 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Investigaciones recientes sobre Toxorhynchites rutilus (Diptera: Culicidae) con referencia al control biológico de mosquitos habitantes en recipientes Phil Lounibos1, Raul E Campos2 1Florida Medical Entomology Laboratory, University of Florida, 200 9th St
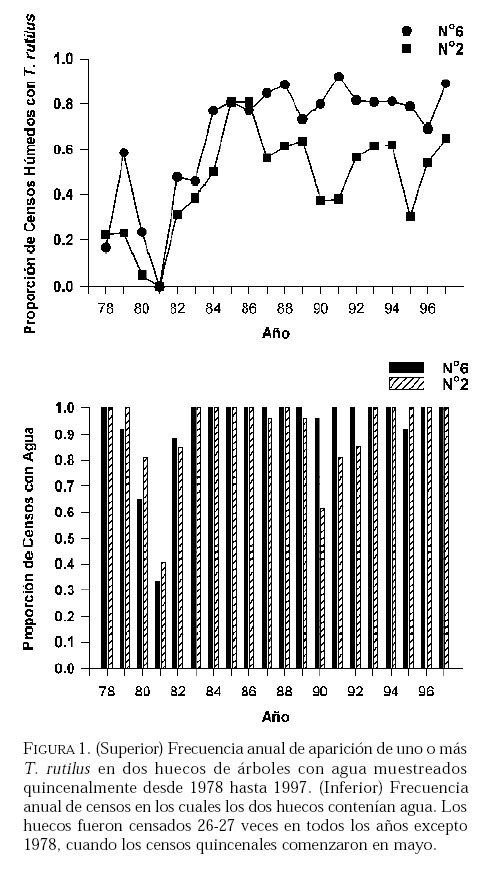
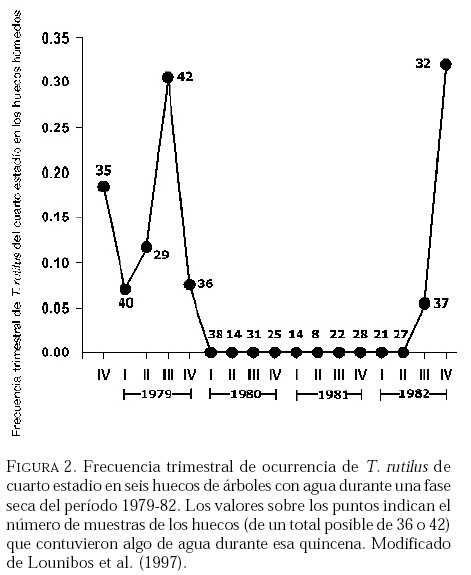
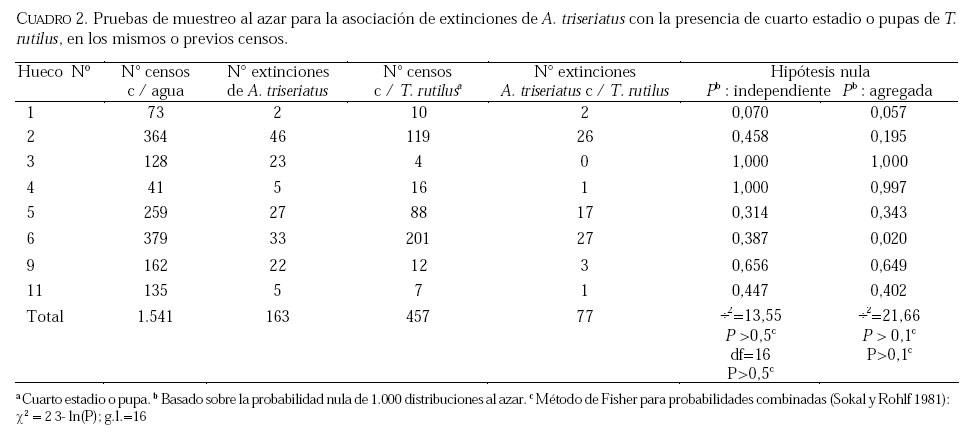
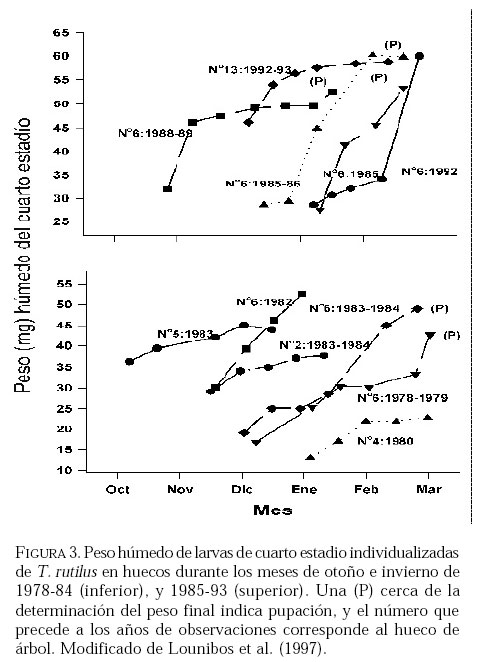
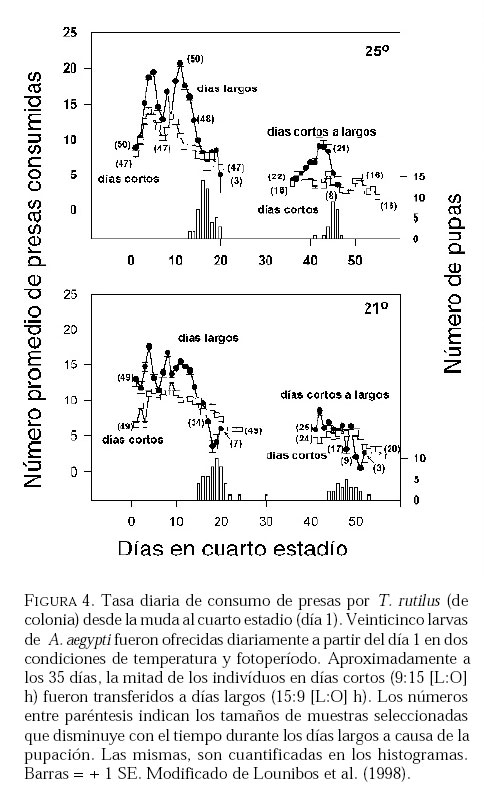
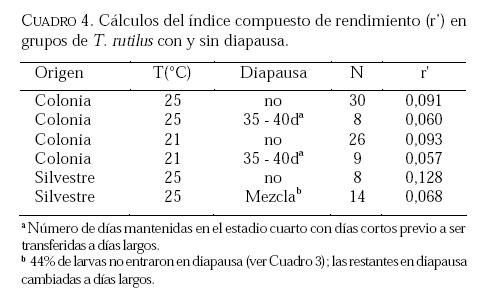
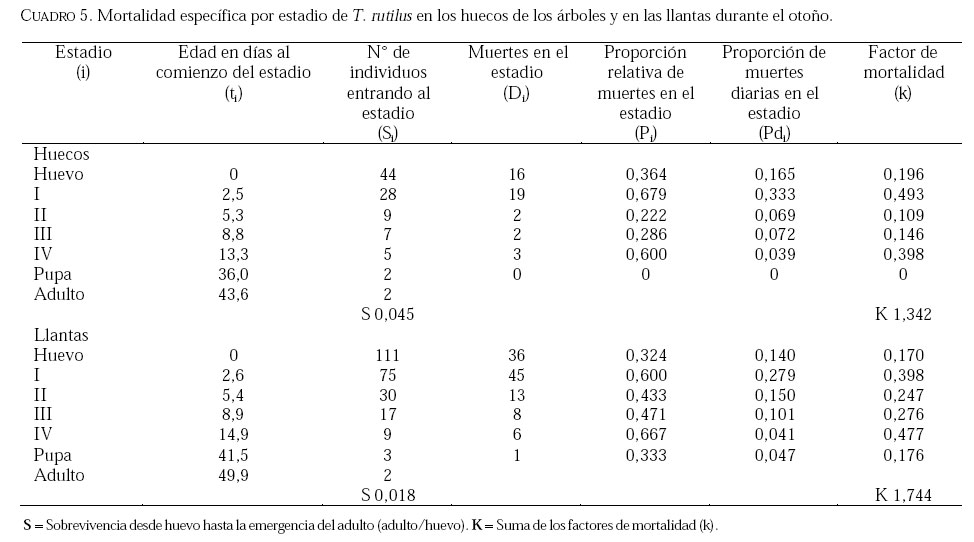
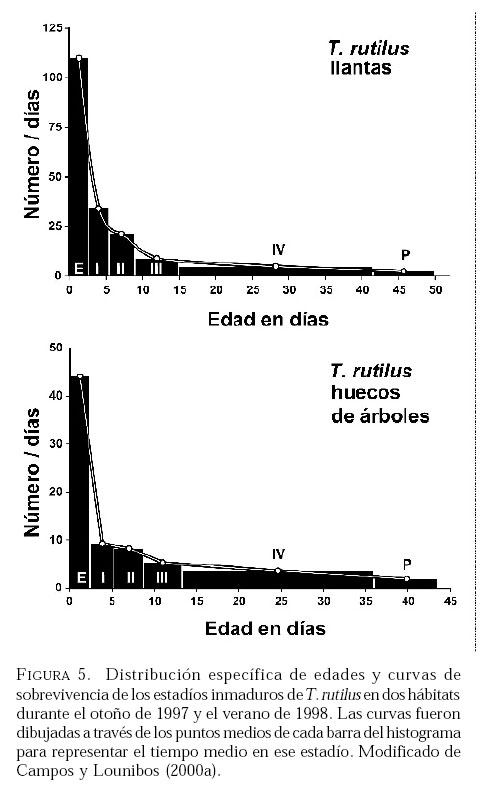
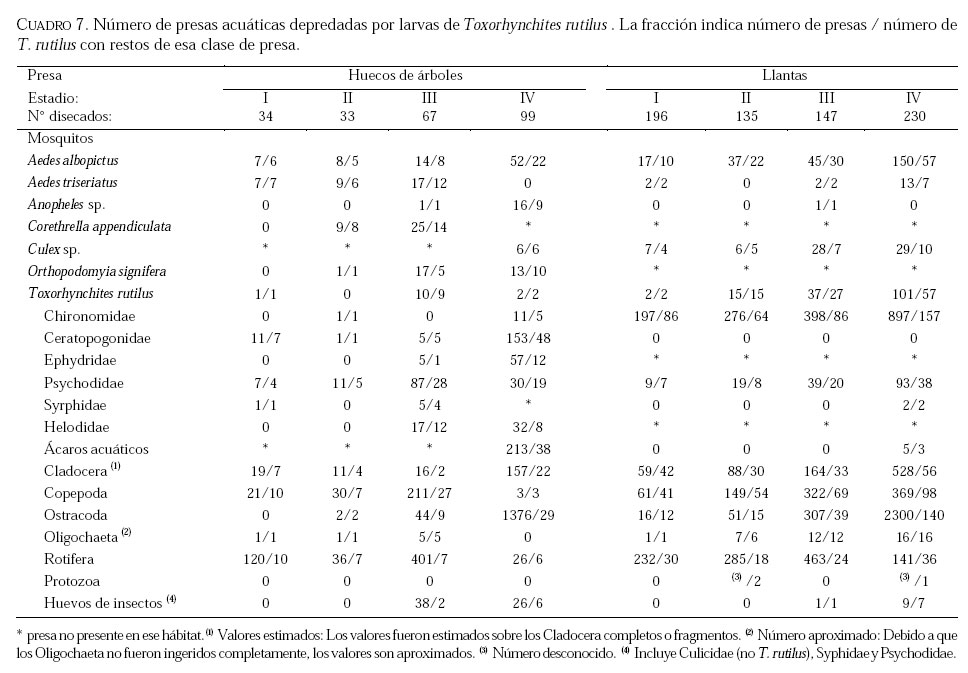
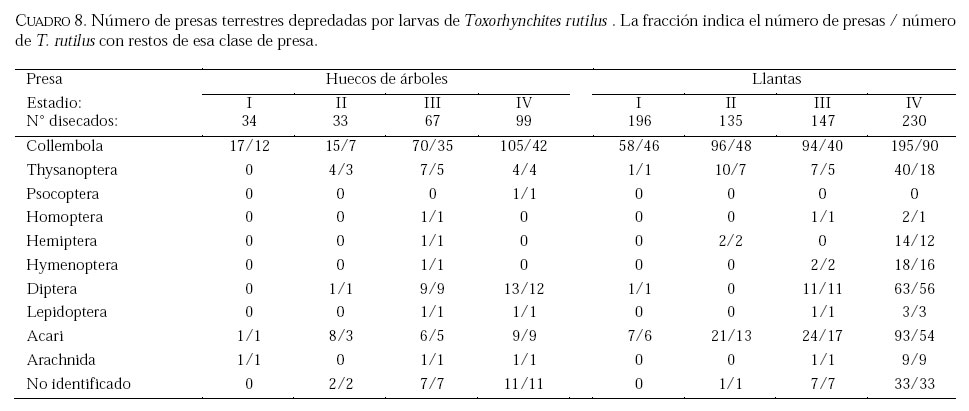
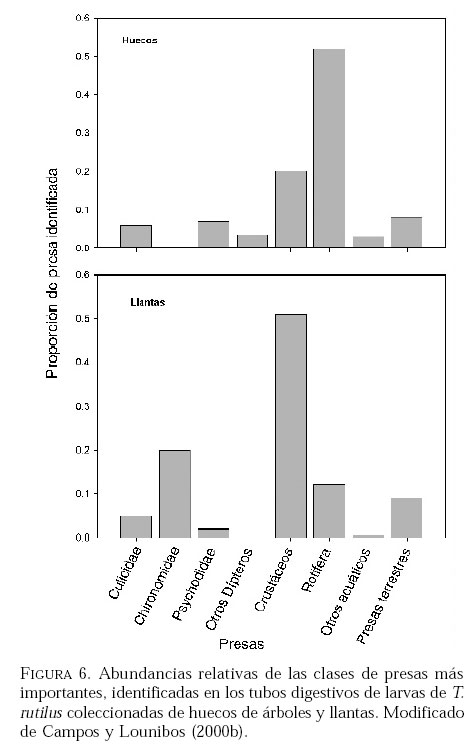
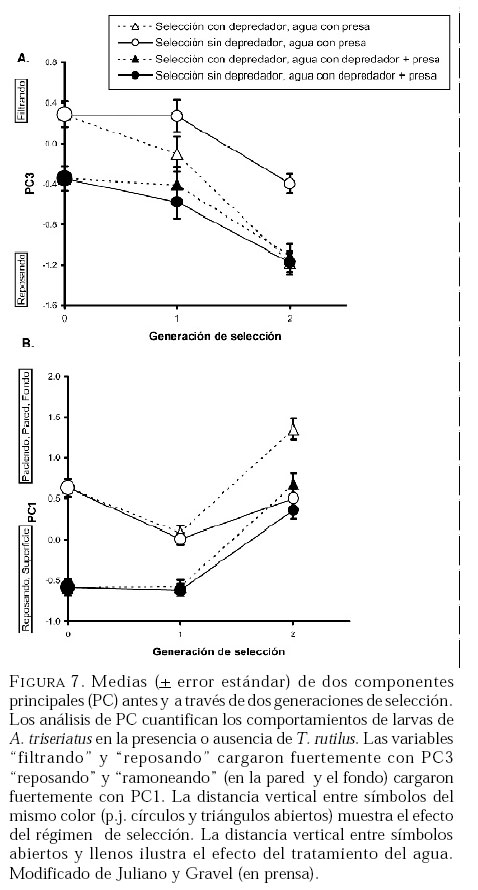
SE, Vero Beach FL 32611, EE UU. Email: lounibos@ufl.edu. Recibido: 23-x-2001 Code Number: em02021 Resumen Lounibos P, Campos RE. 2002. Investigaciones recientes sobre Toxorhynchites rutilus (Diptera: Culicidae) con referencia al control biológico de mosquitos habitantes en recipientes. Entomotropica 17(2):145-156. Esta revisión provee actualización de las investigaciones sobre Toxorhynchites rutilus, un depredador utilizado para el control de mosquitos. Una investigación a largo plazo en Florida (EUA) demostró reducción de mosquitos en huecos de árboles que fue atribuida a depredación por esta especie. Extinciones locales del vector potencial Aedes triseriatus no estuvieron asociadas con la presencia de T. rutilus en huecos. La disminución en el consumo durante la diapause debe explicar parcialmente la reducción del impacto de T. rutilus en el invierno. La sobrevivencia de T. rutilus en huecos y llantas, indican que la mortalidad es mayor en el primer estadío y que la proporción relativa de muertes es alta en el cuarto, debido al canibalismo. La diversidad de presas observadas en disecciones de T. rutilus coleccionados del campo fue amplia, y los mosquitos no fueron las presas más frecuentes. Artrópodos terrestres aparentemente capturados desde la superficie del agua, fueron comunes en la dieta, especialmente en depredadores del tercero y cuarto estadío. Juliano y Gravel (en prensa) demostraron que A. triseriatus expuestos a T. rutilus, son sujetos a una rápida selección de comportamiento(s) anti-depredador. En la naturaleza, esta presión de selección es débil, resultando en una distribución heterogénea de comportamientos anti-depredador en distintas poblaciones geográficas. En experimentos en internodos de bambú, la especie invasiva Aedes albopictus evitó la depredacion de T. rutilus mejor que la especie nativa A. triseriatus. En este trabajo, discutimos el significado de estos resultados para el control biológico de mosquitos por T. rutilus. Palabras clave adicionales: Comportamiento, ecología, depredación, mosquitos, presas. Abstract Lounibos P, Campos RE. 2002. Recent investigations on Toxorhynchites rutilus (Diptera: Culicidae) with reference to the biological control of mosquitos inhabiting containers. Entomotropica 17(2):145-156. This review describes research on Toxorhynchites rutilus, a predator used for mosquito control. A long-term investigation in Florida (USA) demonstrated a reduction in treehole mosquitoes attributable to predation by this species. Local extinctions of the potential vector Aedes triseriatus were not associated with the presence of T. rutilus in treeholes. A decline