 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
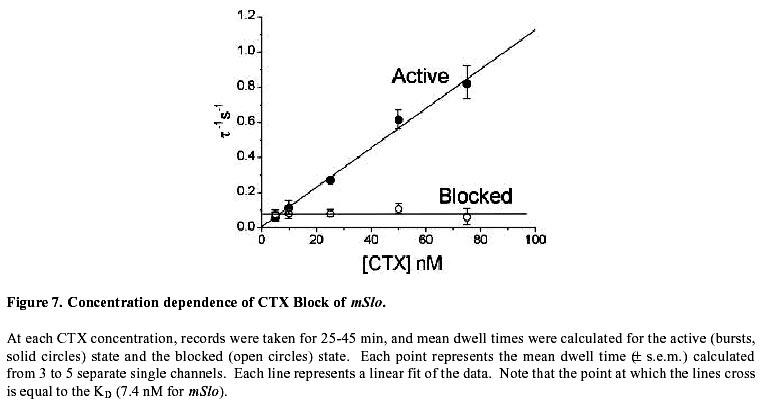
Electronic Journal of Biotechnology, Vol. 2, No. 3, December, 1999 Visualization and functional analysis of a maxi-K channel (mSlo) fused to green fluorescent protein (GFP) Michael P. Myers *1, Jay Yang2 and Per Stampe3 1Department
of Pharmacology and Physiology, University of Rochester School of Medicine and
Dentistry, Rochester, New York, USA 14642-8711. E-mail : adric@ucla.edu
Received
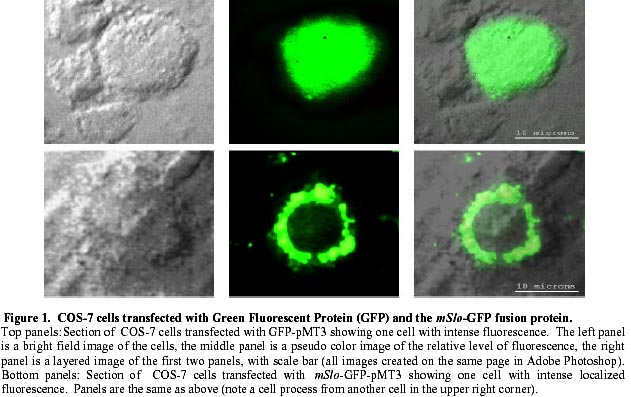
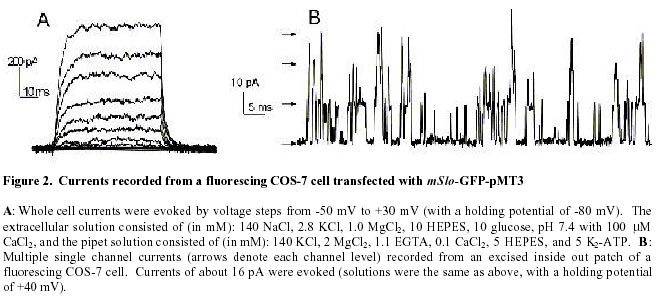
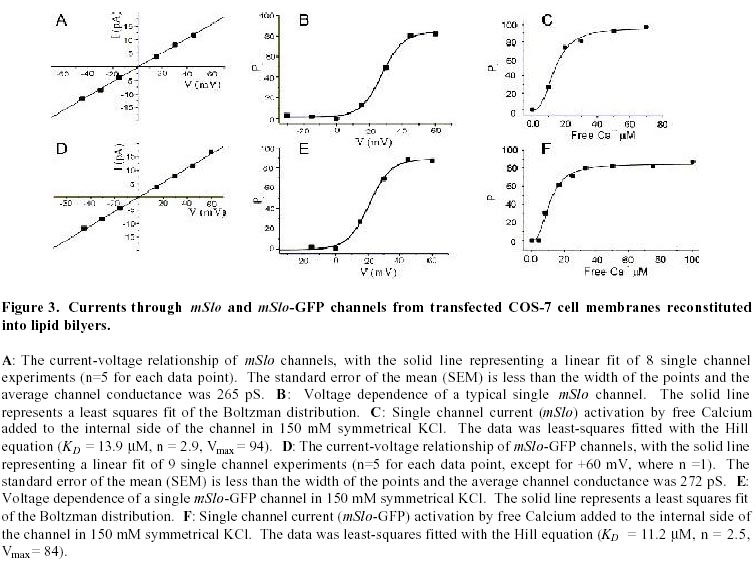
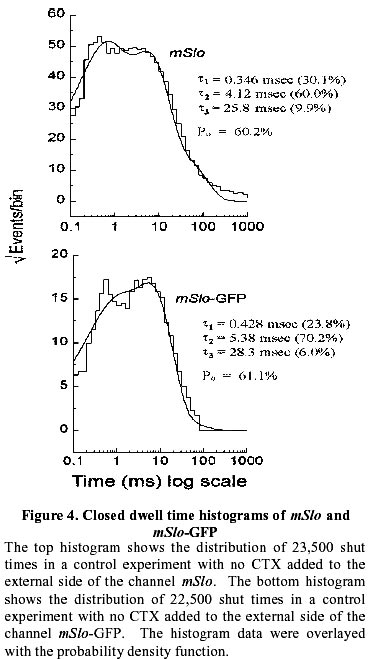
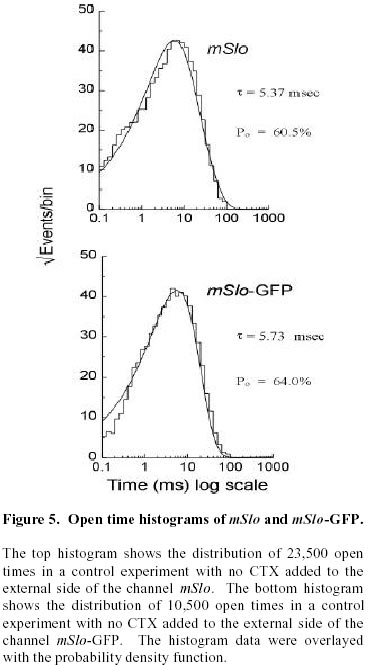
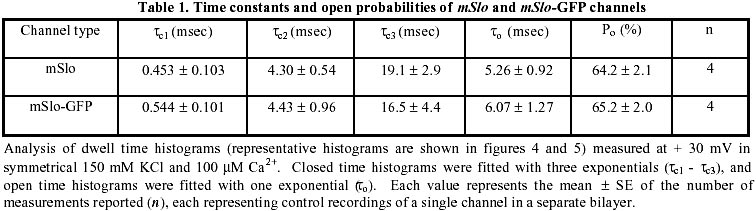
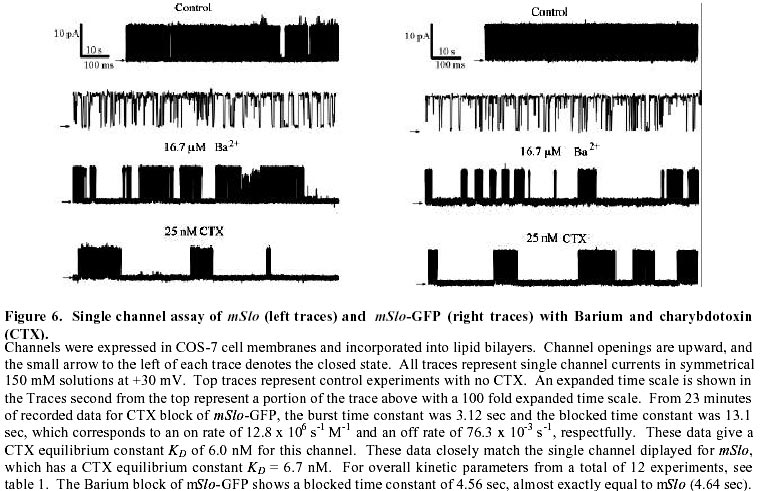
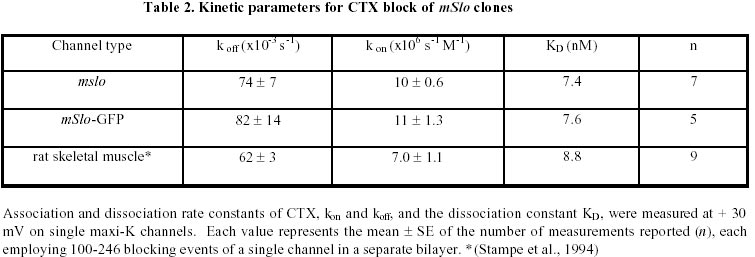
August 20, 1999 Code Number: ej99016 We have constructed a fusion protein between (mSlo) (a recombinant, high conductance, calcium-activated potassium channel or Maxi-K), and GFP (green flourescent protein). This construct represents a tag to not only monitor channel expression, but to locate the protein in living cells. The GFP was fused in frame to the carboxy-terminus of the (mSlo) core protein (mSlo-GFP fusion protein). Expression of this fusion protein in COS-7 cells resulted in robust fluorescence localized near the cell membrane. Fluorescing cells that were patch clamped exhibited whole cell currents with a direction consistent with potassium currents. Conversely, non-fluorescing cells showed no significant whole cell currents. Excised inside out patches revealed single channel currents and calculated conductances in the range of those expected for the maxi-K. The mSlo and mSlo-GFP channels reconstituted into lipid bilayers bound wild-type, recombinant CTX with high affinity and displayed a half-blocking concentration (KD) of 7.4 and 7.6 nM, respectively (at +30 mV in 150 mM equimolar KCl). This resulted in single channel evaluation of the functional inhibition of CTX on these clones. As newly constructed GFP chimeras emerge for the study of physiological processes in living organisms, this work provides another area of insight illuminated by GFP. Keywords : Barium block , Calcium activated potassium channel , Charybdotoxin , GPF fusion protein. Green Fluorescent Protein (GFP), cloned from the jellyfish Aequorea victoria is a powerful tool for biotechnology in studying the expression of various proteins. GFP is easily visualized when excited with UV light and its fluorescence does not depend on any exogenous compounds (Marshall et al., 1995). This protein, when expressed alone, appears cytosolic, with no cellular targeting mechanisms of its own, making it ideal to tag a protein of interest. Our fusion construct, called mSlo-GFP (for a review of GFP nomenclature see Gerdes and Kaether, (1996)), has the green fluorescent protein fused in frame to the c-terminus of the channel. The result is a functional tagged potassium channel. Our results demonstrate that the fusion protein can be expressed in COS-7 cells, harvested, and reconstituted into lipid bilayers allowing detailed single channel analysis. In addition, we show that the fusion protein can be studied by patch clamp techniques as well. Maxi-K channels belong to a class of calcium sensitive potassium channels which possess a large single channel conductance (greater than ~ 200 pS in symmetrical 150 mM KCl), thus they have been called BK (big potassium) or maxi-K channels (Latorre et al., 1989). These high conductance