
|
Entomotropica
Sociedad Venezolana de Entomologia
ISSN: 1317-5262
Vol. 16, Num. 3, 2001, pp. 149-155
|
Entomotropica antes/formerly Boletín de Entomología Venezolana ,
Vol. 16, No. 3, Diciembre ,2001, pp. 149-155
Biology and laboratory culturing of the
root-feeding flea beetle, Longitarsus columbicus columbicus
Harold, 1876 (Chrysomelidae: Alticinae): a potential natural enemy of Lantana
camara L. (Verbenaceae) in South Africa
Jan-Robert Baars
Plant Protection Research Institute, Agricultural Research Council,
Private Bag X134, Pretoria, 0001, South Africa. Current address: 49 Johnawood
Drive Ashbourne, Co. Meath, Ireland.
E-mail: baarsjr@hotmail.com
Recibido: 28-vii-2000
Aceptado: 06-viii-2001
Correcciones devueltas
por el autor: 29-xi-2001
Code Number: em01018
RESUMEN
Baars JR 2001. Biología y
cultivo en laboratorio del coquito pulga, Longitarsus columbicus
columbicus Harold, 1876 (Chrysomelidae: Alticinae): un enemigo potencial
natural de Lantana camara L. (Verbenaceae) en Sur Africa. Entomotropica
16(3):149-155.
La planta ornamental introducida, Lantana camara L.
(Verbenaceae), es una de las peores malezas invasivas de Sur Africa. Esta
especie ha sido objecto de programmas de biocontrol en las últimas cuatro
décadas. Aunque se han reconocido algunos enemigos naturales, el nivel de
control es considerado insatisfactorio y un número de biocontroladores
potenciales están siendo evaluados. El coquito pulga Longitarsus columbicus
columbicus Harold ha sido considerado como altamente destructivo, atacando
las raíces de la Lantana, un nicho en el pasado ignorado en gran medida
en los estudios de biocontroladores. Es poco lo que se sabe acerca de este
potencial enemigo natural y los atributos de su historia de vida y modo de
acción son discutidas. Los adultos se alimentan de hojas y depositan sus huevos
en las hojas bajas, cercanas a la superficie del suelo. Las larvas penetran
dentro del suelo, donde se alimentan externamente de las raíces secundarias. El
tiempo de desarrollo toma alrededor de 60 días durante el verano con el
potencial de 2 a 3 generaciones por año en el campo de Sur Africa. Se describen
técnicas de cría, siendo notable el uso de una jaula modificada y se comparan
los resultados con un medio alternativo de cultivo en cápsulas de petri. Se
discuten las implicaciones de la especifidad con el hospedero de L.
columbicus columbicus, de otras especies de Longitarsus y del coquito
pulga en condiciones de laboratorio.
Palabras clave adicionales: Cariaquito,
comedores de raíces, control biológico clásico, cría en
laboratorio.
ABSTRACT
Baars JR 2001. Biology and
laboratory culturing of the root-feeding flea beetle, Longitarsus columbicus
columbicus Harold, 1876 (Chrysomelidae: Alticinae): a potential natural
enemy of Lantana camara L. (Verbenaceae) in South Africa. Entomotropica
16(3):149-155. The introduced ornamental plant, Lantana camara L.
(Verbenaceae), is one of South Africa’s worst invasive weeds. It has been the
target of a biological control programme here for the past four decades.
Although several natural enemies have been established, the level of control is
considered unsatisfactory, and a number of new potential biocontrol agents are
being evaluated. The flea beetle Longitarsus columbicus columbicus Harold
is considered to be highly destructive, attacking the roots of lantana, a niche
largely ignored by biocontrollers in the past. Little is known about this
potential natural enemy, and attributes of its life history and mode of feeding
are discussed. Adults feed on the leaves and deposit eggs in the leaf litter
near the soil surface. The larvae burrow into the soil, where they feed
externally on the secondary rootlets. Development takes about 60 days during
summer, with the potential of 2-3 generations per annum in the field in South
Africa. Rearing techniques are described, notably the use of a modified cage,
and the results are compared with the alternative of rearing cultures in petri
dishes. Implications for the host-specificity screening of L. columbicus
columbicus and other Longitarsus and root-feeding flea beetle species
under laboratory conditions are discussed.
