 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 2, No. 1, April, 1999 REVIEW ARTICLE Feasible biotechnological and bioremediation strategies for serpentine soils and mine spoils Majeti Narasimha Vara Prasad*1 and Helena Maria de Oliveira Freitas2 1Departamento de Botânica,
Faculdade de Ciências e Tecnologia da Universidade de Coimbra, 3000 Coimbra,
Portugal, Permanent address: Department of Plant Sciences, School of Life Sciences
University of Hyderabad, Hyderabad 500046; Fax: +91-040-3010120/3010145 Email:
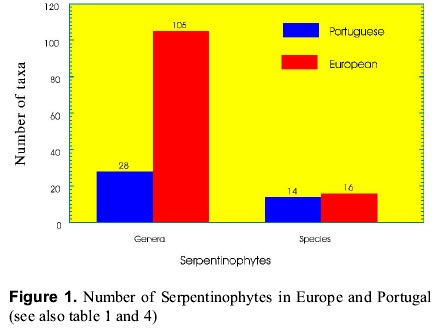
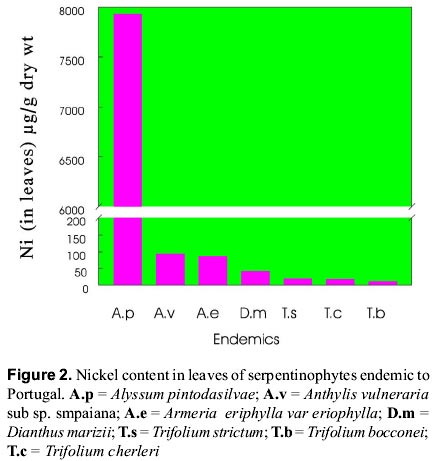
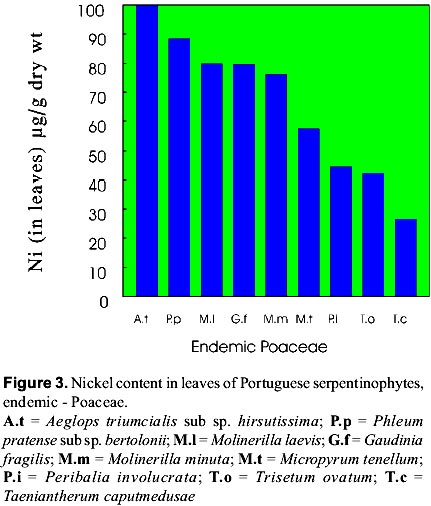
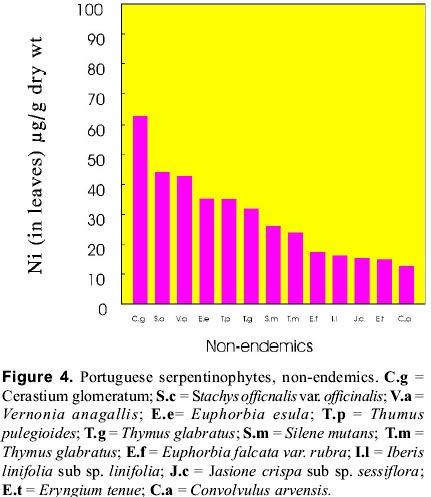
mnvsl@uohyd.ernet.in * MNVP is thankful to the Fundação para a Ciência e Tecnologia, Ministério da Ciencia de Tecnologia, Portugal for awarding visiting fellowship (December 1998 to February 1999). Code Number: ej99004 Reclamation of metalliferous areas is a priority field of biogeochemistry of trace elements. Ultramafic outcrops rich in heavy metals have been mapped in different parts of the world. Heavy metals are potentially cytotoxic, caricinogenic and mutagenic. Environment protection agencies and legislations insisting the mine operators to restore the mine spoils and tailings since the metal leachates have serious implications in production of healthy agricultural products. Hence, restoration of mine spoils, tailings and metalliferous soils is a challenging task for the well being of Humans. Synthetic and natural zeolites have been used as chelators for rapid mobility and uptake of metals from contaminated soils by plants. Use of synthetic chelators significantly increased Pb and Cd uptake and translocation from roots to shoots facilitating phytoextraction of the metals from low grade ores. Contrastingly, synthetic cross linked polyacrylates, hydrogels have protected plant roots from heavy metals toxicity and prevented the entry of metals into roots. However, application of these synthetics on large scale may not be a practical solution due to exorbitant costs. Therefore, introduction of metal tolerant wild plants to metalliferous soils, genetic engineering of plants for enhanced synthesis and exudation of natural chelators into the rhizosphere, improvement of the rhizosphere with the help of mycorrhiza and integrated management of the metalliferous ecosystem following the principles of phytoremediation are discussed in this paper. Key words: Metalliferous soils, Rhizosphere, Mycorrhizae, Genetic engineering, Metal sequesteration, Metal hyperaccumulators, Metal tolerant plants The term "serpentine" is applied to minerals viz, lizardite, antigorite and chrysotile (= well known in asbestos-form in some areas). The famous Roman military surgeon Dioscorides recommended it for the prevention of snakebite or the rocks have the speckled colour of serpents. Serpentinized rocks are distributed all over the world viz., western north America; New foundland, Mount Albert in eastern Canada; Lizard peninsula, Wales and Scotland; north-east Cuba; Portugal; Italy; Balkan peninsula; Turkey; topical far east; Central Brazil; New Caledonia; south east Asia; Philippines; Japan; Zimbabwe; eastern Transvaal Loweveld of South Africa, New Zealand; greenstone belts of western Australia In north-east of Portugal the serpentinized area is about 8,000 ha with characteristic geology and flora. The soils contain low N, P, K, Ca, K, Mo and high Cr, Co, Ni and Mg/Ca quotient. The ecology and floristics of this region has been investigated by Sequeira and Pinto da Silva (1991). For the convenience of readers, the slient serpentinophytes are shown in table 1 and figure 1. The Ni content in leaves analyzed from the herbarium specimens of endemics, endemic-Poaceae, and non-endemics of north-eastern Portugal are shown in figures 2, 3, 4 respectively. By proper understanding of the chemical (pH, Eh, CEC, metal speciation), physical (size, texture, clay content, % organic matter), and biological (flora and microorganisms) processes and their interactions, it is possible to initiate appropriate bioremediation measures. Table 1: Important European serpentinophytes (Sequira and Pinto da Silva 1991)