in prey consumption during diapause may partially explain the reduced winter impact of T. rutilus in the long-term data. Survivorship of T. rutilus in treeholes and discarded tires indicates that the mortality rate is highest in the first instar and that the relative proportion of deaths is also high in the fourth stage, owing to cannibalism. In dissected T. rutilus collected from the field, a broad diversity of prey types was observed, and mosquitoes were not the most common prey. Terrestrial arthropods, apparently captured from the water surface, were common in the diet, especially of third and fourth instars. Juliano and Gravel (in press) demonstrated that A. triseriatus exposed to T. rutilus experience rapid selection for anti-predator behavior(s). This selective pressure is weak in nature, resulting in a heterogeneous distribution of anti-predator behaviors in distinct geographic populations. In experiments conducted in bamboo sections, the invasive species Aedes albopictus escaped predation by T. rutilus better than A. triseriatus. We discuss the significance of these results for the biological control of mosquitoes by T. rutilus. Additional key words: Behavior, ecology, mosquitoes, predation, prey. Introducción Mosquitos del género Toxorhynchites, cuyas hembras no consumen sangre, han atraído la atención porque sus larvas depredadoras consumen otras especies de mosquitos, algunos de ellos vectores. La cría y liberación de Toxorhynchites spp. de enfermedades para el control biológico de mosquitos de los géneros Aedes y Culex que habitan en recipientes, fueron probados y utilizados durante los últimos años de la década de 1970, y comienzo de la década de 1980 (Steffan y Evenhuis 1981; Gerberg 1985). Sin embargo, un efectivo programa usando estos depredadores en la ciudad de New Orleans, Louisiana, debió ser interrumpido por el costo y tedio de la cría masiva de las larvas de Toxorhynchites spp. para las posteriores liberaciones en el campo. La incorporación de depredadores como los mencionados Toxorhynchites spp., la mayoria de los cuales son nativos de los trópicos, dentro de un esquema integrado de manejo de plagas, debe ser evaluado sobre la base de la economía actual y los beneficios prácticos, como el costo, efectividad, y los efectos secundarios de métodos de control alternativos, en particular insecticidas químicos. Toxorhynchites rutilus, es una especie nativa de Norte América, criada y liberada previamente para control de mosquitos (Focks et al. 1980, 1982, 1983) previo a que esta especie pudiese ser conveniente como un componente mas para el manejo integrado de culícidos. Señalamos dos sucesos en la década pasada que favorecen la reanudación del uso de T. rutilus para el control de mosquitos que se crían en recipientes: (1) el incremento de la incidencia del dengue en las Américas, sumada a la invasión y diseminación de Aedes albopictus (Gubler y Trent 1994; Moore and Mitchell 1997), el cual subraya la importancia de controlar los mosquitos en hábitats provistos de recipientes, y (2) el conocimiento del elevado riesgo asociado con la introducción de controles biológicos no nativos (Simberloff y Stiling 1996), lo cual debe favorecer el uso de T. rutilus en Norte América. La siguiente revisión de investigaciones es altamente selectiva, enfatizando los resultados recientes en hábitats naturales, los cuales permiten un entendimiento más detallado de la ecología de