Additional key words: External root-feeder, laboratory culture, classical
biological control.
INTRODUCTION
Several natural enemies have been imported, evaluated and released
as biological control agents against Lantana camara L. in South Africa
(Baars and Neser 1999), and other countries worldwide (Julien and Griffiths
1998). The majority of the agents released are leaf- and flower-feeding and
stem-attacking species. With the exception of two cerambycid beetles,
Plagiohammis spinipennis (Thomson) which bores the stem and root crown,
and Paravander xanthomelas (Guérin-Méneville) which bores into the stem
and large roots, no other root-feeding natural enemies have been considered for
release on L. camara worldwide. Root-feeding flea beetles, in particular
species of Longitarsus, may be able to supplement damage inflicted by the
suite of agents already established on lantana.
Field surveys in the countries of origin of the Lantana
complex have revealed several flea beetle species associated with the plants
(Winder and Harley 1983; Palmer and Pullen 1995). The intensity of adult feeding
damage noted in the field during exploratory surveys in USA (Florida), Mexico,
Jamaica, Trinidad and Venezuela, has encouraged interest in the potential of
various root-feeding flea beetles, including species in the genus
Longitarsus, as biocontrol agents.
Although a few Longitarsus species are considered to be
polyphagous, most are oligophagous and several have been used as biocontrol
agents. Examples include L. jacobeae (Waterhouse) and L.
flavicornis (Stephens) on Senecio jacobeae L. (Compositae) (Frick
1970; Frick and Johnson 1973), L. albinaeus (Foudras) on Heliotropium
europaeum L. (Boraginaceae) (Delfosse and Cullen 1981; Huber 1981) and L.
aeneus Kutschera and L. echii Koch on Echium plantagineum L.
(Boraginaceae) (Wapshere 1982). Other Longitarsus species are under
consideration as biocontrol agents, including L. horni Jacoby on
Chromolaena odorata (Zachariades et al. 1999), and L.
quadriguttatus Pont. on Cynoglossum officinale L. (Boraginaceae)
(Jordon 1997). In addition, the host records of other species, such as L.
columbicus columbicus Harold that has only been recorded from Lantana
spp. in Venezuela (Bechyné 1997), indicate the potential of these species as
biocontrol agents.
Little is known about the life history and host-range of L.
columbicus columbicus, besides its association with the Lantana
complex. This paper investigates the biology and laboratory culturing of
this flea beetle, as the first step towards evaluating its potential for release
as a natural enemy against L. camara in South Africa.
MATERIALS AND METHODS
A laboratory culture was started with some 55 adults that were
collected north of Maracay, along the road to Choroni (lat ° 34’45"N, long °
34’67"W) in Venezuela, and imported into South Africa in October 1998. Field
collections were made with a cupped beating tray, during the late afternoon.
Identifications were confirmed by Prof. Vilma Savini, and voucher specimens of
the beetles are deposited at the Museo del Instituto de Zología Agrícola (MIZA),
Maracay, Venezuela. Longitarsus columbicus columbicus belongs to the
tribe Longitarsini, and has also been collected from five other sites in
Venezuela and one in Colombia (Bechyné 1997). The closely related L.
columbicus centroamericanus Bechyné has been collected in Guatemala, El
Salvador and Nicaragua (Bechyné 1997).
During culturing in quarantine (Pretoria, South Africa)
field-collected adults were split into two groups. Twenty adults were exposed to
shoot tip cuttings in 15 cm petri dishes, and the remaining 35 adults were
exposed to whole plants placed in rearing cages (Figure
1). Each cage comprises a mild steel frame with psylla screen gauze
(Climatex cc, South Africa) and is fitted over a plastic trough with drainage.
In these cages, three small rooted lantana plants were transplanted into the
plastic troughs 2 weeks prior to exposure to promote new root growth. Adults
were removed after two weeks and placed into additional cages with fresh plants.