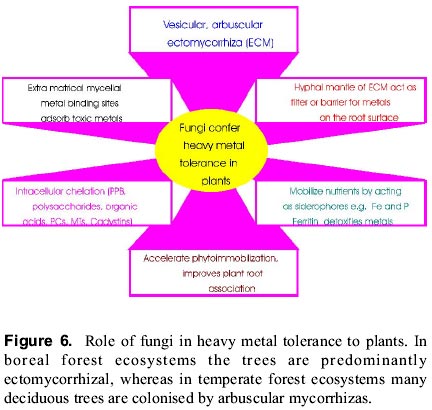
The conventional remediation technologies (other than bioremediation) used for in situ and ex situ remediation are typically expensive and destructive. The include solidification and stabilization, soil flushing, electrokinetics, chemical reduction/oxidation, soil washing, low temperature thermal desorbtion, incineration, vitrification, pneumatic fracturing, excavation/ retrieval, landfill and disposal (Saxena et al 1999; Wenzel et al 1999). Bioremediation, is an integrated management of polluted ecosystem where different organisms are employed which catalyze the natural processes in the polluted or contaminated ecosystem (aquatic or terrestrial). Suitable, but high cost technologies have been identified for clean-up of heavy-metal-polluted soils (Iskandar and Adriano 1997). Recently, the potential role of bioremediation, particularly higher terrestrial plants (phytoremediation) in remediation of metal-polluted soils has been studied by an increasing number of scientists from various disciplines including plant scientists, microbiologist, chemists, soil scientists and geologists owing to its commercial importance (Raskin 1995; Watanabe 1977). Emerging phytoremediation technologies suitable for metal contaminated ecosystems and the integral processes involved and relevance to contemporary environment have been comprehensively reviewed (Saxena et al. 1999; Wenzel et 1999). Recently, Hüttermann and co-workers have used cross linked polyacrylates, hydrogels (Stockosorb K400), to metal-contaminated soils. When Stockosorb K400 was applied to hydrocultures of Scots Pine, Pinus sylvestris, which contained 1 mM of Pb, the hydrogel increased the nutrient efficiency of the plants and, the detrimental effect of the heavy metal was completely remediated (Hüttermann et al. 1999 in Prasad and Hagemeyer 1999). Hydrogel significantly inhibited the uptake of Pb by the fine roots of 3-year-old spruce. Conversely, Anderson et al (1998) have induced Brassica juncea to accumulate gold from ores by treating the substrate with ammonium thiocyanate. Thus, hyperaccumulation can be induced in some plant species by soil amendment using EDTA on an insolube target metal complex such as lead ore, rendering it soluble which otherwise is insoluble. In this review, an update of biotechnological and bioremediation strategies suitable for serpentine soils and mine spoils have been presented in different sections. Ecological improvement of the rhizosphere with the help of mycorrhiza The term "rhizosphere" was refers to the designated the zone of enhanced microbial abundance in and around the roots. The Studies by microbiologists and mycologists improved our understanding of the role of root exudates in the rhizosphere. The soil-root interface is generally called as the rhizoplane. The rhizosphere is represented by few millimeters of soil surrounding the plant roots and influenced by their activities. Because the rhizosphere is characterized by steep gradients of microbial abundance and chemical characteristics, the boundary between rhizosphere and bulk soil is not accurately determined. The rhizosphere is highly dynamic, plant-driven microenvironment which is characterized by feedback loops of interactions