This is a soil gram-negative bacterium that induces adventitious root formation at the site of infection in a large number of plants (Chilton et al., 1982). Root induction is due to the integration and subsequent expression of a portion of bacterial DNA (T-DNA) from the Ri (Root Inducing) plasmid in the plant genome. Four loci involved in the root formation were identified in the T-DNA, and called root loci (rol) A, B, C, D (Spena et al., 1987). Several authors reported successful rooting using Agrobacterium rhizogenes-mediated transformation both in fruit trees, such as almond (Damiano et al., 1995), apple (Sutter and Luza, 1993), kiwi (Rugini et al., 1991), walnut (Caboni et al., 1996), and in woody plants, such as the genus Pinus (McAfee et al., 1993; Magnussen et al., 1994; Mihalievic et al., 1996; Burns and Schwarz, 1996; Tzfira et al., 1996), Larix (McAfee et al., 1993), Eucalyptus (MacRae and Van Staden, 1993). The transformation of microcuttings and seedlings by A. rhizogenes seems to be an excellent way to improve rooting in trees. The aim of this work is to determine the ability of A. rhizogenes to induce root formation on different species and genotypes of micropropagated fruit trees, both recalcitrant and easy-to-root, in the presence or absence of rooting hormones, and to evaluate the nature of these adventitious roots in order to achieve a successful transfer to the field. Materials and MethodsPlant material. In vitro micropropagated plants were used for the experiments on the almond cultivars Fascionello, Ferragnes, Tuono; the almond selections M49, M50, M51, M52, M53, M55; the apple cvs. Gala and McIntosh; three seedlings of Pyrus pyraster (clones P8, P38 and P50); the plum cultivar Ontario, and two hybrid rootstocks, Citation (plum X peach) and GF677 (almond X peach). The microplants were multiplied on Murashige and Skoog medium (1962), except the pear clones for which Quoirin and Lepoivre (1977) was used and Ontario where Schenk and Hildebrandt (1972), supplemented with hormones, as reported in Table 1. Culture conditions were 21±2° C, photoperiod of 16 hrs, with a light intensity 37 m mol m-2 s-1. Preliminary tests were conducted to determine the best root induction medium for each genotype. The auxins concentrations used in the experiments are reported in Table 1. Bacterial inoculum and transformation. Agrobacterium rhizogenes strain 1855 was grown at 28° C on YMB (Hooykaas et al., 1977) for 24-36 hrs until an optical density of 0.4 at 600 nm. was obtained. The infections were carried out according to Damiano et al. (1995), dipping the basal part of the plants for 24 hours in darkness in 0.5 ml bacterial suspension. The microcuttings were prepared after subculturing for three weeks; the most homogeneous shoots were selected and the base cleanly cut. After infection, the explants were transferred to the appropriate co-cultivation root induction medium, with or without hormone, and finally transferred to the same medium containing the antibiotic cefotaxime (250 mg/l). Control uninfected shoots, were dipped in YMB for 24 hrs and then cultured under the same conditions as the infected plants. After infection, microcuttings of Fascionello, Ferragnes, Tuono, M49, M50, M52, M53 M55, P50 were cultured in darkness for ten days before being exposed to light. Some of the explants of Ferragnes and Tuono were maintained with the basal part in darkness after the exposure to light . DNA extraction and PCR. The DNA was extracted from single roots, whose weight ranged from 0.1g to 0.4g (Damiano et al., 1995). The bacteria were removed from the roots by washing in NaOH, 200 mM and SDS 1% w/v, for some minutes and then rinsing in sterile deionised water. The roots, dried on sterile filter paper, were quickly frozen in liquid nitrogen and ground to a fine powder in a mortar containing insoluble 40% w/v PVPP (Damiano et al., 1995). Each PCR reaction