 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic Journal of Biotechnology, Vol. 2, No. 3, December, 1999 INVITED RESEARCH ARTICLE VNTR-based diversity analysis of 2x and 4x full-sib Musa hybrids Jonathan H. Crouch *1, Hutokshi K. Crouch2, Abdou Tenkouano3 and Rodomiro Ortiz4 1Elsoms
Seeds Ltd., Spalding, Lincolnshire, PE11 1QG, England, United Kingdom E-mail
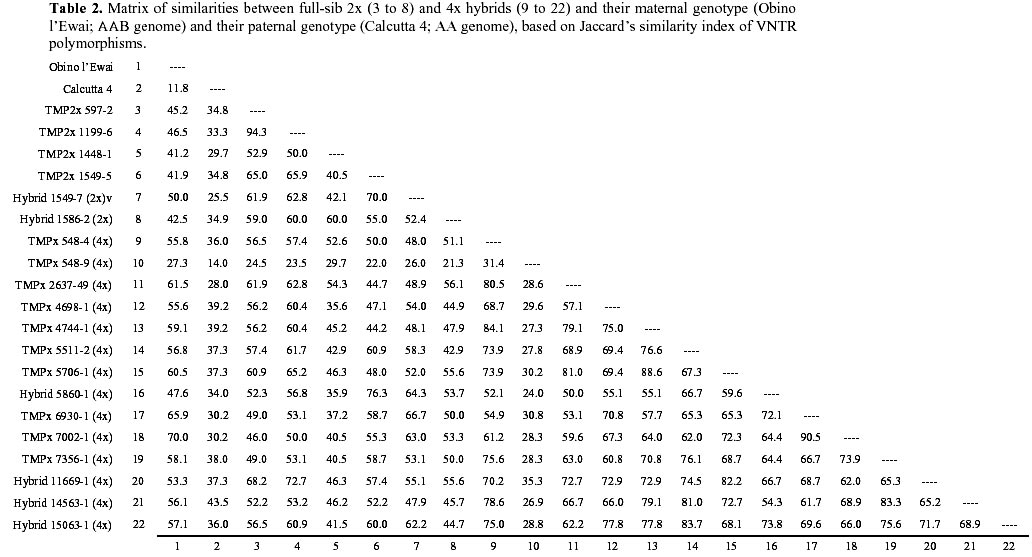
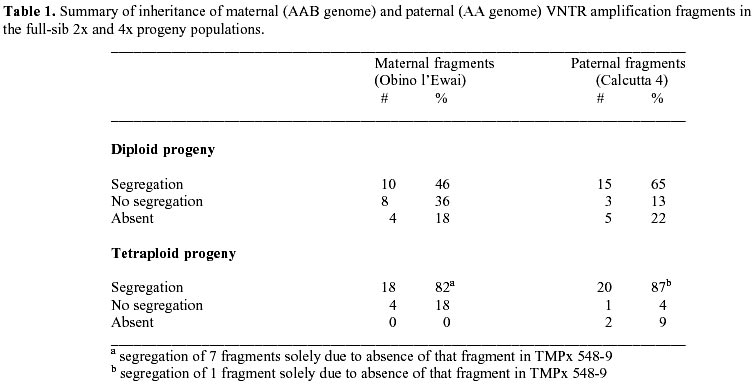
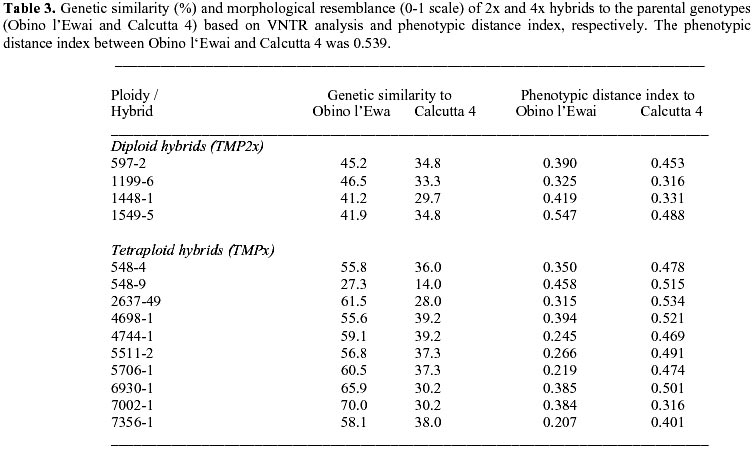
: jonathanc@elsoms.co.uk Received June 24, 1999 Code Number: ej99015 The triploid plantain landrace Obino l’Ewai (Musa spp., AAB genome) has been crossed with a wild diploid banana (M. acuminata subsp. burmannica var. ‘Calcutta 4’, AA genome) to generate full-sib diploid and tetraploid hybrids combining good agronomic performance and disease resistance. Microsatellite marker analysis of the parental genotypes confirmed the highly heterozygous nature of both parental genotypes. Comparative analysis of 2x and 4x full-sib hybrids with their parental genotypes indicated that tetraploid hybrids are generally more closely related to Obino l’Ewai than their diploid full-sibs. Based on VNTR analysis it is possible to identify those hybrids, which may be most useful in subsequent breeding of secondary triploid hybrids. There was a significant (P<0.05) negative association between the VNTR-based genetic similarity of hybrids to Obino l’Ewai and a phenotypic distance index based on eight agronomic descriptors. However, there was no association between the molecular genetic similarity of hybrids to Calcutta 4 and the respective phenotypic distance index. Many microsatellite markers generated an unexpectedly high number of amplification products from AA and AAB genotypes plus their progeny which may suggest the presence of a high frequency of loci duplication in both A and B genomes, in addition to the detection of heterozygous and/or homoeologous loci. Keywords : Breeding, Diversity, Hybrids, Musa, VNTR. Plantain (Musa spp., AAB group) is a triploid (2n = 3x = 33) giant perennial herb of considerable importance to the agriculture of tropical humid forest regions in Africa, America and Asia (FAO 1999; Robinson 1996). In terms of global production value, Musa crops are ranked as the fourth most important food commodity after rice, wheat and milk (Ortiz and Vuylsteke 1996). Yet despite the importance of these crops for international trade and food security in tropical regions, little attention has been given to their genetic improvement in comparison to other major food crops. As a consequence, major advances in Musa productivity have traditionally relied on improvements in crop husbandry. However, promising tetraploid hybrids have recently been generated by several breeding programs across the world. In all cases these have been produced through interspecific hybridisation of triploid landraces with diploid accessions from wild, cultivated or improved genepools (Rowe and Rosales 1993; Vuylsteke et al. 1993b; Dantas et al. 1995; Vuylsteke et al. 1995). Nevertheless, there remains an enormous potential for increasing yields in Musa crops through genetic improvement (de Vries et al. 1967). Considerable advances have recently been made in the understanding of the genetic basis of specific traits in Musa (Ortiz 1995; 1997a). However, there is an urgent need to make similar progress in our understanding of the genomic structure and genetic relationship of the parental and progeny genotypes used in Musa breeding programs across the world. Analysis of diploid and tetraploid progeny from the same parental genotypes offers a unique opportunity in the study of Musa breeding