 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica antes/formerly Boletín de Entomología Venezolana, Vol. 20, No. 1, April, 2005, pp. 59-65 Interaction between ants and orchids in the Soconusco region, Chiapas, Mexico. Interacciones entre hormigas y orquídeas en la region de Sonocusco, Chiapas, México Anne Damon and Marco Antonio Pérez-Soriano El Colegio de la Frontera Sur (ECOSUR), Carretera Antiguo Aeropuerto Km. 2.5, C.P. 30700, Tapachula, Chiapas, México. Code Number: em05009Abstract DAMON A, PÉREZ-SORIANO MA. 2005. Intereaction between ants and orchids in the Sonocusco region, Chiapas, Mexico. Entomotropica 20(1): 63-69. Ants were seen to visit extrafloral nectar sources mostly on the flower bases and petiole junctions of Encyclia cordigera, E. chacaoensis and E. belizensis var. parviflora and the production of extrafloral nectar in relation to the presence of ants was observed during the flowering periods of these orchids in the Regional Botanical Garden “El Soconusco” and the orchidarium Santo Domingo, in Chiapas, Mexico. Ants were also seen to feed on secretions produced at flower bases and on the surface tissue and tips of buds of Catasetum integerrimum and Cycnoches ventricosum. Mention is included of ants associated with other species of orchids collected in the field. Eleven species of ants were fully identified out of a total of 19 species collected. Most species were common inhabitants of secondary vegetation, with some species registered as pests in agroecosystems and human dwellings. It was not clear how the plants benefited by investing scarce resources in the provision of food to attract ants, but a defense role is possible for one species, Camponotus novogranadensis, which was observed to be aggressive when the plant was disturbed by touching during visits to the extrafloral nectaries of E. chacaoensis. It is suggested that E. belizensis var. parviflora benefits from the presence of Crematogaster aff. torosa resident in old pseudobulbs, which effectively act as domatias, by absorbing nutrients from organic wastes produced by those ants. Additional key words: Ants, mutualism, orchids, extrafloral nectar, reproductive structures. Resumen DAMON A, PÉREZ-SORIANO MA. 2005. Interacciones entre hormigas y orquídeas en la region de Sonocusco, Chiapas, México. Entomotropica 20(1): 63-69. Observamos hormigas visitando fuentes de néctar extraflorales en la base de las flores y la unión de los pecíolos de Encyclia cordigera, E. chacaoensis y E. belizensis var. parviflora, también observamos la producción de néctar extrafloral en la relación con la presencia de hormigas durante el período de floración de estas orquídeas en le Jardín Botánico Regional “El Sonocusco” y el orquidiario Santo Domingo, en Chiapas, México. También observamos hormigas alimentándose en las secreciones de la base de las flores, en el tejido superficial y en los ápices de los retoños de Catasetum integerrimum y Cycnoches ventricosum. Hacemos mención de observaciones de hormogas asociados con otras especies de orquídeas en el campo. De un total de 19 especies de hormigas colectas, 11 fueron identificadas hasta especie. La mayoria de las especies son habitantes comunes de vegetación secundaria, con algunas especies registradas como plagas en agroecosistemas ó en viviendas. No está claro cómo las plantas se benefician al invertir recursos escasos al proveer alimento para atraer hormigas, pero al menos en el caso de una especie, Camponotus novogranadensis, es posible que exista un rol defensivo ya que se evidenció un comportamiento agresivo cuando se tocaba la planta (E. chacoensis) y esta especie estaba visitando nectarios extraflorares. Se sugiere que E. belizensis var. parviflora se beneficia de la presencia de Crematogaster aff. torosa residentes en seudobulbos viejos, que fungen como domacios absorviendo nutrientes de desechos orgánicos producidfos por estas hormigas. Palabras clave adicionales: Hormigas, mutualismo, orquídeas, nectar extrafloral, estructuras reproductivas. Introduction The state of Chiapas, in the southeast of Mexico is renowned for its biodiversity and also for its social, political and economic problems that have led to serious environmental deterioration in recent decades. Within this scenario, orchids are disappearing at an alarming rate and only small fragmented populations persist in the wild. The project “Ecology and rustic, sustainable production