T. rutilus. Esperamos que esta revisión pueda facilitar futuras gestiones para el control utilizando esta especie u otras del mismo género. Resultados Dinámica de una población de T. rutilus durante un período prolongado. Estudios previos sobre la ecología de T. rutilus en hábitats naturales han sido limitados a menos de un año de observaciones (ej., Bradshaw y Holzapfel 1983). Lounibos (1983) empezó censos quincenales no destructivos durante un período prolongado para mostrar el patrón de abundancia de T. rutilus, y otras especies de mosquitos que habitan en huecos de árboles en el Florida Medical Entomology Laboratory (FMEL) en Vero Beach, ubicado en una región subtropical en el sur de Florida. En censos regulares de dos huecos durante 20 años, las larvas de T. rutilus estuvieron presentes aproximadamente 40-90% de los 26-27 censos realizados por año, excepto en los primeros 4-5 años de las investigaciones (Figura 1). En 1980-81 el sur de Florida sufrió una sequía que mantuvo secos a los huecos de los árboles durante muchos censos (Figura 1), causando ausencia del cuarto estadío de T. rutilus durante aproximadamente dos años (Figura 2). Para estimar el impacto de depredación de T. rutilus sobre Aedes triseriatus, el mosquito presa más abundante en los censos, se seleccionaron seis huecos y se calcularon las auto-regresiones entre los estadíos jóvenes de A. triseriatus al tiempo t, contra los estadíos maduros al tiempo t+1. En cinco de los seis huecos analizados, fueron registrados auto-regresiones significativas con auto-correlaciones de orden uno o dos lo cual indica dependencias seriales entre los numeros en censos separados por dos o cuatro semanas (Lounibos et al. 1997). Para el análisis de varianza, se utilizaron como variables dependientes las desviaciones de los números residuales de pupas y cuartos estadíos de A. triseriatus obtenidos de los modelos auto-regresivos. Las variables independientes fueron los huecos y la presencia/ausencia de un depredador de cuarto estadío. La presencia de T. rutilus y los huecos de los árboles afectaron significativamente los números residuales de pupas y de larvas del cuarto estadío de A. triseriatus, aunque la influencia del depredador fue mayor (Cuadro 1). Si bien los modelos de ANOVA explican sólo una pequeña proporción de la variación total en la abundancia residual de A. triseriatus, el efecto consistente de T. rutilus en todos los huecos examinados confirman los resultados de Bradshaw y Holzapfel (1983) que la depredación es importante en este hábitat. Para determinar si las extinciones locales de A. triseriatus estuvieron asociadas con la presencia del depredador, examinamos los datos provenientes de muestras de ocho huecos de árboles durante el período 1978-93, y registramos aquellas ocasiones en que la ocurrencia de A. triseriatus fue seguida por su ausencia en el censo siguiente mientras el hueco permaneció con agua. Para comprobar que la asociación de T. rutilus con las extinciones dentro de cada hueco eran significativamente más frecuentes que lo esperado por azar, utilizamos pruebas de muestreos aleatorios que incorporan dos diferentes hipótesis de nulidad con respecto a la subyacente distribución del depredador (Lounibos et al. 1997). Aunque las extinciones de A. triseriatus fueron registradas frecuentemente, menos de la mitad ocurrió en presencia de T. rutilus (Cuadro 2). Entre los ocho huecos no hubo asociaciones significativas entre