was carried out on 20-30ng of total DNA with 1.2U TaqDNA polymerase (Pharmacia) in a volume of 30 µl (Hamill et al., 1991). Primers used for the amplification reactions came from internal sequences of the rolB gene (0.78kb) and from virD1 gene (0.45kb) (Hamill et al., 1991). The amplification temperatures for rolB gene were 95° C for denaturation (duration 1 min.), 34° C for annealing (2 min.) and 72° C for extension (2 min.), with 25 amplification cycles. For virD1 gene amplification denaturation, annealing and extension temperatures were respectively 95° C (1 min.), 55° C (1 min.) and 72° C (2 min.) for 25 cycles. The amplified sequences were made visible on agarose/EtBr gels. Experimental design and data. There were three replicates with 8-10 shoots per plot for each cultivar or selection. Data was recorded after 35 days from the start of the experiment. Data collected was the number of rooted plants, the number of roots per plant and the length of roots in mm. Results, expressed as percentages, were normalised according to the formula arcsineÖ % before statistical analysis. Table 1. Cultural media used
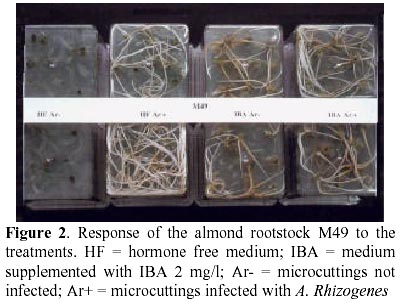
BN, Bourgin and Nitsch (1967); MS, Murashige and Skoog (1962); QL, Quoirin and Lepoivre (1977); SH, Schenk and Hildebrandt (1972); MSmod: MS modified (NH4NO3 660 mg/l, KNO3 1662 mg/l) ResultsTable 2 shows the rooting percentages of the genotypes exposed to the different treatments. According to their rooting ability, the genotypes can be divided into three groups: a) those rooted without auxins: Citation, GF677, M51, Ontario, Gala, McIntosh (Figure 1); b) those rooted only with auxins: M49 (Figure 2), M50, M52, M53, M55, P38, P50 (Figure 3); c) those rooted only after A. rhizogenes infection: P8, Fascionello, Ferragnes, Tuono (Figure 4). In general, the addition of auxins to the media improved rooting in group (a). After bacterial infection all genotypes rooted. However, some differences could be detected. For group (a) no substantial differences were found between hormone free treatments, while a tendency to decrease the percentage rooting was observed with the combination of auxin and infection. With group (b) the positive effect of infection on shoots on hormone free media should be noted and also the synergism between the hormones in the media and infection in the cases of P38 and P50 (Figure 3). No comments are needed on the results of group (c). With Ferragnes and Tuono darkening of the plant bases tended to decrease the rooting, although not significantly (Figure 4). Table 2. Rooting percentages (± S.E.) of microcuttings infected with A. rhizogenes (Ar+) and not infected (Ar-) in hormone free medium (HF) or in the hormone supplemented medium (WH)
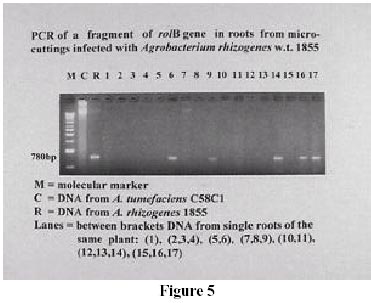
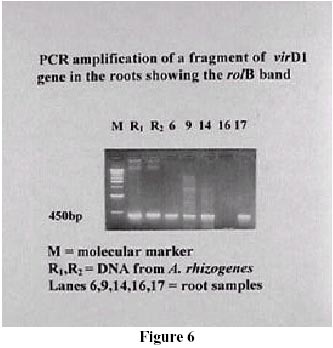
Some changes in the roots number and root length occurred, but more evident was a reduced geotropic response with several infected plants, particularly almond; this growth behaviour was reported as typical in transformed roots (Leguè et al., 1996). Molecular analyses were carried on a random sample of roots of some almond genotypes and GF677. PCR was used to detect the integration of the rolB gene in the roots induced by A. rhizogenes (Figure 5). The virD1 amplification by PCR was done to avoid false positives (Figure 6). In fact, with 26.2 % of the samples it was not possible to exclude the bacterial contamination (Figure 6, lanes 6-9-14-17), thus making the confirmation of the transgenic nature of roots impossible. In the remaining samples, 67 % of the roots were not transformed (Figure 5, lanes 1-2-3-4-5-7-8-10-11-12-13-15) and only the 6.8 % were positively transgenic (Figure 6, lane 16). Some roots with negative geotropism were subjected to PCR analysis in order to correlate the reduced geotropic response with the transgenic nature of the roots. This was not possible, as