Cages were maintained under controlled glasshouse conditions and watered (from
above) so as to maintain fairly damp soil conditions. Temperatures ranged from
30°C to 20°C in a day/night cycle, under a natural summer photoperiod of about
14 hours. Emerging first generation adults were recorded and collected weekly,
and exposed to similar rearing conditions.
Petri dish rearing trials were conducted under laboratory
conditions, in a controlled temperature room at temperatures ranging from 29°C
to 22°C in a day/night cycle. Experiments were conducted on laboratory benches
under artificial light banks on a 13 hour photophase. Adults were exposed to
fresh cuttings every two days. The number of eggs, size of egg clusters,
oviposition site, and mode of feeding was recorded. These trials were repeated
using newly emerged adults from the first laboratory generation when the
preoviposition period was also measured. Eggs were maintained under moist
conditions and the duration of embryonic development recorded. Measurements were
made with the use of an ocular micrometer mounted on a Wild microscope.
Illustrations by the author were made with the use of a camera lucida.
Eight dominant L. camara varieties from the Mpumalanga,
Northern and KwaZulu-Natal Provinces of South Africa were used to rear the
beetles in both petri dishes and cages. In the cages, different varieties were
used to compare relative susceptibility for feeding and development, which was
subjectively rated by the amount of shot hole damage to the leaves, and the
number of progeny emerging from rearing cages respectively.
RESULTS
Biology
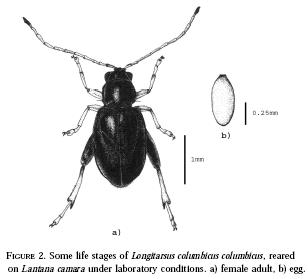
Adults of Longitarsus columbicus columbicus (Figure
2a) are small (about 2 mm in length) and have a dark
copper-brown colour. Adults feed on the leaves causing small shot holes, leaving
the upper epidermal layer intact. Field-collected adults survived for two months
under laboratory conditions, and remained fertile during this period. Adults
emerging from the laboratory cultures survived for up to three months, and
displayed a preoviposition period of about 2 weeks.
Adult feeding damage in the field was severe, and was similar to
damage observed for other unidentified species of flea beetles (including prob.
Longitarsus spp.) in Florida and Trinidad in April 1996, Mexico in
October 1998, and Jamaica in July 1999. Adults are active and jump readily, and
peak activity appears to occur during the cooler hours of the late afternoon, or
under cloudy conditions. Adults shelter in the lower parts of the lantana
plants, and amongst the surrounding vegetation during the hotter periods of the
day.
Eggs were deposited under moist filter paper and cotton wool in
petri dishes, but are normally deposited under moist leaf litter in cages. The
eggs are minute, measuring 0.58±0.01mm (mean ± SE) by 0.25±0.02x10-1mm (n=63)
(Figure 2b), and were deposited in groups averaging
about 3.2±0.5 eggs (n=26) in the petri dishes. Eggs took 12.0±0.2 days (n=18) to
hatch and the emerging larvae are highly active and mobile, but suffer high
mortality under dry soil conditions in the laboratory. First instar larvae enter
the soil in search of secondary rootlets on which they feed externally. After
exposure to a culture of L. columbicus columbicus plants were found to be
largely devoid of secondary rootlets. Late instar larvae pupate in a hardened
soil capsule near the soil surface. Newly emerged adults are active and highly
mobile.