las extinciones y la presencia de T. rutilus cuando asumimos que el depredador ocurre independientemente en el tiempo, y sólo una asociación significativa bajo la suposición de una distribución agregada. Combinando pruebas de probabilidades independientes, no se observó asociación neta significante entre las extinciones y la presencia del depredador, bajo ambas suposiciones de distribución agregada o independiente de T. rutilus. Fotoperíodo y diapausa en una población subtropical de T. rutilus de Estados Unidos de América. Durante el transcurso de los censos, particularmente a fines del otoño e invierno, los mismos individuos de T. rutilus de cuarto estadío fueron frecuentemente colectados en muestras sucesivas. Estos individuos, medidos por su peso húmedo, incrementaron su tamaño en los censos sucesivos pero no puparon hasta fines del invierno (Figura 3). Se sabe que la diapausa inducida por el fotoperíodo en poblaciones de T. rutilus del norte de los Estados Unidos, ocurre en el cuarto estadío (Bradshaw y Holzapfel 1975, Trimble y Smith 1978), por este motivo decidimos investigar si los días largos pueden inducir la diapausa en los representantes subtropicales de esta especie. Más del 50% de T. rutilus de cuarto estadío, luego de haber sido colectados como huevo en el campo del FMEL y expuestos a fotoperíodo 9 L:15 O, entraron en diapausa (Cuadro 3). Esta reacción intermedia al fotoperíodo en larvas de la península de Florida es comparada con la reacción completa de T. rutilus colonizada de una región del norte (Cuadro 3 y Lounibos et al. 1998). El consumo de presas de T. rutilus en diapausa y en no diapausa, fue comparado ofreciéndoles 25 larvas de cuarto estadío de Aedes aegypti por día. T. rutilus expuesto a 21° y 25°C durante días largos, consumió un promedio de aproximadamente 50% más presas que durante los primeros 12 días del cuarto estadío expuestos a días cortos (Figura 4). Un análisis de la varianza de medidas repetidas sobre el consumo de presas diario, indicó que la influencia de los días largos fue mayor que el efecto de la temperatura (Lounibos et al. 1998). Mientras que durante los días largos las larvas de cuarto estadío puparon durante 12-23 días, la tasa diaria de consumo de presas de T. rutilus en diapausa continuó declinando (Figura 4). Las larvas expuestas a días cortos durante aproximadamente 35 días, fueron posteriormente sometidas a días largos provocando un incremento significativo en el consumo de presas, seguido de pupación entre los 6-20 días posteriores (Figura 4 y Lounibos et al. 1998). Con fin de estimar las consecuencias producidas por la diapausa en el nivel de la población de T. rutilus, se calculó el índice de rendimiento r' análogo a la tasa de cambio per capita (Livdahl 1984) entre cohortes con y sin diapausa. Este índice incluye estimados de la fecundidad, mortalidad, y tiempo de desarrollo. En los cohortes con diapausa, los valores de r' se redujeron entre 33-47% con respecto a los valores estimados para los cohortes sin diapausa (Cuadro 4). Esta reducción en r' indica que la prolongación del desarrollo causado por diapausa, no fue equiparado por el incremento de la fecundidad de las grandes hembras que emergieron luego de que la diapausa finalizara (Lounibos et al. 1998). Curvas de sobrevivencia y tablas de vida. Aunque se han construido tablas de vida de Toxorhynchites spp. anteriormente, éstas fueron realizadas en condiciones de laboratorio (O'Flynn 1975). Nuestro propósito fue describir y comparar la duración de los estadíos inmaduros, la mortalidad específica por estadío, la sobrevivencia y la expectativa de vida de T. rutilus en huecos de árboles y llantas en desuso. Los censos diarios, se realizaron en el FMEL durante el otoño de 1997 y el verano de 1998. A diferencia de otros estudios, incluimos el estadío de huevo. Las larvas se identificaron individualmente con una marca específica producida por el corte de una seta del abdomen sin causarles daño. En ambos hábitats las curvas de sobrevivencia fueron del tipo IV de Slobodkin (1962) característica de organismos con alta mortalidad en los estadíos jóvenes. La gama de sobrevivencia desde huevo hasta adulto fue 1,8-3,4% en las llantas y 4,5-5,6% en los huecos de los árboles. El patrón de sobrevivencia fue significativo y heterogéneo entre las estaciones, y entre ambos tipos de recipientes. La tasa de mortalidad diaria fue alta en el primer estadío y baja en el cuarto (Cuadros 5 y 6). Esto se debe a que el tiempo en que las larvas permanecen en el cuarto estadío es aproximadamente el doble del tiempo que permanecen en algunos de los estadíos anteriores (Figura 5). Sin embargo la probabilidad de muerte al finalizar el estadío de larva fue igual o mayor que las del primer estadío (Cuadros 5 y 6). Las frecuentes lluvias y sequías fueron factores de riesgo poco importantes, sin embargo la probabilidad de desaparición de los huevos, y de los estadíos I y IV fue significativamente más alta en presencia de larvas co-específicas del último estadío que en su ausencia, sugiriendo que el canibalismo es una de las principales causas de mortalidad de T. rutilus (Campos y Lounibos 2000a). Presas de T. rutilus en la naturaleza. Sólo una investigación previa ha examinado las presas naturales en tubos digestivos disectados de Toxorhynchites spp. a través del análisis de exoesqueletos de artópodos (Kazana et al. 1983). Para nuestro estudio fueron seleccionadas 32 llantas y 29 huecos de árboles en el FMEL, y ocho de cada uno fueron examinados semanalmente. Las larvas de T. rutilus fueron matadas, disectadas en solución salina y se observó dentro del tubo digestivo las partes duras de las presas, lo cual permitió su identificación (Campos y Lounibos 2000b). Se disectaron 941 larvas de T. rutilus de todos los estadíos, y se reconocieron 20 entidades taxonómicas de presas acuáticas, y huevos de tres familias de dípteros. Los ostrácodos y los quironómidos fueron las presas acuáticas más abundantes en las larvas provenientes de las llantas, y los rotíferos y copépodos, en las de los huecos de los árboles (Cuadro 7). Además, fueron identificados restos de artrópodos terrestres, representados por nueve órdenes de insectos, ácaros y arácnidos (Cuadro 8), posiblemente capturadas desde la superficie del agua. Los artrópodos terrestres representaron el 8% de las presas identificadas de los huecos de los árboles y 9% de las llantas (Figura 6). Sin embargo las presas más frecuentemente halladas fueron los microcrustáceos, los cuales representaron el 51% de las presas identificadas en T. rutilus de las llantas y el 20% de los huecos de los árboles (Figura 6). El coeficiente de selectividad, R, fue calculado para las presas del orden Diptera y para cada estadío del depredador. La heterogeneidad significativa entre los valores R (Campos y Lounibos 2000b) entre las llantas fue atribuida, en parte, a