schemes. Furthermore, current efforts concerning the genetic improvement of plantain at IITA are focused on the production of secondary triploid hybrids from crosses between diploid and tetraploid accessions (Ortiz et al. 1998). For this reason there is considerable interest in defining the genetic relatedness of the elite diploid and tetraploid hybrids already available, as a possible criterion for selection of parental genotypes in triploid hybrid breeding schemes. Microsatellite markers have proven useful for genetic analysis in a number of systems (Powell et al. 1996) including Musa (Kaemmer et al. 1997; Crouch et al. 1998). This study uses variable number of tandem repeats (VNTR) analysis of microsatellite loci to compare the genetic similarity of full-sib 2x and 4x hybrids, and their parental genotypes. The implications of this analysis for the breeding of polyploid Musa cultivars, and for the evolution of the Musa genome are discussed. Materials and MethodsPlant material. A total of 990 bagged inflorescences of the triploid plantain landrace Obino l’Ewai (AAB) were pollinated with the diploid wild banana M. acuminata subsp. burmannica var. ‘Calcutta 4’ (AA). These crosses generated 4871 seeds, from which 16 tetraploid hybrids and 22 diploid hybrids were recovered. No other genotypes survived to field establishment, although it is known that triploid and aneuploid regenerants can be obtained at low frequency from such crosses. From the 38 hybrids, 14 tetraploid hybrids and 6 diploid hybrids were selected due to their agronomic performance (Vuylsteke et al. 1993a; Vuylsteke and Ortiz 1995). The ploidy level of these hybrids was confirmed through morphological evaluation and chromosome counts (Vandenhout et al. 1995). The parental genotypes ‘Obino l’Ewai’ and ‘Calcutta 4’ together with the following diploid hybrids were used: TMP2x 597-2, TMP2x 1199-6, TMP2x 1448-1, TMP2x 1549-5, hybrid 1549-7 and hybrid 1586-2. In addition, the following tetraploid hybrids were used: TMPx 548-4, TMPx 548-9, TMPx 2637-49, TMPx 4698-1, TMPx 4744-1, TMPx 5511-2, TMPx 5706-1, hybrid 5860-1, TMPx 6930-1, TMPx 7002-1, TMPx 7356-1, hybrid 11669-1, hybrid 14563-1 and hybrid 15063-1. Leaf tissue was harvested from parental and hybrid plants grown in Onne, Nigeria at the IITA High Rainfall Station, and used for isolation of DNA for VNTR analyses as described previously (Crouch et al. 1998). Multiple preparations from parental genotypes were stored and used separately (there was no bulking of DNA from different individuals or different samples of the same genotype). Generation of PCR primers and microsatellite amplification. An accession of M. acuminata subsp. malaccensis was used for generating a genomic library (Jarret et al. 1994). Clones from this library which contained microsatellites were sequenced and primers designed from flanking regions as described previously (Crouch et al. 1998; 1999), to generate microsatellite markers with Ma prefixes, viz. Ma 0-1, Ma 0-3, Ma 0-5, Ma 0-7, Ma 0-9, Ma 1-0, Ma 1-2, Ma 1-3, Ma 1-5, Ma 1-6, Ma 1-7, Ma 1-9, Ma 1-11, Ma 1-12, Ma 1-16, Ma 1-17, Ma 1-18, Ma 1-19, Ma 1-23, Ma 1-24, Ma 1-27, Ma 1-29, Ma 1-31, Ma 1-32, Ma 2-0, Ma 2-3, Ma 2-4, Ma 2-4B, Ma 2-5, Ma 2-7, Ma 2-8, Ma 2-10, Ma 2-12, Ma 2-23, Ma 3-1, Ma 3-1N, Ma 3-2, Ma 3-41, Ma 3-46, Ma 3-48, Ma 3-50, Ma 3-55, Ma 3-57, Ma 3-59, Ma 3-60, Ma 3-63, Ma 3-64, Ma 3-68, Ma 3-77, Ma 3-79, Ma 3-81, Ma 3-87, Ma 3-90, Ma 3-92, Ma 3-103, Ma 3-104, Ma 3-109, Ma 3-111, Ma 3-127, Ma 3-130, Ma 3-131, Ma 3-132, Ma 3-139, Ma 3-161. These microsatellite markers cover a wide range of core motifs including perfect di- and trinucleotide repeats, compound dinucleotide repeats