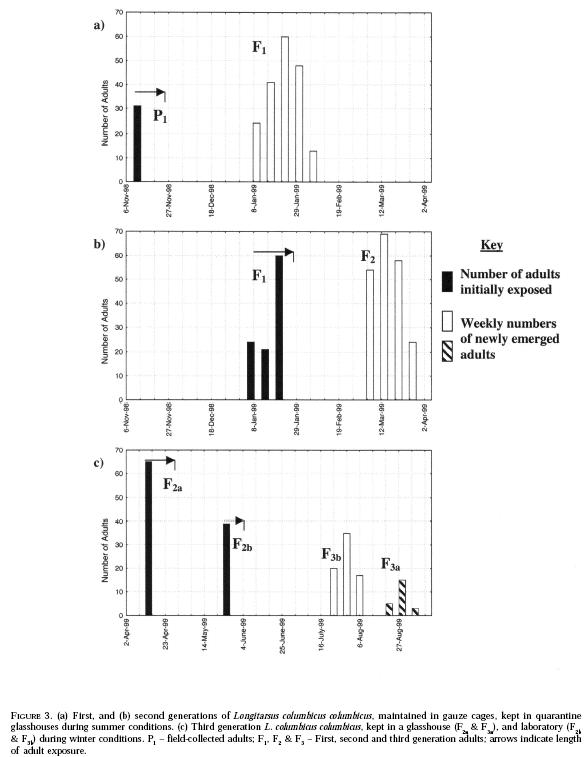
The entire life cycle (egg-laying to adult eclosion) took about 60
days in summer. The parent:progeny ratios of cultures in glasshouses, for the
first two generations (Figure 3a &
b) were high,
P:F1 (35:186) and F1:F2 (105:205). Concurrent
F1:F2 cultures showed similar ratios of 132:215, and
106:201. The third generation development (Figure
3c)
varied with the culturing conditions, with a ratio of 65:23
(F2a:F3a) exposed to shorter day lengths during the
natural winter conditions in a glasshouse, and 39:79
(F2a:F3a) exposed in a laboratory (as described in the
Materials and Methods). Under natural winter conditions adults emerged after
about 126 days (Figure 3c
F2a:F3a), whereas those under laboratory conditions
emerged after about 60 days (Figure
3c
F2a:F3a).
Rearing techniques
Adults reared on lantana shoot tips in petri dishes, fed and
oviposited readily but experienced high mortality of up to 100% within 2 weeks.
Adults were noted to jump readily when disturbed under these confined
conditions, notably during the renewal of shoots. Eggs deposited under the moist
filter paper were transferred to containers with moist cotton wool and the
emerging larvae were transferred to the stem bases of lantana plants grown in
15cm pots. However, these plants periodically dried out and wilted, and no adult
emergence occurred under these conditions. By contrast adults survived better
under the large cage conditions, with larger numbers of progeny emerging (Figure
3a, b & c). However, plants in cage troughs were
prone to invasion by ants, and in these instances larvae and pupae suffered
extensively from ant predation.
The intensity of adult feeding damage, and the number of progeny
emerging in the first and second generations were noted to be similar on the
eight lantana varieties that were exposed in both petri dish and cage
experiments. The varieties of L. camara used, including orange, pink and
white flowering forms, differed in morphological features like leaf hairiness
and toughness, factors which seemingly had no noticeable effect on rearing
success.
DISCUSSION
Previous attempts to culture an unidentified ?Longitarsus
species on lantana from Mexico, during the winter period, have proved
unsuccessful (Baars and Neser 1999). In this study, a culture of L.
columbicus columbicus was successfully manipulated to maintain egg laying
and immature development under artificial light and temperature regimes.
Cultures reared in cages as described in this paper, were maintained under
natural winter photo periods in glasshouses. The delayed emergence of the
F3 progeny of L. columbicus columbicus exposed to natural
winter conditions, suggest a significantly slow developmental rate or a state of
diapause was induced. An immature stage probably enters a state of oligopause,
as described by Mansingh (1971), which may be induced by a gradual change in
photoperiod and/or plant physiology. Presumably, field populations enter
a similar state to overcome the drier conditions and plant dormancy in
Venezuela.
Laboratory cultures of L. columbicus columbicus are best
maintained under the caged conditions described above. Under these conditions,
adults should be exposed to the plants for short periods (about 2 weeks) to
reduce the egg load per cage. Plants with larvae should be carefully watered to
maintain intermediate soil moisture levels, in order to promote secondary root
development and reduce larval mortality. Although laboratory rearing techniques
are largely successful, mass-collection of insects in the country of origin for
use in host-specificity tests may provide an efficient alternative. This method
has been used for other Longitarsus species (Wapshere 1982; Jordon 1997),
especially univoltine species, which take too long to culture.