of orchids” based in “El Colegio de la Frontera Sur (ECOSUR)”, in Tapachula, Chiapas is dedicated to the study of key aspects of the ecology and cultivation of native orchids, aimed at the conservation and recovery of these plants and the ecosystems that support them. Very few studies have been carried out on the ecology of the estimated 25,000- 30,000 members of the orchid family (Pridgeon, 2000), and studies on the tropical epiphytic species are particularly scarce, possibly due to the inaccessibility and rarity of many of these plants. In particular, little attention has been paid toward the implications of the presence of ants that are frequently seen patrolling the surface of the reproductive structures of orchids and of the nature of the secretions produced by these plants (but see Jeffrey et al., 1970). The characteristics of ants which have permitted the development of the varied and intricate mutualistic interactions with plants that are of vital importance to the persistence of tropical forest ecosystems are widely discussed in the literature (Wilson, 1971; Bentley, 1977: Janzen, 1977; Beattie, 1985; Holldobler and Wilson, 1990; Huxley and Cutler, 1991; Madden and Young, 1992; Blüthgen, 2003; Davidson et al., 2004, etc.). Dietary flexibility, based on a requirement for soft or liquid food which incurs no damage to plant tissue, has permitted ants to enter into close and often mutualistic relationships with plants, wherein ants may remain fixed on a preferred plant but forage temporarily off that “base” plant during periods of resource deficiency (Rico-Gray, 1993). The majority of studies upon ant-plant mutualism have attempted to show that ants defend their host plants against herbivores in exchange for a secure and complete food source which can be presented as extrafloral nectar (EFN) or different categories of food bodies (Janzen, 1966 and 1969); this may be particularly important for monocotyledonous plants, such as orchids, that cannot repair tissue damaged by herbivory (Almeida & Figueiredo, 2003). Such behavior has been observed in the case of species within the orchid genus Coryanthes (Walters & Bergold, 2001) and ants present on the surface of plants of Encyclia cordigera (Kunth) Dressler were found to be predators of insects with a beneficial net effect, in favor of the orchid. One of the very few detailed studies was carried out by Almeida & Figueiredo (2003) on the interaction between the epiphytic orchid Epidendrum denticulatum Jacq. and Camponotus sericeiventris (Guérin-Méneville) (Formicinae) and Ectotomma tuberculatum (Olivier) (Ponerinae). They concluded that ants protected reproductive structures during the relatively long flowering periods which are associated with low visitation rate by pollinators, which is characteristic of the majority of orchid species. Extrafloral nectaries (EFNs) are found in 68 families of plants (Elias, 1983), are not directly involved in pollination strategies and offer sweet secretions which attract the attention of ants and give rise to interactions of various intensities from casual visits to obligate mutualism. The fact that ants are rarely seen in the role of pollinators is widely discussed in the literature (e.g. Janzen, 1969; Beattie, 1985; Puterbaugh, 1998), they have been reported to secrete antibiotic substances that inhibit pollen tube germination and, more recently, Ghazoul (1999) reported that ants were deterred by repellants in the floral tissue of many plants. EFN, therefore, offers plants a way of attracting and co-opting the services of ants without interfering with pollinators. The literature mentions 36 genera of orchids, the vast majority of which are epiphytic, which produce EFN usually associated with reproductive structures (Peakall 1994). Caularthron (Diacrium) bilamellatum (Reichb.f.) R.E. Schultes is the only orchid known to present EFNs on non reproductive structures, in this case on the leaves (Fisher et al. 1990); this species has been seen by the author to deteriorate in the absence of ants. In the most developed examples of ant-plant mutualism, the plant may also offer refuges or nesting sites, termed domatia (Koptur 1984; Keeler, 1989; Rico Gray et al., 1989) and accumulations of organic matter derived from ant activity within these structures may serve as a nutritional source for the host plants. The literature includes mention of various species of orchids that maintain