channels have been measured in numerous tissues and are associated with various physiological processes, including the repolarization of action potentials in neurons, regulation of fluid secretion in exocrine cells, and maintenance of contractile tone in smooth muscle (McManus, 1991). Recent physiological work has linked maxi-K channels to the mediation of vasorelaxant effects of K+ channel openers in the porcine coronary artery (Balwierczak et al., 1995), to the regulation of airway cholinergic neurotransmission (Tagaya et al., 1995), to the control of the dynamics of aqueous humor outflow in the anterior of the human eye (Stumpff et al., 1997), and to second messenger systems contributing to the membrane conductance of retinal Müller cells (Bringmann et al., 1997). These large conductance channels have also been implicated in several diseases, including the pathogenesis of hypertension in spontaneously hypertensive rats (Asano et al., 1995), the molecular mechanism of antiasthma therapy (Barnes, 1995), and to the inherited susceptibility to non-insulin-dependent diabetes mellitus (NIDDM) (Ferrer et al., 1996). While a great deal of information has been accumulated from studying native maxi-K channels, the channel was difficult to clone and then to express. This has barred it from the explosion of data provided by molecular biology that has increased our knowledge of other potassium channels. Maxi-K channels have been cloned and expressed from the slowpoke locus (Adelman et al., 1992). This began the molecular characterization of the channel. The first maxi-K mammalian clone from the mouse brain (mSlo) appeared in 1993 (Butler et al., 1993). Human analogues (hSlo) of this channel have been cloned from a variety of sources including the brain (Dworetzky et al., 1994; Tseng-Crank et al., 1994), arterial smooth muscle (McCobb et al., 1995), myometrium (Wallner et al., 1995), and pancreatic islet cells (Ferrer et al., 1996). While structurally homologous to the purely voltage-gated potassium channels such as Shaker (both are characterized by six n-terminus hydrophobic membrane spanning domains and an additional pore domain), the cloned DNA for maxi-K channels is more than double the size, containing a large c-terminus coding region believed to confer their calcium sensitivity (Knaus et al., 1995; McCobb et al., 1995; Adelman et al., 1992). This increased size may be one reason that causes the difficulty of channel expression. Cloned maxi-K channels appear to express well in oocytes, yielding robust macropatch cell currents, but single channel analysis necessary for detailed kinetic analysis of CTX block has not been forthcoming, especially in the mammalian clones (Butler et al., 1993; McCobb et al., 1995). Furthermore, studies measuring the affinity of CTX to BK channels reconstituted into artificial bilayers such as the one described here, have an advantage in that oocytes as an alternative expression system are now known to be contaminated with native BK channels (Krause et al., 1996). The initial characterization of the pharmacology of cloned maxi-K channels was completed only recently (Gribkoff et al., 1996), and our work adds to the analysis of the mSlo clone, particularly with regard to its interaction with CTX. CTX has been very useful in studies of ion channel structure and function. Its interaction with specific residues in the pore region of ion channels makes it an ideal probe when relating specific amino acids to their physiological role. Studies show CTX to be 37 