between plant root processes, soil characteristics, and the dynamics of the associated microbial populations. In short, it is characterized by very dynamic biogeochemical processes as compared to bulk soil (Wenzel et al 1999). Soil communities are among the most species rich components of terrestrial ecosystems (Giller, 1996). In addition, most of the annual flux of carbon and nutrients occurs in the top 5-10 cm of the soil profile and the magnitude and the chemical nature of these fluxes are controlled by soil organisms and are therefore essential for ecosystems functioning (Wolters and Schaefer, 1994). It is generally accepted that soil diversity is functionally important for buffering against stress related to environment fluctuations, composite habitats and complex substrates. Yet, due to the enormous species richness of soil organisms combined with poor taxonomic information, no complete inventory of soil biota is available for any habitat or region (Lee, 1991). Little quantitative information on the ability of soil microbial communities to recover from perturbation or of the effects such perturbations have on essential ecosystems services. Rhiozphere microorganisms promote the establishment of plant species on mine spoils by immobilizing the heavy metals in the soil, thereby reducing the plant availability of the metals (Chanmugathes and Bollag 1987). This effect is particularly true for one group of soil fungi, viz., mycorrhizae. These mycorrhizal fungi contribute directly to plant establishment in soils contaminated with heavy metals by binding metals to fungal hyphae in roots or in the rizosphere (Wasserman et al. 1987; El-Kherbawy et al. 1989; Koslowsky and Boerner 1989; Gadd, 1993). As a result, plants infected by these fungi show a higher degre of resistance to metals, since metals are complexed in fungal hyphae and are not translocated to shoots (Bradley et al. 1981; Brown and Wilkins 1985; Dehn and Schüepp 1989). A wide range of fungi from all major taxonomic groups are reported in metal-polluted habitats and are capable of surviving and growing in the presence of toxic concentrations of heavy metals (Gadd 1993). Toxic metals are believed to affect fungal populations by reducing abundance and species diversity and selection for a resistant/tolerant population (Babich and Stotzky 1985; Duxbury 1985). The mechanisms by which they are able to deal with these generally toxic metal are numerous and varied in their action (Gadd 1993). Fungi confer tolerance by various mechanisms (Figure 6; Galli et al. 1993, 1994; Turnau et al. 1996) Several reports have shown that mycorrhizal associations are able to ameliorate the toxicity of heavy metals to higher plants (Table 2; Hartley et al. 1997; Johanson and Mc Graw 1988a,b; Tamm 1995). For example ericoid mycorrhizas (Bradley et al. 1981), ectomycorrhizas (Denny and Wilkins 1987; Dixon and Buschena 1988) and for arbuscular mycorrhizas (Gildon and Tinker 1983; Heggo and Angle 1990) are able to ameliorate heavy metal toxicity to higher plants. However, this depends on the mycorrhiza and the metal involved (Galli et al. 1994). For e.g. Scleroderma citrunum, Paxillus involutus, Laccaria laccata have no effect on Pinus sylvestris. However, Suillus bovinus and S. leutus have increased tolerance to Cd (Colpert and Van Asssche 1992). Jones and Hutchinson (1986) showed that Scleroderma flavidum was the only species of four tested which was able to increase tolerance of Betula papyriferae to nickel; none of the fungi they tested increased protection against copper. Brown and Wilkins (1985) found that Zn toxicity to Betula pendula was ameliorated by colonization with ectomycorrhizal fungi. The ameliorating influence of ectomycorrhiza on Zn toxicity to Betula was positively correlated with the degree of compatibility between the fungal strain (Paxillus involutus) and the plant (Denny and Wilkins 1987). Table