plus imperfect di- and trinucleotide repeats (comprising GA, AT, CT, GAA, CTT and ATT) as described previously (Crouch et al. 1997). Primer sequences for additional microsatellite markers with prefix CIR or prefix STMS have been reported elsewhere, viz. CIR 37, CIR 38a, CIR 38b, CIR 276, CIR 1113, CIR 332a, CIR 327a, CIR 327b, CIR 631a, STMS 7, STMS 8, STMS 9, STMS 12, STMS 14, STMS 15. (Kaemmer et al. 1997). All primers were synthesised by MWG-Biotech (Germany).The PCR samples consisted of 25 ng template DNA, 1.2 m M each of forward and reverse primer, 10 mM tris-HCl (pH 9), 2.5 mM Mg2+, 0.2 mM each dNTP and 1 unit Taq polymerase (Appligene) in a reaction volume of 15 m l. Reaction components were initially denatured for 4 min. at 94° C followed by 30 amplification cycles each consisting of 1 min. denaturation at 94° C, 1 min. annealing at primer melting temperature (specific for each primer, see Kaemmer et al. 1997; Crouch et al. 1998; 1999), 45 sec. extension at 72° C. Amplifications were carried out using a Perkin Elmer thermal cycler model 9600. PCR products were separated electrophoretically using 1.5% w/v Nusieve GTG (FMC) + 1.5% w/v Metaphor (FMC) + 0.5% Multipurpose (Appligene) agarose gels containing 0.3 m g/ml ethidium bromide, in 1 x TBE buffer at 5V/cm for approximately 4 h. Amplification products were visualised and photographed using UV illumination. The following primers generated amplification products when pre-screening the parental genotypes: Ma 0-9, Ma 1-2, Ma 1-16, Ma 1-17, Ma 1-19, Ma 1-24, Ma 1-27, Ma 1-29, Ma 2-4, Ma 2-4B, Ma 2-7, Ma 2-10, Ma 3-1N, Ma 3-41, Ma 3-46, Ma 3-48, Ma 3-50, Ma 3-55, Ma 3-59, Ma 3-60, Ma 3-64, Ma 3-77, Ma 3-79, Ma 3-81, Ma 3-90, Ma 3-92, Ma 3-103, Ma 3-104, Ma 3-109, Ma 3-127, Ma 3-132, Ma 3-139, Ma 3-161, CIR 37, CIR 38a, CIR 38b, CIR 276, CIR 1113, CIR 332a, CIR 327a, CIR 327b, CIR 631a, STMS 7, STMS 8, STMS 9, STMS 12, STMS 14, STMS 15. Statistical analysis. The dataset from microsatellite marker screening across parental genotypes plus diploid and tetraploid progeny was subjected to similarity analysis based on Jaccard’s index (Jaccard 1908), to derive a matrix of similarity coefficients (Table 2). Pairwise comparisons from this similarity matrix were used to generate a dendrogram of genetic relatedness (Figure 1). In this analysis we wished to focus on the differences between genotypes. Thus, markers and alleles which failed to distinguish differences between the parental genotypes or within the diploid or tetraploid progeny populations have been excluded from this analysis. However, a small degree of similarity between Obino l’Ewai and Calcutta 4 is still observed (Table 2). This similarity results from the absence in both parents of amplification products, which were observed in certain progeny individuals (i.e. non-parental bands). All analyses were carried out with the aid of a program run within the Genstat software (Payne et al. 1989). A distance index was developed to compare the phenotypic diversity between the parental genotypes and a subset of four diploid and ten tetraploid hybrid offspring, which have been released as improved germplasm by IITA (Vuylsteke et al. 1993b; Vuylsteke and Ortiz 1995). The phenotypic distance index was based on the eight morphological descriptors reported in the registration of this germplasm: days to flowering, plant height (cm), height of tallest sucker (cm), days for fruit filling, bunch weight (kg per plant), number of hands, fruits per hand, and average fruit weight (g). The phenotypic distance between each hybrid and its respective parents was calculated as the difference in the morphological descriptor values divided by the overall