The external mode of larval feeding appears to expose the immature
stages of L. columbicus columbicus to high rates of mortality during
excessive soil desiccation, and also probably excessive soil saturation. Areas
infested with L. camara in South Africa are generally subject to summer
rainfall and dry winters. During winters the topsoil dries excessively forcing
the lantana plants into a state of dormancy. As a result natural enemies
attacking this niche have to be adapted to cope with these extreme winter
conditions. For biocontrol agents to successfully suppress lantana in the drier
areas of South Africa, insect populations must survive the winter period in
sufficient numbers to maintain levels of damage on plants during the spring
regrowth, as was the case for L. albineus on H. europaeum (Huber
1981). By means of diapause the pupae of L. columbicus columbicus may
thus provide a resistant life stage, which is able to cope with the unfavourable
winter conditions. Wapshere (1983) discussed the effectiveness of biotypes of
L. jacobaeae, originating from different geographical regions in Europe,
as biocontrol agents in Australia, highlighting the importance of environmental
synchrony.
Adults of root-feeding Longitarsus spp. that have been
employed as biocontrol agents either feed externally, like L. aeneus on
Echium plantagineum (Wapshere 1982) and L. albineus on H.
europaeum (Delfosse and Cullen 1981; Huber 1981), or internally in the root
crown, like L. echii on Echium plantagineum (Wapshere 1982), and
L. jacobaeae on S. jacobaeae (Frick and Johnson 1973). Through
resource partitioning, Wapshere (1982) argued that the combined impact of the
two Longitarsus species on Echium plantagineum improved the
prospects for control. Similarly the biocontrol programme against L.
camara in South Africa may benefit if priority be given to determine whether
there are Longitarsus species, or other root-feeding flea beetles, which
feed internally to avoid dry soil conditions. Surveys for other root-feeding
agents have so far been conducted in Mexico and Jamaica, in 1998 and 1999
respectively (Baars unpubl.). The Mexican survey, conducted in the northern
coastal areas of the Yucatan, Tabasco and Veracruz provinces, resulted in the
collection of an unidentified flea beetle (Accession number: AcSN 2431, National
Collection of Insects, ARC- Plant Protection Research Institute, Pretoria, South
Africa) that has a similar life cycle to L. columbicus columbicus except
that the larvae feed internally in the cortex of the primary roots.
Lantana camara
in South Africa is a highly variable entity
with numerous varieties differing in morphology, physiology and genotype (Baars
and Neser 1999). Varietal resistance is deemed to have contributed to natural
enemies either failing to establish or performing poorly on certain varieties
(Cilliers and Neser 1991). It is thus important to expose natural enemies under
evaluation to the main varieties of lantana, which have invaded large areas
of
South Africa. Feeding intensity and progeny development of L. columbicus
columbicus appeared not to be influenced by the lantana varieties exposed,
indicating its potential to cope with this diverse weed.
CONCLUSIONS
Field and laboratory observations indicate that L. columbicus
columbicus is a potentially damaging natural enemy that warrants more
intensive host-specificity evaluation in South Africa. Longitarsus columbicus
columbicus and other root-feeding flea beetles constitute a specialized
guild that has not been represented in the biocontrol programme against
Lantana camara worldwide. The rearing methods discussed are able to
supply healthy laboratory cultures for host-specificity screening, but
alternative methods such as mass-collections in the country of origin may also
prove effective. The seasonal synchrony between L. camara and
Longitarsus species will be an important factor influencing establishment
and population increases in South Africa. With reference to the dry winter
conditions, further exploration for other internal root-feeding flea beetle
species remains a priority.
ACKNOWLEDGEMENTS
I thank my colleague Dr Costas Zachariades for accompanying me on
the survey in Venezuela. Thanks are due to Fritz Heystek for his continual
assistance during laboratory culturing. I also thank Beth Grobbelar of the
National Collection of Insects (ARC- Plant Protection Research Institute,
Pretoria, South Africa) and Prof. Vilma Savini (Universidad Central de
Venezuela, Maracay) for their identification support and services, respectively.