la selección principal de ciertas especies de mosquitos como Aedes albopictus (Skuse) y larvas de Psychodidae como presa, y por la escasa selección de larvas de T. rutilus y Orthopodomyia signifera (Coquillett). La dieta de T. rutilus en las llantas fue semejante entre los distintos estadíos, mientras que en los huecos hubieron diferencias, posiblemente causadas por la mayor diversidad de presas presentes en ese hábitat. Evolución del comportamiento de defensa de las presas. Estudios de laboratorio realizados por Livdahl (1979, 1984) han indicado que los A. triseriatus alopátricos con T. rutilus fueron menos exitosos en escapar de la depredación, que los A. triseriatus cuyo origen fue simpátrico con el depredador. Posiblemente, los A. triseriatus simpátricos han desarrollado defensas anti-depredador en presencia de la presión selectiva de depredación. Posteriormente, Juliano y Reminger (1992) identificaron diferentes comportamientos entre poblaciones como causas inmediatas de las diferencias en la vulnerabilidad hacia la depredación. Sin embargo, comparaciones posteriores entre larvas de ocho poblaciones de esta presa, no demostraron tendencia consistente entre poblaciones alopátricas y simpátricas que pueda confirmar que estos comportamientos evolucionan donde las presas y los depredadores ocurren juntos (Juliano et al. 1993). Recientemente Juliano y Gravel (en prensa), desarrollaron un experimento en el cual dos generaciones de A. triseriatus fueron expuestas en el laboratorio a la depredación por T. rutilus. Las líneas de selección fueron reducidas por 50% cada generación por la depredación de T. rutilus, mientras las líneas de los controles fueron reducidas igualmente por traslados aleatorios de larvas. Luego de dos generaciones de selección, los resultados mostraron una reducción significativa en la actividad de filtrado entre larvas expuestas al depredador (Figura 7). Esos resultados muestran que la depredación por T. rutilus puede seleccionar rápidamente cambios en el comportamiento. Además sugieren que la falta de diferenciación consistente con respecto al comportamiento entre poblaciones simpátricas y alopátricas de A. triseriatus, puede ser el resultado de presiones de selección débiles o heterogéneas en algunas poblaciones naturales expuestas a la depredación. Una especie invasiva como presa de T. rutilus. Desde su establecimiento en
Florida en 1986, la especie de mosquito invasivo Aedes albopictus llegó a
ser un habitante frecuente de recipientes con agua (O'Meara et al. 1993). Aunque
la distribución de Aedes aegypti se redujo por causa del invasor (O'Meara
et al. 1995), no se observó tal efecto en A. triseriatus. Debido a que
A. albopictus y A. triseriatus ocurren frecuentemente juntos en recipientes
con T. rutilus, Lounibos et al. (en prensa) compararon la susceptibilidad de
las dos especies al depredador en internodos de bambú. La sobrevivencia
hasta imago fue significativamente más alta en A. albopictus que en
A. triseriatus cuando larvas del primer estadío de las dos fueron expuestas
a una larva de T. rutilus, empezando uno o tres días después