amino acids in length, 20 x 20 x 25 Angstroms in dimension, with a globular structure formed by a three-turn alpha helix lying on an antiparallel b sheet with three internal disulfide bonds (Bontems et al., 1991). Use of CTX has not only identified residues in contact with the toxin, it marks residues in proximity to the receptor for CTX (Goldstein et al., 1994). When CTX is applied to preparations of native maxi-K channels, it blocks the channel in a bimolecular fashion, binding to the external vestibule of the channel and physically occluding or "plugging" the outer entry to the conduction pore (Miller, 1995). Moreover, studies of CTX binding have linked specific residues on the toxin to permeation sites in the pore, enriching our molecular picture of the toxin and the nature of its receptor in potassium channels (Naini et al., 1996). Recently, CTX and its close scorpion toxin analogue, agitoxin2, have been used to show that prokaryotic K+ channels have the same pore structure as eukaryotic K+ channels (MacKinnon et al., 1998). This result is important, given that the crystal structure of a prokaryotic K+ channel from Streptomyces lividans has been revealed by Doyle et al. (1998). The testing of this ground-breaking structure will require detailed toxin profiles, such as those presented here, especially if we are to extrapolate the structure of a two-transmembrane K+ channel to a six-transmembrane K+ channel, such as Shaker, or to an even more complex ten-transmembrane K+ channel, such as the maxi-K described in this report. Preliminary data of these findings have been reported in abstract form (Myers et al., 1997). Molecular biology. For the expression of GFP alone, Green Fluorescent Protein cDNA (from the pGFP clone, Clontech, Palo Alto, CA) was subcloned into expression vector pMT3 (from the laboratory of Daniel D. Oprian, Brandeis University). The pMT3 vector is pMT2 (Sambrook et al., 1989) with the Pst I and Eco RI sites mutated to Kpn I and Not I, respectively. To express GFP as a fusion protein, the GFP was subcloned into an expression vector of pMT3 containing the a subunit of mSlo (mbr5) (Butler et al., 1993). The stop codon in mSlo was mutated to a lysine residue (TTG), which allowed for the translation of nine additional bases (CTCCCAGGA) at the c-terminus of mSlo before the sequence of GFP began. As a result of the subcloning steps, fourteen base pairs were eliminated from the 3' untranslated sequence of mSlo (CTATTTTTTTAAAG). A mismatch mutagenesis strategy (Kammann et al., 1989) using PCR (polymerase chain reaction) was employed to produce the mutations of the stop codon and subcloning. DNA fragments were separated by agarose gel electrophoresis and purified using QIAQUICK gel extraction kit (Qiagen, Chatsworth, CA). Oligonucleotides used in this study were custom made by the Oligonucleotide Core Facility at the University of Rochester. To confirm the fusion construct, DNA sequencing was carried out at the Oligonucleotide Core Facility using the dideoxy chain termination method (Sanger et al., 1977). Whole cell and vesicle preparation. COS-7 cells (ATCC, Rockville, MD) were incubated in a CO2 incubator in 100 mM tissue culture plates at 37 °C until they reached 70-80% confluency. The cells were then transfected using a DEAE-Dextran cationic-lipid method (Oprian et al., 1987) using 2 mg of DNA per plate. Cells were assayed for channel activity by patch clamp or harvested into vesicles for reconstitution into lipid bilayers 48 to 72 hours after transfection. For