I would like to thank Profs Carlos Julio Rosales (Universidad Central de
Venezuela, Maracay), and Vilma Savini for their assistance during our visits,
and the Venezuelan Entomological Society for the opportunity to submit this
manuscript. I thank Drs Martin Hill, Terry Olckers and Costas Zachariades for
their very valuable comments on the manuscript. This research would not have
been possible without the financial support of the National Department of
Agriculture, Department of Water Affairs and Forestry and the Agricultural
Research Council of South Africa.
REFERENCES
- Baars JR, Neser S. 1999. Past and present initiatives on the
biological control of Lantana camara (Verbenaceae) in South Africa. Afr
Entomol Mem 1:21-33.
- Bechyné J. 1997. Evaluación de los datos sobre los Phytophaga
dañinos en Venezuela (Coleoptera) Parte I & II. V. Savini (Ed.). Bol Entomol
Venez, Ser Monogr 1.
- Cilliers CJ, Neser S. 1991. Biological control of Lantana
camara (Verbenaceae) in South Africa. Agric Ecosyst Environ 37:57-75.
- Delfosse ES, Cullen JM. 1981. New activities in biological control
of weeds in Australia. Common heliotrope, Heliotropium europaeum. Proc.
5th Int Symp Biol Contr Weeds 545-561.
- Frick KE. 1970. Longitarsus jacobaeae (Coleoptera:
Chrysomelidae), a flea beetle for the biological control of Tansy Ragwort. 1.
Host plant specificity studies. Ann Entomol Soc Am 63(1):284-296.
- Frick KE, Johnson GR. 1973. Longitarsus jacobaeae
(Coleoptera: Chrysomelidae), a flea beetle for the biological control of Tansy
Ragwort. 4. Life history and adult aestivation of an Italian biotype. Ann
Entomol Soc Am 66(2):358-367.
- Huber JT. 1981. Observations on the heliotrope flea beetle,
Longitarsus albineus [Col.: Chrysomelidae] with tests of its host
specificity. Entomophaga 26(3):265-273.
- Jordon T. 1997. Host specificity of Longitarsus
quadriguttatus (Pont., 1765) (Col., Chrysomelidae), an agent for the
biological control of hound’s-tongue (Cynoglossum officinale L.,
Boraginaceae) in North America. J App Entomol 121:457-464.
- Julien MH, Griffiths MW. 1998. Biological Control of Weeds: A
world catalogue of agents and their target weeds. 4th Edition. CABI Publishing,
UK.
- Mansingh A. 1971. Physiological classification of dormancies in
insects. Can Entomol 103:983-1009.
- Palmer WA, Pullen KR. 1995. The phytophagous arthropods associated
with Lantana camara, L. hirsuta, L. urticifolia and L.
urticoides (Verbenaceae) in North America. Biol Contr 5:54-72.
- Winder JA, Harley KLS. 1983. Phytophagous insects on lantana in
Brazil and their potential for biological control in Australia. Trop Pest
Manage 29:346-362.
- Wapshere AJ. 1982. Life histories and host specificities of the
Echium flea beetles Longitarsus echii and L. aeneus [Col.
Chrysomelidae]. Entomophaga 27(2):173-181.
- Wapshere AJ. 1983. Discovery and testing of a climatically adapted
strain of Longitarsus jacobaeae [Col.: Chrysomelidae] for Australia.
Entomophaga 28(1):27-32.
- Zachariades C, Strathie-Korrûbel LW. and Kluge RL. 1999. The South
African programme on the biological control of Chromolaena odorata (L.)
King & Robinson (Asteraceae) using insects. Afr Entomol Mem 1:
89-102.
© 2001- Sociedad Venezolana de Entomología
The following images related to this document are available:
Photo images
[em01018f2.jpg]
[em01018f3.jpg]
[em01018f1.jpg]
|

{kind=link}
{kind=link}
{kind=link}