2: Mycorrihiza increased metal tolerance, enhanced the growth and
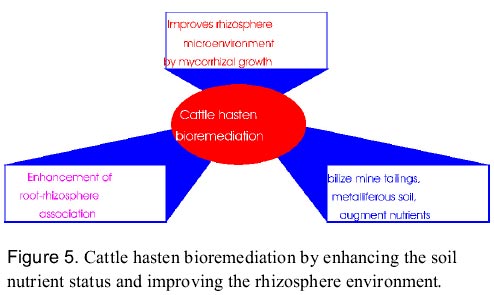
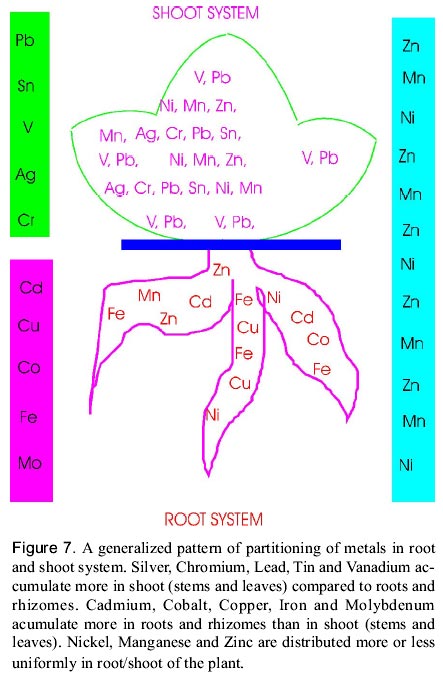
Dueck et al. (1986) have found that the colonization of the two grasses Festuca rubra and Calamagrostis epigejos by VAM fungi reduced the toxic effects of Zn on the growth of both species. Tallgrass prairie species native to Minnesota can withstand drought and grow in nutrient-poor soils and they are likely to benefit from inoculation with arbuscular mycorrhizal (AM) fungi, because tailing deposits are deficient in phosporus and contain low levels of indigenous AM fungal inoculum (Noyd et al. 1995). Some of the decomposer fungi, i.e. Agaricus hyperaccumulate toxic metals like Hg, Pb and Cd compared to some mycorrhizal fungi (Lodenius et al. 1981). Saprophytic fungi decomposes the wood and bark the reservoirs of metal binding ligands like pectins, cell wall polysaccharides and polyphenols etc. and bioconcentrate the toxic metals in large amounts. In copper mines near Globe and Tucson, Arizona, USA (Arizona Ranch, Resource Management and Mine Reclamation; ASARCO Inc. Copper Operations) the ecosystem rehabilitation and mine reclamation programme is primarily based on cattle. Herds of cows are impounded with electric fence on mine tailings, metal contaminated soils by providing fodder and water for varying durations. Cows not only stabilize the soil by their hoofs but also enrich soil nutrient status via urination. They also augment microbes to the soil through the dung. This process is repeated at regular intervals in cycles. Thus, cattle accelerates rhizosphere development and improves plant root association via enriching soil microorganisms and nutrients. The results obtained with cattle for rhizosphere ecodevelopment are spectacular (Figure 5; Dagget 1997) Genetic engineering of metal sequestering, transporting and modifying biomolecules Plants growing in metal contaminated environment would accumulate toxic metal ions and efficiently compartmentalize these into various plant parts. Data gleaned from literature indicate that the partitioning of heavy metals at the whole plant level can be broadly be devided into 3 categories (Figure 7; Prasad 1996, 1998). Uptake of toxic metals, their translocation to plant parts and the plant resistance to them are dependent on metal speciation which can be further modified by the metabolic processes of the plant (Szpunar and Lobinski 1999, in Prasad and Hagemeyer 1999). Primarily two types of mechanisms may confer resistance to the toxicity of metal ions in plants. They are (i), avoidance, involving various ways of preventing toxic ions to reach their target sites, and (ii), tolerance to metal ions in symplasm by complexation (Prasad 1997, 1988; Rengel 1997). The prominent metal complexation processes being the synthesis of phytochelatins and metal chelating peptides (Prasad 1997; Rauser 1990, 1995; Reddy and Prasad 1990; Rengel 1997; Steffens 1990). Different types of metal-chelating compounds and processes have been implicated in the regulation of the intracellular metal ion concentration. The biosynthesis of these compounds, e.g. phytochelatins is enzymatically mediated, i.e. requires a protein activated by the metal of interest. The heavy metal deposition pattern has been correlated with forest decline and the concentration of phytochelatins (Gawel et al. 1996). It has also been reported that certain plants function as hyperaccumulators of specific heavy metals (Brooks 1998; Reeves et al. 1995; Krämer et al. 