range for that descriptor (Gower 1985), thereby generating phenotypic distances on a 0 to 1 scale. The individual descriptor distances were summed and then divided by 8 (viz. the total number of morphological descriptors used).Genetic analysis of parental genotypes A total of 79 primer pairs (hereafter referred to as primers) were used to screen the parental genotypes (Obino l’Ewai and Calcutta 4). Forty-eight of these primers generated amplification products of which 34 (71%) detected polymorphisms between these two genotypes. A total of 119 amplification products were generated by the 48 primers of which 70 (59%) distinguished the two parental genotypes. Twenty-seven primers (56%) detected two or more alleles in Calcutta 4, suggesting the presence of heterozygous loci. A similarly high level of heterozygosity (48%) of this apparently true breeding line has been reported previously based on a limited data set (Crouch et al. 1998). However, in the present study eight primers (17%) identified three or four alleles in Calcutta 4; Ma 1-2, Ma 1-17, Ma 1-19, Ma 3-48, STMS 5, STMS 15, CIR 37, CIR 631a. This finding suggests a high frequency of duplicated alleles or duplicated chromosomal regions in the diploid (AA) genome of Calcutta 4. Hence, markers detecting multiple alleles may be detecting loci which are heterozygous and/or duplicated. Thus, our previous estimate for the heterozygosity of Calcutta 4 (Crouch et al. 1998) is likely to be a significant over estimate of the true level in this clone. Similarly, the majority of primers detected from one to three alleles in Obino l’Ewai, which is consistent with the heterozygous triploid nature of this accession. However, four primers (8%) revealed four or five alleles in Obino l’Ewai; Ma 3-48, STMS 8, STMS 9, CIR 631a, again, suggesting a high frequency of duplicated alleles or duplicated chromosomal regions in the allotriploid (AAB) genome of Obino l’Ewai. Finally, four primers (Ma-3-41, Ma-3-60, Ma-3-161 and CIR327b) detected one or two alleles in Obino l’Ewai but failed to generate amplification products from Calcutta 4. In contrast, two primers (Ma-1-29 and Ma-3-64) detected one or two alleles in Calcutta 4 and none in Obino l’Ewai. This finding may suggest the occurrence of single base changes or larger alterations in the genomic regions homologous to the primer sequences in these two genotypes as compared to the M. acuminata accessions from which the microsatellites were isolated. Evolutionary divergence of accessions in this way is well documented, particularly as a consequence of genetic drift (Gillespie 1991). Segregation patterns in progeny populationsSeventeen primers detected polymorphisms between Obino l’Ewai and Calcutta 4 where the alleles had very large differences in rate of migration, and, therefore, were easy to routinely score unambiguously; Ma 1-16, Ma 1-17, Ma 1-19, Ma 1-24, Ma 1-27, Ma 2-7, Ma 2-10, Ma 3-48, Ma 3-90, Ma 3-139, STMS 7, STMS 8, STMS 14, STMS 15, CIR 276, CIR 38a, CIR 631a. These primers were used to screen the full-sib population of 14 tetraploid hybrids and 6 diploid hybrids, generating a total of 74 amplification products. A summary of segregation patterns is shown in Table 1. Forty-five of these bands (61%) were present in only one of the parental genotypes while eight (11%) were monomorphic across all parental and progeny genotypes. A further six of the 74 bands (8%) were present in both parental genotypes but segregated in the diploid and/or tetraploid progeny populations. The remaining 15 bands (20%) were not present in either parent but five (7%) segregated in both diploid and tetraploid progeny populations while nine segregated only in the