not all the roots growing upwards appeared to be transgenic. Both transformed and non-transformed roots differentiated on plants cultured in hormone free medium or in the presence of hormones. In some cases the same microcutting showed both transformed and non-transformed roots (Figure 5, lanes 15-16). The results presented in this paper indicate that the use of A. rhizogenes can be a successful approach to improve rooting on fruit trees, although different responses to infection occurred. While with the difficult-to-root genotypes the infection always had a positive effect, sometimes, in association with a hormonal treatment, the responses were larger for easy-to-root ones. From our experiments, it is possible to differentiate two principal behaviours. Most of the species did not appear to be affected by infection and the hormonal supply may be sufficient to give good rooting, as it occurred with the hybrids Citation and GF677, in the apple cultivars Gala and McIntosh and in the almond selection M51. On the other hand, the infection had a detrimental effect on rooting, with or without hormone, on the plum cv. Ontario. The first step in the infection process is the host/pathogen interaction, where several factors contribute to the establishment of this relationship (Winans, 1992). Sugars and phenolic compounds released from the plants, and membrane binding protein can differ according to plant genotypes, thus producing differing responses. By now the hypothesis of a direct auxin release from auxin-glucoside esters by the action of the rolB products has been abandoned (Maurel et al., 1994). Changes in the auxin sensitivity of the rolB in transformed tobacco cells seems to be a consequence of an altered permeability of the membrane auxin binding protein (Filippini et al., 1994; Maurel et al., 1994). Thus, the different responses observed could result from alterations of the auxin reception-transduction pathway. Again, Capone et al. (1994) identified some regulatory domains of the rolB promoter region, responding to endogenous plant regulatory proteins whose presence and/or concentration can differ in different cell types, and maybe in different genetic backgrounds. According to Di Cola et al. (1996), the presence and expression of rolB are necessary but not sufficient to induce rhizogenesis in transformed cells, because a high rooting potential should be associated with the presence of pre-committed cells, whose potentiality could be amplify by rolB. An interesting result emerging from the molecular analysis is that only a few roots were shown to be transgenic. Magnussen et al. (1994) found in infected conifers the induction of normal adventitious roots, but not in untreated seedlings. No correlation could be established between the tranformation rate an the genotipes or to the treatments, because of the size of the sample. The washing step must be changed to eliminate the high percentage of doubtful results. Our results seem to suggest that the rooting process involves more than the transformed cells. McAfee et al. (1993) proposed a role for the bacterium in modifying the root environment by means of organic acid secretions and lowering the pH. Finally, Guivarc'h et al. (1996a, b) suggested the existence of diffuse factors associated with the expression of rolA and rolC, implying that the sites of action and expression of these genes are physically separated. Further investigations are necessary to understand better the rooting and the transformation processes in fruit trees and to improve propagation protocols. Furthermore the field behaviour of plants rooted by localised infection must be investigated. References
Supported by UNESCO / MIRCEN network © 1998 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej98022f4.jpg] [ej98022f2.jpg] [ej98022f1.jpg] [ej98022f3.jpg] [ej98022f6.jpg] [ej98022f5.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}