1996) owing to their efficient metal complexation processes (Prasad 1998). The identification of the complex of the metal with the enzyme and the characterization of the metal complexes with products of the enzyme-catalyzed reaction is an emerging field of research in environmental speciation analysis. a) Genetic engineering for metal-sequestering proteins and peptides Phytochelatins (class III metallothioneins) are short metal-induced sulfhydryl-rich peptides possessing the general structure: (g -GluCys)n-Gly with n = 2-11. They are synthesized from glutathione in plants and fungi exposed to Ag, Bi, Cd, Cu, Hg, Ni, Sn, Sb, Te, W, Z and anions such as SeO4-2, SeO3-2 and AsO4-3. The non-inducers of phytochelatins are Na, Mg, Al, Ca, V, Cr, Mn, Fe, Co and Cs (Rauser 1995). Metals are chelated through coordination with the sulfhydryl group in cystein. An intracellular complex formed by these thiol peptides is thought to detoxify the metal by sequestration in the vacuole (Rauser 1995; Zenk 1996). Metal sequestering complexes in plants have been named differently by different authors viz. cadystin, poly (g -glutamyl-cysteinyl) glycine, phytometallothioneins, g -glutamyl metal-binding peptide, g-glutamyl cysteinyl isopeptides, metallothiopeptide, metallopeptides, des glycyl peptides (g -glu-cys), phytochelatins (Prasad 1999, in Prasad and Hagemeyer 1999). Glutathione is the precursor of phytochelatin synthesis. These peptides bind metal ions and form stable metal binding complexes that are transported and sequestered in the plant vacuoles. Glutathione and its homologues viz., homoglutathione and hydroxymethyl-glutathione are the abundant low molecular weight thiols in plants. Sulphur rich compounds are known to reduce the metal toxicity probably by influencing gluathione metabolism (Csermely 1998). Metal induced phytochelatin production decrease cellular levels of glutathione. Alteration of thiol pools of metal exposed plants could be regulated by intermediates and effectors of glutathione synthesis. Hence, there exists a possibility of increasing the level of these metal-binding peptides in plants by increasing the level of GSH. This could be achieved by upregulating the expression of the enzymes responsible for GSH synthesis. Recently, poplar trees have been transformed by introduction of the gene coding for the bacterial enzyme g-glutamylcysteine synthetase, the key enzyme in the synthesis of GSH. Some of the transgenic plants obtained showed higher levels of GSH and are currently being assayed for their heavy metal tolerance and uptake (Arisi et al. 1997). In an alternate approach, synthetic phytochelatin analogs (Glu-Cys)nGly peptides were tested for their metal binding properties. These synthetic peptides have been shown to exhibit metal-chelating properties similar to natural phytochelatins (Bae and Mehra 1997). Synthetic genes coding for these unnatural peptides are being tested now aiming the production of metal resistant trangenic plants. Class I and II metallothioneins are another group of non-enzymatic polypeptides with low molecular mass (6-7 kDa), rich in cysteinyl residues and able to complex metal ions having sulphur affinity. This group of proteins are characterized by their resistance to thermocoagulation and acid precipitation, by the presence of ca. 60 non-aromatic amino acids, and by the absence of disulfide bonding (Stillman et al. 1992; Stillman 1995). Although MTs are most common in the animals, several plants are reported to contain MT genes viz., Pea (Pisum sativum), Soybean (Glycine max), Thale cress (Arabidopsis thaliana), Monkey flower (Mimulus guttatus), Maize (Zea mays), Barley (Avena sativa), Wheat (Triticum aestivum), Castor bean (Ricinus communis) and