lipid bilayer preparation, the cells were scraped off the culture plates and collected into ice cold 10 mM sodium phosphate buffer, pH 7.0, containing 150 mM NaCl. The cells were resuspended in a high pH solution (150 mM KCl, 2mM MgCl2, 5 mM EGTA, pH 10.6 with ammonium hydroxide) and broken up by being taken up in a 20 ml syringe 4 times through a high gauge needle (2 times with a 22 gauge, then 2 times with a 25 gauge). The broken cells were sonicated 30 secs to form lipid vesicles (Branson model 450 sonicator, duty cycle 50%, output 4). The vesicles were purified on a step sucrose gradient (40% and 20% sucrose cushion, interface between these layers contains the vesicles). The purified vesicles containing the channels were resuspended in isotonic sucrose buffer (10 mM MOPS and 250 mM sucrose, pH 7.4), quick frozen on dry ice, and stored at -80°C. Electrophysiology. For single channel analysis, the vesicles containing the recombinant channels were reconstituted into lipid bilayers and characterized. The recording chamber was made out of acetyl copolymer (Patriot Plastics, Londonderry, NH). It includes two open wells (one with a glass side called the cis chamber and another behind it called the trans chamber) separated by a thin wall with a 250 mm hole drilled through it. A forming solution of 100 ml of N2 dried lipid (1-palmitoyl, 2-oleoyl-phosphotidylethanolamine/1-palmitoyl, 2-oleoyl-phosphati- dylcholine, 10 mg/ml, (Avanti Polar Lipid, Alabaster, Al), 7:3 respectively and 50 ml n-decane (Sigma) was used to prepaint the chamber. The cis chamber contained 650 mM KCl, 10 mM HEPES, 100 mM Ca2+, pH 7.4. The trans chamber contained an identical solution, except its KCl concentration was 50 mM. Vesicles from the COS cell prep were added to the cis chamber and after incorporation of a single maxi-K channel, the cis side of the chamber was perfused with 150 mM KCl and KCl was added to the trans chamber resulting in an isotonic KCl concentration of 150 mM for all recordings. To measure calcium activation and sensitivity to block, stock solutions of CaCl2, BaCl2, and recombinant CTX were pipetted into the appropriate chamber when needed. The CTX used was purified to its active form after cleavage from a fusion protein in E. coli (Park et al., 1991). The trans chamber was connected to ground and all the voltages in the cis chamber are expressed relative to ground. The CTX experiments were all carried out at + 30 mV. For patch clamp experiments, whole cell currents were evoked by voltage steps from -50 mV to +30 mV (with a holding potential of -80 mV), and single channel currents evoked in excised inside out patches at a holding potential of +40 mV. The extracellular solution consisted of (in mM): 140 NaCl, 2.8 KCl, 1.0 MgCl2, 10 HEPES, 10 glucose, pH 7.4 with 100mM CaCl2, and the pipet solution consisted of (in mM): 140 KCl, 2 MgCl2, 1.1 EGTA, 0.1 CaCl2, 5 HEPES, and 5 K2-ATP. Data acquisition and analysis. The cis and trans chambers were connected with electrodes to an Axopatch 200A patch clamp amplifier (Axon Instruments, Foster City, CA) in capacitive feedback configuration. The lowpass cutoff filtering frequency was set at 1 kHz. The analog output signal was digitized by a Data Acquisition Processor (DAP) 3200e, (Microstar Laboratories) at 100 kHz. Open and closed times were computed by an on-board processor (486DX) on-line at 100 kHz with a threshold set at 50% of the whole open-level current. At the same time, single-channel currents were sampled at 1 kHz. Cumulative open