de la eclosión Discusión La persistencia de T. rutilus en hábitats naturales (Figura 1) produce un bajo nivel de control durante períodos largos (Cuadro 1) y la re-introducción de este depredador debería ser innecesaria. Sin embargo, cuando ocurre extinción en el hábitat debido a las sequías, la recolonización natural por Toxorhynchites spp. puede ser lenta (Figura 2 y Lounibos 1979). La preferencia de T. rutilus por huecos de árboles que resisten la desecación (Bradshaw y Holzapfel 1988), puede ser considerada como una adaptación para maximizar la persistencia de la especie, cuyos estados inmaduros no son resistentes a la sequía. El impacto que produjo T. rutilus en reducir el número de A. triseriatus durante períodos prolongados fue relativamente pequeño comparado a reducciones de 74% de Aedes aegypti y Culex quinquefasciatus que produjeron las liberaciones experimentales de este depredador cada diez días durante períodos cortos en la ciudad de New Orleans (Focks et al. 1982). Serían necesarias liberaciones de T. rutilus (los estadíos e intervalos dependientes de las condiciones del hábitat y el ambiente), para alcanzar un mayor nivel en el control de mosquitos, y además, para incrementar las poblaciones naturales disminuidas por las sequías. Al menos tres estudios previos en sistemas acuáticos han indicado que los depredadores no específicos pueden causar extinciones locales de mosquitos (Bence 1988, Blaustein et al. 1995, Bradshaw y Holzapfel 1983), los cuales pueden desestabilizar la dinámica poblacional de la interacción depredador-presa (Murdoch y Bence 1987). Los tres estudios incluyeron una investigación previa de T. rutilus en huecos de árboles en otro sitio de Florida (Bradshaw y Holzapfel 1983), sin embargo esta investigación de nueve meses de duración fue corta comparada con los resultados de los estudios a largo plazo resumidos aquí. Tampoco fueron incluidos los períodos de tiempo en que el depredador estuvo inactivo. Los estudios de Bradshaw y Holzapfel (1983) y Lounibos et al. (1997) realizados en Florida, discrepan entre sí, debido a los diferentes procedimientos estadísticos, y a las diferencias entre la dinámica de las poblaciones locales. El escaso consumo de presas por larvas de T. rutilus en diapausa (Figura 4), la falta de asociación entre la presencia del depredador, y la extinción de A. triseriatus en los censos durante períodos prolongados, pueden ser algunas de las causas del bajo impacto de la depredación. La investigación de Bradshaw y Holzapfel (1983) en el norte de Florida no incluyó muestreos en el invierno durante el período de diapausa de T. rutilus. Para propósitos de control biológico, las liberaciones de T. rutilus que no estén en diapausa durante el invierno pueden compensar el reducido r' atribuido a la diapausa en poblaciones naturales (Cuadro 2), y consecuentemente, aumentar la mortalidad de mosquitos presas por depredación. Presumimos que gran parte de la mortalidad de las larvas de T. rutilus en la naturaleza es causada por canibalismo. Esto ha sido documentado en estudios previos (Bradshaw y Holzapfel 1983; Lounibos et al. 1996), como así también por nuestras disecciones de los tubos digestivos (Cuadro 7). Diferencias entre el tamaño de T. rutilus de cuarto estadío y otros más pequeños habitando el mismo recipiente, frecuentemente conducen al canibalismo (Lounibos 1985; Lounibos et al. 1996), lo es consistente con patrones de depredación interespecíficos entre otros carnívoros limitados por alimento (Elgar y Crespi 1992). Nuestros análisis del contenido de los tubos digestivos de T. rutilus, revelaron una dieta más diversa (Cuadros 7, 8) que la consumida por Toxorhynchites theobaldi en un cementerio de Caracas (Kazana et al. 1983). Estos datos indican que la