colonies of ants within old, hollow pseudobulbs or sustain ants patrolling the surface of the plant, such as species of Cattleya, Coryanthes, Dendrobium, Oncidium, Schomburgkia, Vanda and Vanilla (Beattie, 1985; Hölldobler & Wilson, 1990). This study sought to observe in detail the daily and long term activity of ants in relation to the reproductive phenology and EFN production of Encyclia cordigera (Kunth) Dressler, Encyclia chacaoensis (Reichb.f.) Dressler and Encyclia belizensis var. parviflora (Regel) Dressler & Pollard. Brief observations were also carried out on Catasetum integerrimum Hook and Cycnoches ventricosum Bateman and a few species of ant were found associated with orchids collected in the field in the coastal region of Soconusco. Attempts were made to identify all of the ant species collected. These five orchid species are all found in the region of Soconusco, in the state of Chiapas, southeast Mexico. E. chacaoensis is now the most common, weedy and opportunistic species in the region, found from 10-1000 masl; E. cordigera is found on mature trees in the better conserved pastures and coffee and cocoa plantations from 10-500 masl and E. belizensis var. parviflora and the very similar E. adenocarpa (La Lave & Lex.) Schltr. are coastal species and are now exceedingly rare in the region due to the destruction of the narrow belts of low spiny forest that used to line the mangrove swamps where these orchids thrive. C. integerrimum is a weedy species found in varying habitats at lower altitudes and prefers rotting wood. C. ventricosum is a beautiful and now relatively rare orchid, which prefers shaded natural forest at low altitudes but can also be found in traditional coffee and cocoa plantations. C. integerrimum and C. ventricosum are both deciduous species. Materials and Methods Study sites: Ant activity was observed in the Regional Botanical Garden “El Soconusco” at 80m.a.s.l. in the municipality of Tuzantán (Site 1) and in the orchidarium Santo Domingo at 900m.a.s.l. in the municipality of Unión Juárez (Site 2), both in the state of Chiapas, southeast Mexico. Collecting trips were carried out in rural areas surrounding the two study sites. Identification of ants: Ant samples collected from all five species subjected to detailed study, and those collected in the field, were sent to the Institute of Ecology A.C. in Xalapa, Veracruz, for identification. Observations of ant behavior on the reproductive structures of E. chacaoensis, E. cordigera, E. belizensis var. parviflora were carried out during the flowering period of these three species, every fifteen days, from 11/04/03 to 28/06/03. A total of 32 plants that demonstrated ant activity were selected and marked to facilitate continued recognition during the study period (8 E. belizensis var. parviflora, 8 E. chacaoensis and 6 E. cordigera in Site 1 and 5 E. chacaoensis and 5 E. cordigera in Site 2). Observations consisted of: 1. Structures patrolled by the ants; 2. Feeding sites of the ants, such as buds, fruits, flower bases, petiole junctions; 3. Signs of EFN production; 4. Presence of domatias; 5. Interaction of visiting ants with other insects, such as passivity, aggression or escape; 6. No. of species of ant present on the plant; 7. The no. of ants that visited the observed feeding sites during five minutes, every two hours, between 08.00 and 18.00hrs. Ants were considered to be feeding when they remained stationary for several seconds with the mouth in contact with the surface of the orchid. Results Identification of ants: A total of 19 species of ants were found associated with 9 species of orchids in the region of Soconusco during the period of study (Table 1). A total of 12 species of ants were associated with the reproductive structures of E. cordigera, E. chacaoensis and E. belizensis var. parviflora and two species were associated with the reproductive structures of each of Cycnoches ventricosum and Catasetum integerrimum (Table 2a). The ant species most commonly associated with the orchids was Crematogaster aff. torosa, found associated with three of the orchids studied in detail and three other species in the field. Table 2b presents a list of the ants found associated with orchids during field trips to areas near to sites 1 and 2. Table 1. Ant species found associated with orchids in the region of Soconusco.
Table 2a. Distribution of ant species among the five orchid species studied.