probability (Po) was computed from the total open and closed times. During the recording, Po vs. number of transitions was displayed after every 1,000 transitions. Patch Clamp data were also obtained using an Axopatch 200 amplifier (5 kHz low pass filter) and currents were recorded using pCLAMP 5.2 software. Rate constants for association and dissociation of CTX and Ba2+ were calculated from the statistical distributions of the blocked and unblocked dwell times, as previously described (Anderson et al., 1988). A pentium/150 MHz Personal Computer and a custom written program were employed to control the experiment, and to store and analyze the data. Detection of expressed fluorescence. The cells were imaged for analysis in one of two ways: 1) To obtain higher resolution in imaging the fusion construct, the cells were fixed in 4% paraformaldehyde and washed in 0.05 M phosphate buffer. The cells were air dried and mounted for observation on a Nikon microphot fluorescence microscope (figure 1). All cells were imaged using a Hoffman Modulated contrast objective. The cells were excited with UV light (mercury lamp), and observed in the green wavelength (with a FITC filter cube). 2) Similar results were obtained using confocal microscopy (data not shown) at the Adherent Cell Analysis and Sorting (ACAS) Facility of the University of Rochester. They were trypsinized and transferred to quartz plates for imaging. The cells were excited at a wavelength of 395 nm and their relative fluorescence was recorded at 488 nm. ResultsVisualization of GFP and expressed channels To find the optimal conditions for transfection, the fusion protein was used to transfect COS-7 cells under differing conditions of cell density, concentrations of DNA to lipid reagent, and transfection times. When GFP was expressed alone in our cloning vehicle (plasmid vector GFP-pMT3), conventional fluorescent microscopy revealed cells labeled with intense fluorescence throughout the cell (figure 1, top panels). When GFP was fused to the maxi-K channel (plasmid vector mSlo-GFP-pMT3), the same microscopy revealed robust fluorescence localized near the cell membrane (figure 1, bottom panels). After transfection with mSlo-GFP-pMT3, fluorescent cells that were patch-clamped (4 out of 5), displayed typical outward going voltage gated maxi-K currents (figure 2, A and B). COS-7 cells that were not fluorescing showed no currents (4 out of 4). Characterization of channels reconstituted into lipid bilayers Currents through mSlo channels reconstituted into lipid bilayers had an average conductance of 265 pS and were voltage dependent (figure 3, A and B). The opening of the tagged channel was also voltage dependent with an average channel conductance of 272 pS (figure 3, D and E). mSlo was calcium sensitive in the mM range (figure 3C), which closely matches the calcium sensitivity of mSlo-GFP (figure 3F). The calcium activation data was least-squares fitted with the Hill equation (KD = 13.9 mM, n = 2.9, Vmax = 94, and KD = 11.2 mM, n = 2.5, Vmax = 84 for mSlo and mSlo-GFP, respectively). Although only one distribution of the voltage dependence is displayed for typical mSlo and mSlo-GFP channels, both the calcium and voltage sensitivity varied from channel to channel. Wu et al. (1996), describe the calcium variation as a clustered distribution of calcium sensitivities in maxi-K channels from avian nasal gland cells. To determine any differences in voltage or calcium sensitivity, the open probability of several single channel experiments