tetraploid population and one segregated only in the diploid population. DNA for this study was extracted from individuals clonally propagated from those used for the original crosses. However, estimates of somatic and somaclonal variation for these genotype based on the analysis of phenotype (Vuylsteke et al. 1988) and RAPD (unpub. data) are considerably lower than the frequency of non-parental bands observed in this study. Genetic and phenotypic similarities between progeny and parental genotypesMost of the tetraploid hybrids were grouped together by the similarity analysis based on Jaccard’s index (Figure 1). Within this group of tetraploid hybrids, TMPx 6930-1 and TMPx 7002-1 are most closely related to the maternal genotype (Obino l’Ewai). In contrast, all the diploid hybrids form two groups associated at the 60% level of similarity, which are distinct from both parental genotypes (Figure 1). The major exceptions are TMPx 548-9, which is separated at the 20% level of similarity from all other genotypes tested and hybrid 5860-1 (4x) that is grouped with the diploid progeny. Hybrid 5860-1 (4x) was found to have 75% similarity to TMP2x 1549-5, both of which have 65% similarity to hybrid 1549-7 (2x) (Figure 1). This anomaly may be a consequence of hybrid 5860-1 (4x) possessing a high proportion of bi-allelic loci in common with these particular diploid full-sibs, as compared to multi-allelic loci of the other tetraploid hybrids. Clearly, this type of molecular marker analysis can not be used for inferring hybrid ploidy. Finally, TMP2x 1448-1 and hybrid 1586-2 (2x) appear particularly distinct from the majority of their 2x and 4x full-sib hybrids (Table 2). Both diploid and tetraploid hybrids had comparable mean levels of similarity to Calcutta 4 (32.2% and 34.3 %, respectively). However, tetraploid hybrids had more fragments in common with Obino l’Ewai than their diploid full-sibs (56.1% and 44.6%, respectively). This relationship was expected because the tetraploid hybrids have 75% of their genes donated by the maternal plantain 2n gametes (and the remaining 25% from the diploid banana), while their diploid full-sibs inherit 50% of their genes from each parent. The average phenotype of the tetraploid hybrids was also more closely related to Obino l’Ewai than Calcutta 4 (Table 3). The phenotypic distance between the tetraploid hybrids and Obino l’Ewai was 0.322 (±0.027), while the phenotypic distance between the tetraploid hybrids and Calcutta 4 was 0.470 (±0.021). Conversely, the phenotypic distance indices for comparisons of diploid hybrids with Obino l’Ewai and Calcutta 4 were very similar; 0.42 (±0.046) and 0.397 (±0.043), respectively. This was expected based on the gametic contributions to diploid and tetraploid hybrids described above. For the relationship between hybrids and Obino l’Ewai, there was a significant association between the genetic similarity (based on VNTR) and the phenotypic resemblance (r = -0.546, P = 0.042). The linear relationship (R2 = 0.298, F = 5.08, P<0.05) between the two measurements was as follows: Sp = 0.590 – (0.456 x Sg) where Sp = phenotypic distance to Obino l’Ewai, and Sg = genetic similarity to Obino l’Ewai. On this basis, the genetic similarity of hybrids to Obino l’Ewai explained only 30% of the phenotypic resemblance between hybrids and this plantain landrace. This poor relationship may be a consequence of the phenotypic distance index being based mainly on components of yield while the microsatellite marker analysis covers a much greater extent of the entire genome. Genetic constitution of progeny genotypesSeven primers (Ma 1-16, Ma 1-17, Ma 1-19, Ma 3-48, STMS 15, CIR 276, CIR 631a) (41%) generated