Rape seed (Brassica napus). Chinese hamster MT-II is expressed and functional in Brassica campestris leaf tissue when the plant is infected with a cloned CMV pCa-BB1 containing cDNA of the MT-II gene. Heavy metal tolerant transgenic Brassica napus and Nicotiana tabacum plants were obtained by infecting with Agrobacterium tumefaciens containing human MT-II processed gene on a disarmed Ti plasmid. Mouse MT gene expression was studied in transgenic tobacco (Prasad 1995). The use of MT genes for phytoremediation purposes can be further enhanced by altering their metal binding specificity in order to increase their affinity to specific metal ions. The transfer of mammalian MT genes to higher plants has been successfully demonstrated and provided some benefit to plants by increasing their metal tolerance (Brandle et al. 1993, Hattori et al. 1994, Pan et al. 1994). However, in such transgenic plants, the synthesis of metal inducible peptides/proteins and their role in metal tolerance warrant detailed investigation. Sczekan and Joshi (1989) illustrated that ferritin binds non-ferrous metal ions both in vitro and in vivo. Hence, induction of ferritins in cultivated and wild plants would be useful in mitigating heavy metal toxicity. Kumar and Prasad (1999) reported the metal binding properties ferritin in Vigna mungo (Black gram) and suggested its role in heavy metal detoxification (Table 3). Table 3: Metal detoxification mechanisms in plants (Prasad 1998)
b) Genetic engineering for encoding metal ion transporters Saxena et al. (1999) indicated that the most promising approaches for increasing the metal uptake potential of plants is through the control of ion transport across root cell plasma membranes. Metal transport-complexes of plant cell walls, increased protection of sulphydril groups in channel proteins of metal exposed plants, symplasmic and tonoplast transporters characterization is necessary for further progress in genetic engineering (Ernst et al. 1992). Metal tolerant plants are reported to contain elevated concentrations of organic acids (Table 3). Several of the metal-ion transporters have already been isolated, characterized and, in some cases, cloned. Through functional complementation in yeast, Eide et al. (1996) have isolated an Arabidopsis IRT1 gene, identified as being a plasma membrane iron (Fe2+) transporter involved in iron uptake by the roots. They suggested that cadmium might serve as a substrate for this transporter. Recently, Grotz et al. (1998) have isolated from Arabidopsis three ZIP genes encoding putative Zn transporters, similar to the ZRT genes of Saccharomyces cerevisiae. The Arabidopsis COPT1 gene can complement a yeast ctr1 mutant deficient in copper uptake, suggesting that this gene may encode a copper transporter (Kampfenkel et al. 1995). Overexpression of these ion transporters can be used to enhance heavy metal uptake by transformed plant tissues. Moreover, the specificity of the transporters could be altered (through point mutation for example) in order to increase the pumping of metal ions such as lead, cadmium, mercury or even radionuclides. c) Genetic engineering of enzymes to enhance phytovolatilization Saxena et al. (1999) indicated that metals like mercury, selenium, arsenic or chromium can be rendered harmless by either enzymatic reduction or by incorporation into less toxic organic/metal compounds. These processes occur in nature and can be enhanced by genetic manipulation of plants through introduction of genes coding for enzymes responsible for the underlying biochemical reactions. A well-known example of such manipulation is the transfer and expression of a modified bacterial Hg2+ reductase gene in transgenic Arabidopsis