under identical conditions of voltage and free calcium concentration were averaged together. The mean open probabilities of mSlo and mSlo-GFP channels (63.9 ± 4.5% and 52.8 ± 12%, n = 10 and 9, respectively) were not significantly different from each other at +30 mV and 100 mM Ca2+. Thus, the addition of green fluorescent protein to the mSlo channel had very little, if any, effect on its functional characteristics described above. Statistical distribution of mSlo and mSlo-GFP dwell timesTo further test if the gating of the channel was unchanged, the statistical distributions of the shut and open times of control records were analyzed using the probability density function (representative distributions of shut and open dwell times are shown in figures 4 and 5. respectively). Rate constants from 4 channels for each clone with similar open probabilities were averaged together (see table 1). We were able to resolve 3 exponents to describe the distribution of the shut times of the channels. The average shut time rate constants ± SEM, were: t c1 = 0.453 msec ± 0.103, t c2 = 4.30 msec ± 0.540, t c3 = 19.1 msec ± 2.95 for mSlo, and t c1 = 0.544 msec ± 0.101, t c2 = 4.43 msec ± 0.962, t c3 = 16.5 msec ± 2.95 for mSlo-GFP, respectively (see table 1). The distribution of the open dwell times were well described by a single exponent, 5.26 ± 0.923 msec for mSlo, and 6.07 ± 1.27 msec for mSlo-GFP (see table 1). These values are consistant with those recently reported for wild type mSlo (Sullivan et al., 1997). The rate constants describing the open and shut times of both channels were nearly identical for the wild type mSlo and mSlo-GFP fusion channels. Thus adding GFP to the channel does not appear to have any significant effect on the gating of the channel. CTX block of mSlo and mSlo-GFP Structural features of the mSlo-GFP fusion channel, namely the outer vestibule, were explored using CTX and Barium, respectively, as probes. CTX block of typical single channels (figure 6) revealed similar functioning of the two channels. The KD for CTX block for the fused channel was 7.5 nM compared to 7.3 nM for mSlo (see table 2). Ba2+ was also used to probe the channels in a single experiment on each clone (see figure 6). The Barium block of mSlo-GFP showed a blocked time constant of 4.56 sec, compared to a blocked time constant of 4.64 sec for mSlo. Concentration dependence of CTX block of mSlo To investigate the concentration dependence of CTX block of mSlo, the channel was assayed with the toxin at several different toxin concentrations (see figure 7). As seen with native maxi-K channels, the interaction of CTX with mSlo was bimolecular. The blocked dwell times were exponentially distributed and the average blocked time, t b, was independent of toxin concentration (figure 7, open circles). Increasing the concentration of CTX has no effect on the off rate. The burst times were also exponentially distributed and the on rate increased with increased toxin concentration (figure 7, closed circles). As the concentration of CTX was increased, the on rate increased linearly.