three or four amplification products per individual within the full-sib diploid hybrid population. For the majority of primers, this high multiplex ratio included the presence of multiple alleles donated by Calcutta 4 or Obino l’Ewai. Thus, genomic duplications in the parental genotypes are consistently observed in their interspecific progeny populations where they are observed to segregate normally. Most primers generated between one and four amplification products per individual within the full-sib tetraploid hybrid population, which is consistent with a heterozygous allotetraploid genotype. However, two primers (Ma 3-48, CIR 631a) generated five bands in certain tetraploid hybrids and this further supports the hypothesis of abundant duplications in the Musa genome (in addition to the presence of heterozygous and homoeologous loci). TMPx 548-9 was found to be highly unrelated to all parental and hybrid genotypes (Table 2). This lack of relationship is largely due to the unique absence in this hybrid of a high proportion of bands observed in Obino l’Ewai (Table 1) and its possession of eight unique non-parental bands. Replicated VNTR-based diversity analyses involving independent DNA isolations from this accession indicate a consistent divergence of this hybrid from its full-sibs (unpub. data). DiscussionBased on the data generated from this study, we propose that there is a significant frequency of duplicated loci in both Calcutta 4 (AA) and Obino l’Ewai (AAB). The duplication of genes (and also of entire chromosomes) has been reported in many crop species, where it is often indicative of evolutionary polyploidization (Leitch and Bennett 1997). The Musaceae consists of two genera containing species with several chromosome numbers. The Musa genus comprises four sections, Eumusa and Rhodoclamys (2n = 22) plus Australimusa and Callimusa (2n = 20) while the Ensete genus has only 2n = 18 species. Cultivated and wild bananas have a basic chromosome set of x = 11. However, it has been postulated that this is a secondary haploid number derived from an x = 8 progenitor (Cheesman 1962). A high proportion of loci segregated within the full-sib tetraploid progeny population (Table 1). This result confirms the high level of heterozygosity of both parental genotypes and the high rate of recombination during the formation of 2n (=3x) megaspores by Obino l’Ewai (Crouch et al. 1998). Musa breeding strategies are commonly based on crosses between diploid and triploid accessions, which only yield small progeny populations. Nevertheless, hybrids with up to 70% similarity to Obino l’Ewai but also expressing partial black sigatoka resistance could be selected even from this limited population size (Vuylsteke et al. 1993b). This result is particularly significant as the expression of this disease resistance character has a complex genetic basis, being the result of recessive and additive genes inherited from both parents (Ortiz and Vuylsteke 1994). Thus, these data demonstrate that even when moderate population sizes are used, the level of segregation and recombination observed is sufficient to facilitate efficient introgression of important agronomic characters from exotic germplasm. Implications for molecular breeding of plantainAnalysis of breeding populations with microsatellite markers clearly holds considerable promise for improving the design and operating efficiency of Musa breeding schemes. In this study we have demonstrated the ability of microsatellite markers to detect very high levels of polymorphism in full-sib Musa breeding populations. Over three-quarters of the functional microsatellite markers detected polymorphisms within