Table 2b. Ant species found associated with orchids in the field.
General characteristics The characteristics of the interaction between the various species of ants observed and the three species of Encyclia are presented in Table 3. The ants patrolled the mature flowers in all cases, and also the immature flowers of E. chacaoensis and E. belizensis var. parviflora and the buds of E. cordigera y E. chacaoensis. Feeding sites were more specific and for all species, much time was spent feeding at the base of mature flowers and at the union, or internodes between the petioles of adjacent flowers. The ants also scraped the general surface of the flower buds of E. chacaoensis and E. cordigera and fed from secretions produced at the immature flower tips of E. belizensis var. parviflora y E. chacaoensis. All three species of orchid showed visible signs of EFN production. E. belizensis var. parviflora was the only species which presented domatia, and on only one of the plants studied, in old, dry pseudobulbs, accessed by conspicuous holes near the base.
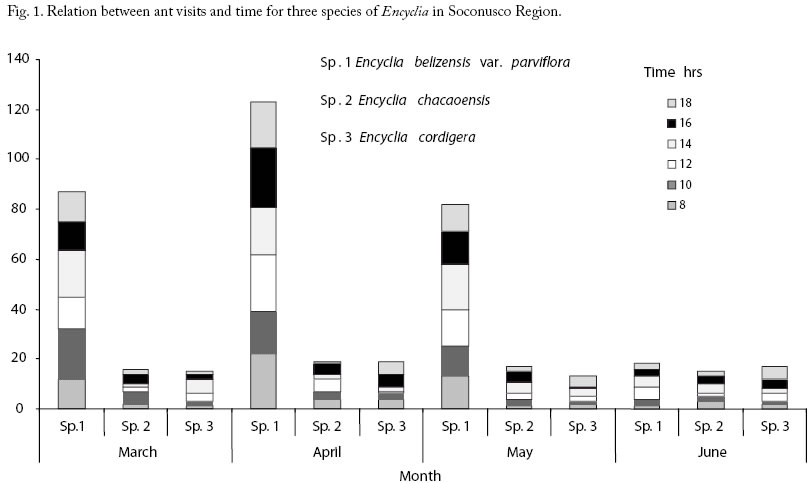
Ant behaviour Camponotus novogranadensis was the only one of the seven species of ant associated with E. chacaoensis observed to be aggressive and to actively defend the plant; this ant recruited to the point where the plant was touched (Table 3). These seven species of ant were never found together on the same plant. The ants patrolling E. cordigera were passive and did not respond to disturbance (touching the plant), and visits were the least frequent of the three species. The ants that nested in the domatia on one of the plants of E. belizensis var. parviflora left their nest and fled the plant when it was touched, and clearly do not defend their host plant. No pests were present on the orchid plants at any time during the study, we could not, therefore, evaluate the value of ant presence in relation to herbivory. Frequency of visits Visits were most frequent to the EFNs of E. belizensis var. parviflora and peaked at 14.00hrs, with an average of 15 independent visits during the 5 min periods. Activity was reduced toward dusk, with 10.75 visits per 5 minute period at 18.00hrs. EFNs of E. chacaoensis and E. cordigera received a constant trickle of an average of 2.75 and 2.25 visits, respectively, per 5 min period during the whole day. It is clear from Fig. 1 that E. belizensis var. parviflora received the most visits during the study period, involving only one species of ant. Most visits were observed at the time of peak flowering in April, with a sudden reduction in activity in June, when flowering ceased. In previous observations, in the field and in Site 1, the same species of ant was seen to continue patrolling plants of E. belizensis var. parviflora during the development and maturation of the seed capsules and to feed from secretions produced at the base of the withered flower. It is not clear why, in these observations, visits ceased at the end of the flowering period despite the presence of developing seed capsules. E. chacaoensis and E. cordigera show a different pattern of ant interaction, in which a similar level of ant activity on the plants was maintained at all times. These orchids flowered during March to May, after which the ants switched their attention to young shoots. Discussion Extrafloral nectaries produce secretions containing sugars (Baskin and Bliss, 1969) which attract the attention of ants and give rise to interactions of various intensities from casual, opportunistic visits to obligate mutualism; these interactions have been little studied in orchids. Most orchid species are notorious for offering no reward to pollinating insects, supposedly for reasons of resource economy in stressful environments. In that context, it is surprising to find that orchids invest scarce energy resources in the production of EFN to attract ants. In the case of E. belizensis var. parviflora, E. cordigera and E. chacaoensis, well defined areas on flower bases and petiole junctions were visited by the ants. There was a greater frequency of ant visits to EFNs of E. belizensis