We have used GFP to estimate expression of maxi-K channels prior to reconstitution into lipid bilayers and to locate transfected cells for patch clamping. The fusion protein has allowed us to monitor transfection conditions necessary to express the a subunit of mSlo. Given that single channel recordings of mSlo are rare (Muller et al., 1996) yet essential for a detailed evaluation of CTX block including its interaction with the b subunit (Strobaek et al., 1996), a functional reporter gene to monitor expression in various expression systems and conditions is a powerful tool. In this study, we have demonstrated that the mSlo channel expression and protein targeting, kinetics, and functional properties of the outer vestibule are all unchanged by GFP fusion at the carboxy-terminus. Our halo like location of the protein (figure 1, bottom panels) has also been seen in a preliminary report of another tagged potassium channel (John et al., 1997), and suggests that the channels are inserted into the plasma membrane. Therefore, sub-cellular localization of ion channels now appears possible. As expression of various constructs are undertaken to elucidate ion channel structure and function, it has been suggested that tagging of these clones will help us to distinguish between non-functional channels and those whose biogenesis was arrested (Makhina et al., 1997). Thus this new construct represents a tag to not only monitor channel expression, but to locate the protein in living cells. A preparation of transfected cells can now be checked for efficiency before they are reconstituted simply by evaluating the fluorescence of the cells. The fluorescence observed is quite stable, and we demonstrate here that cells with no fluorescence do not express channels. The patch clamp and lipid bilayer results, as well as the fluorescent images suggest that the GFP is indeed fused to the channel and that the channel is fully functional. Moreover, a functional delayed rectifier K+ channel clone (RCK1) has recently been fused to GFP and characterized (Trouet et al., 1997), further demonstrating that GFP can mark cells while not affecting channel behavior. Our analysis of the time constants and distributions for the mSlo and mSlo-GFP fusion channels revealed that the normal gating of the channel was apparently unaffected by addition of GFP to the c-terminus. Our time constants are consistent with those previously reported for mSlo. Sullivan et al. (1997), describe the open times with one exponent (t o = 58 msec), and were able to resolve 4 exponents to describe the shut times of mSlo: t c1 = 0.59 msec, t c2 = 2.5 msec, t c3 = 9.4 msec, and t c4 = 238 msec at +20 mV in 200 mM Ca2+. Their experimental conditions (designed to analyze calcium sensitivity) were very different from ours, making a comparison difficult. While attaching GFP to the carboxy-terminus of mSlo had no effect on channel function, results from an amino-terminus attachment to hSlo were quite different. Tagging the amino-terminus of hSlo with GFP apparently alters its calcium sensitivity (Meyer and Fromherz, 1999). No such affect on calcium sensitivity seen in our carboxy-terminus construct could be a clue that only the amino-terminus of the protein is involved in sensing calcium. A more detailed examination of the calcium sensitivity of our tagged protein will need to be done before a final conclusion can be made. The amino-terminus tagged hSlo result also suggests that care should be taken before extrapolating the results presented here to other channels. Another result of this study has been a single channel look at the functional inhibition of CTX on the a subunit of mSlo. Our KD value of 7.4 nM for mSlo and 7.6 nM for mSlo-GFP are on the order of the inhibition constant recently reported for the other CTX sensitive BK clone hSlo (Gribkoff et al., 1996). The EC50 for CTX of hSlo in the Gribkoff et al. study (estimated from inhibition of two-electrode voltage clamped