thaliana plants (Rugh et al. 1996). Plants containing this new gene were more tolerant to the presence of mercury, compared to plants lacking it. The transgenic plants reduced Hg2+ to elemental mercury, which is easily volatilized at room temperature. Rugh et al (1998) also examined the ability of yellow poplar (Liriodendron tulipifera) tissue cultures and plantlets to express modified mercuric reductase (merA) gene constructs. Bacteria possessing merA are capable of converting highly toxic, ionic mercury , Hg(II), to less toxic elemental mercury , Hg (0). Thus, expression of merA in trangenic plants or identification of plants ecotypicaslly expressing such genes would certainly be helpful for ecologically compatible remediation options. Similarly, some plants can also volatilize selenium (Azaizeh et al. 1997; Terry and Zayed 1994). The selenium accumulator Astragalus racemosus releases the metal to the air as dimethyl diselenide avoiding the formation of toxic selenocysteine and selenomethionine. Metal hyperaccumulator plants form an important biological resource and their introduction to metal contaminated and polluted areas would be helpful in revegetation and phytoremediation (Table 4; Liston and Balkwill 1997). Considerable progress has been achieved in this direction by converting metal hyperaccumulation wild plants into commercial phytoremediation systems. Amelioration of soils prior to the introduction of tolerant plant species may lead to greater success. Immobilization of heavy metals in polluted soils by application of a modified alumino-silicate; amelioration of soil Al toxicity by Si have been reported (Hodson and Evans 1995). Infact for amelioration of Ni contaminated soils, lowering of pH with ammoniacal fertilizers or elemental sulphur will be most effective favoring Ni availability and bioconcentration in plant tissues (Sumner and Naidu 1997). Chemical remediation treatments have been suggested for soils contaminated with cadmium and lead. Toxic metals have been inactivated in polluted soils using natural zeolite and apatite. Application of synthetic chelates and zeolites have triggered lead hyperaccumulation in plants and hastened reclamation of metal contaminated sites (Baylock et al. 1997; Huang et al. 1997). However, the long term effects and cost benefits of usage of synthetic zeolites as amendments for metal polluted soils need critical investigation. Thus, biotechnology for bioremediation of metal contaminated and polluted soils involves integration of several aspects of various disciplines which need to be addressed concurrently for optimization of the results. Table 4: Data gleaned from literature clearly indicate that the enumerated plants are reported to be metal hyperaccumulators/tolerant to heavy metals. These would be useful for phytoremediation of metal contaminated/polluted ecosystems and further experimentation.
MNVP is thankful to Prof Richard S. Gordon, School of Agribusiness and Resource Management Arizona State University East, Mesa AZ, USA for hosting and arranging field study visits to copper mines in the vicinity of Tucson and Globe. Thanks are due to Mr Stuart A. Bengson, Mines Land Reclamation Cosultanting in Ecosystem Rehabilitation, ASARCO Inc. Copper Operations, Tucson AZ, USA and Mr Terence O. Wheeler, President and General Manager, Arizona Ranch Management and Natural Resurce Management and Consulting, Globe, AZ, USA for demonstrating the cattle reclaimed copper mine tailings.
Supported by UNESCO / MIRCEN network © 1999 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej99004f5.jpg] [ej99004f7.jpg] [ej99004f6.jpg] [ej99004f1.jpg] [ej99004f2.jpg] [ej99004f3.jpg] [ej99004f4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}