amplia dieta en Florida es una consecuencia de la alta diversidad de la fauna en las llantas y huecos de árboles (Campos y Lounibos 2000b), comparado con los floreros urbanos ocupados por T. theobaldi. En el cementerio de Venezuela, el mosquito Culex fatigans fue más común que otras presas, mientras que en nuestras llantas y huecos, las larvas de quironómidos y psicódidos fueron respectivamente las presas consumidas con mayor frecuencia. En tres tipos de fitotelmatas en la selva húmeda de Venezuela, la depredación por Toxorhynchites haemorrhoidalis alteró los patrones de abundancia, y el tamaño de las larvas de cinco familias de dípteros (Lounibos et al. 1987). Experimentos para medir el tiempo en que las presas más comunes en llantas y huecos tardan en pasar por el intestino de T. rutilus, demostraron que el consumo continuo de presas disminuye el tiempo de la digestión en relación a las larvas en ayuno (Campos y Lounibos 2000b). Recientemente se ha podido apreciar que los mosquitos pueden desarrollar resistencia genética contra insecticidas microbiales usados para su control (ej., Becker y Ludwig 1993, Rao et al. 1995). Por otro lado, la rápida selección en comportamientos anti-depredador en A. triseriatus (Figura 7) indica que las presas pueden desarrollar defensas frente a un depredador cuando es utilizado como control biológico. Sin embargo, la evidencia disponible sobre depredadores y parasitoides, sugiere que tales defensas a causa de fuerte y rápida selección ocurre infrecuentemente en la naturaleza, quizás porque la selección es espaciada y temporalmente débil en ambientes heterogéneos (Holt y Hochberg 1997). Los métodos citados para cuantificar las defensas anti-depredador provistos por Juliano et al. (1993) y Juliano y Gravel (en prensa) pueden ser de gran interés para medir cambios en el comportamiento de las presas, y en la eficiencia del depredador durante un programa de control biológico, el que artificialmente intensifica las presiones de selección natural. Las investigaciones han demostrado que T. rutilus, y otras especies del mismo género, reducen significativamente los números de mosquitos vectores. Programas de control integrado deben preservar las poblaciones naturales de las especies indígenas de Toxorhynchites. Liberaciones para incrementar el impacto de este depredador requieren las crias de la presa y tienen mayor valor donde las hembras liberadas oviponen directamente en los recipientes que contienen los estadíos acuáticos de vectores y plagas. En tales condiciones, los Toxorhynchites pueden ser un componente valioso de un programa integrado de control. Agradecimientos Deseamos agradecer los comentarios de los Drs. S. Juliano, J. Rey y R. Darsie y la invitación del Dr. M. Badii (UANL, Nuevo León, México) a participar en el taller Manejos Integrados de Mosquitos en donde se presentó una primera versión de este trabajo. Esta revisión se aprobó en parte por la subvención AI-47793 del Instituto Nacional de Salud de EU y es Agricultural Research Station Publication No. R-08463 de la Universidad de la Florida. Referencias
© 2002- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em02021t8.jpg] [em02021t5.jpg] [em02021t2.jpg] [em02021f2.jpg] [em02021t6.jpg] [em02021t4.jpg] [em02021f1.jpg] [em02021t7.jpg] [em02021t3.jpg] [em02021f7.jpg] [em02021f3.jpg] [em02021f6.jpg] [em02021t1.jpg] [em02021f5.jpg] [em02021f4.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}