current from oocytes) is 30.8 nM and 7.9 nM for Iberiotoxin (IbTX), a close structural homologue of CTX. The EC50 of IbTX (also estimated from inhibition of two-electrode voltage clamped current from oocytes) in mSlo was estimated to be 9.1 nM in the Gribkoff et al. (1996) study. In addition, we demonstrate here that CTX block of mSlo is bimolecular, showing that the clone follows the interaction of CTX with native maxi-K channels (Miller et al., 1985). The Gribkoff et al. (1996) study did not assess the concentration dependence of CTX block on mSlo. They did find a bimolecular interaction for IbTX with both hSlo and mSlo channels which corresponds to our finding. Their result of the interaction of CTX with hSlo was less definitive. A functional inhibition of CTX on hSlo single channel current may reveal a more detailed answer. The green fluorescent protein (GFP) from the jellyfish Aequorea victoria has become an important reporter molecule for monitoring gene expression. With this work, an RCK1 chimera (Trouet et al., 1997), and preliminary reports of other potassium channel GFP fusion proteins (John et al., 1997; Makhina et al., 1997; Veyna-Burke et al., 1997), as well as calcium channel GFP chimeras (Grabner et al., 1997; Zhou et al., 1997), a new list of GFP constructs will have to be added to the existing categories (Gerdes and Kaether, 1996), namely ion channels. In light of the fact that Xenopus laevis oocytes contain endogenous large conductance Ca2+-activated K+ channels, mSlo-GFP could be utilized as one of several precautions taken to reduce the risk of contaminating recordings with native BK channels. For a list of suggestions, see Krause et al., (1996). Since we have demonstrated here that only fluorescent COS-7 cells express the clone, patch clamping only fluorescent oocytes should increase the chances of recording from BK clones and not endogenous maxi-K channels. Although we have not expressed and imaged mSlo-GFP in Xenopus laevis oocytes in this study, a recent report of another GFP fused K+ channel demonstrated that the fluorescent protein (constructed with the same fluorescent tag from Clontech, Palo Alto, CA) displayed surface expression in oocyte membranes resulting in visible fluorescence and macroscopic currents (Chan et al., 1997). Just as mSlo-GFP has allowed us to asses the affinity of CTX for mSlo, currents through GFP tagged G protein inwardly rectifying K+ (GIRK) channels have recently identified specific regions of subunits involved in channel openings (Chan et al., 1997). Ion channels fused to GFP have limitless applications in studying the structure and function of these proteins. Our fluorescing protein represents a tagged maxi-K channel. The use of fluorescent mSlo-GFP will hopefully shed some light on the limited information we have about these critical proteins. The authors would like to thank Lawrence Salkoff for the use of mSlo, Peter Shrager for the use of the Nikon fluorescent microscope, and Ted Begenisich for help in improving the manuscript. A special thanks goes to Ian Vabnick for help in recording the fluorescent images and for the support of the members of the Papazian Lab at UCLA.
Supported by UNESCO / MIRCEN network © 1999 by Universidad Católica de Valparaíso -- Chile The following images related to this document are available:Photo images[ej99016f2.jpg] [ej99016f7.jpg] [ej99016f1.jpg] [ej99016f5.jpg] [ej99016t1.jpg] [ej99016f4.jpg] [ej99016f6.jpg] [ej99016f3.jpg] [ej99016t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}