the breeding populations studied here. Musa improvement programs across the world have concentrated on the generation of diploid and tetraploid hybrids such as those studied here. However, Musa breeding at IITA is currently focused on crossing diploid and tetraploid hybrids in order to generate secondary triploid Musa hybrids suitable for cultivar release (Ortiz et al. 1998). TMP2x 1448-1 and hybrid 1586-2 (2x) were found to be particularly dissimilar to all accessions studied here, forming an out-group at the 45% level of similarity (Figure 1). TMPx 6930-1 and TMPx 7002-1 have also been grouped separately from the other full-sib tetraploid hybrids at the 65% level of similarity (Figure 1). Based on this dissimilarity, the hybridisation of these two diploid hybrids with the two genetically diverse full-sib tetraploid hybrids would be an appropriate strategy for capturing genetic diversity in the resultant secondary triploid progeny. Similarly, a number of other specific pairwise comparisons with similarities below 50% may also be identified (Table 2). However, it is likely that specific genetic factors (as opposed to broad genetic diversity) also have a major contribution to high yield in polyploid Musa hybrids (Ortiz 1997a, b). On this basis two strategies will be important in breeding secondary triploid Musa hybrids: the generation of breeding populations presenting diverse heterotic groups (at the 2x and 4x levels), and, the utilisation of recombination to form genetic structures more favourable than those already present in plantain landraces. Progress in breeding Musa hybrids with ever increasing yields will depend on the ability of breeders to identify and access the genes and gene combinations most likely to generate enhanced heterosis in yield components. Precise selection of prospective male and female parents is a critical step for determining success in achieving significant genetic gains. Parental selection is traditionally carried out a posteriori based on progeny testing. However, this approach cannot be routinely used for Musa breeding due to the large land requirements of this crop (6m2 plant-1) which would render this option inefficient both in terms of time and cost. PCR-based analyses offer unique opportunities for the dissection of genetic relationships among breeding lines. Thus, the relationship between genetic divergence at the molecular level and hybrid performance can now be assessed and may provide an a priori means for parental selection in Musa breeding. Finally, we report a high multiplex ratio for microsatellite marker analysis of diploid and polyploid genotypes plus their hybrids, which may result from the presence of genomic duplications and homoeologous loci. This complex genomic structure confounds the reliable definition of allelic relationships for a large proportion of microsatellite loci. Thus, for many applications, pre-screening will be necessary to define a subset of markers which detect genome specific single copy loci. Only in this way will it be possible to generate co-dominant microsatellite marker information in Musa. This type of marker information will become increasingly important, as for example, attempts are made to accurately define combining ability of potential parental genotypes in triploid Musa breeding, and to apply marker assisted selection in progeny populations. AcknowledgementsWe thank Dr. C. Fatokun and Dr. H.D. Mignouna (IITA) for critical review of the manuscript.
Supported by UNESCO / MIRCEN network © 1999 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej99015f1.jpg] [ej99015t3.jpg] [ej99015t1.jpg] [ej99015t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}