var. parviflora, although by one species of ant only, Crematogaster aff. torosa, suggesting a degree of specialization towards a mutualistic interaction and this orchid species also offered refuge to the ants in the form of hollow, dry, old pseudobulbs, with access holes at the base. The benefit to the orchid, in this case, is likely to be alimentation via absorptive cells in the lining of the domatia, and not defense. The ant in question, C. aff. torosa was also found feeding upon the EFNs of other orchids in this study. In general, the activities of these ant visitors may contribute towards the persistence of reproductive structures, without interfering with the pollination mechanism. Visits by pollinators for many species of orchid are rare and intermittent; it may benefit the plant to protect the flowers so that they persist for a greater period of time, increasing the likelihood of receiving a visit. However, in this study, pests were not a problem, so the contribution of patrolling ants as a defense strategy could not be evaluated. Furthermore, most of the ants observed were seen to be passive or neutral towards disturbance involving “their” plant, which does not suggest a defense role, except in the case C. novogranadensis associated with E. chacaoensis. Further studies are required to see whether EFNs are present on immature flower tips and shoots, where ants were also seen to be feeding on occasion, or whether the ants are simple rasping and feeding on surface tissue. A comparative analysis of the content of the EFN produced at different times of day and at different phenological stages would be informative. However, the minute quantities of EFN produced would require the destructive sampling of many plants in order to obtain workable samples, which was not compatible with the aims of this study and the number of plants available. Most of the ants collected in this study are common, amply distributed species, with many references to invasion of human dwellings and agroecosystems where they may be considered as pests (Klotz et al, 1995). Furthermore, most species are cited as generalized scavengers and frequent, opportunistic visitors to extrafloral nectaries. Wasmannia auropunctata, given the name “little fire ant” is native to continental Central and South America and reported as a particularly aggressive and invasive species (Wetterer et al. 1999). Some species, such as Crematogaster carinatus and C. torosus are cited as arboreal species, abundant in mature, tropical rainforest canopies but also adaptable to seasonally dry areas, highly disturbed areas, and pasture edges (Longino, 2003). C. carinatus was cited as one of the dominant species in ant mosaics in Colombian rainforest and has been found sharing nests with Dolichoderus debilis, also found attending the EFN’s of orchids in this study. Cyphomyrmex rimosus is a fungus growing species (Snelling and Longino, 1992). Pseudomyrmex gracilis associates with acacias (Cronin, 1998, Wetterer and Wetterer, 2003). W. auropunctata, C. rimosus and different species of Pheidole and Dolichoderus were found nesting in random association with tank bromeliads in the Venezuelan Amazonia (Blüthgen et al 2000). This study presents evidence of encounters between opportunistic, foraging ants and 4 species of non myrmecophytic orchids, E. chacaoensis, E. cordigera, C. integerrimum and C. ventricosum. Interaction between E. belizensis var. parviflora and Crematogaster aff. torosa may be more specific and this orchid may be considered as a mymecophyte. However, there was no evidence to suggest that this orchid benefited from reduced herbivory during the study period but is likely that the resident ants provide organic matter for the plant. The most important question arising from this study is why do resource-constrained orchids produce EFN, that is attractive only to ants, when there is minimal likelihood of pest attack and the ants attracted are mostly non-aggressive and would not, in any case, repel insect pests? The study was carried at the beginning of the rainy season, humidity was not particularly high (max. 55% at 16.00hrs) so passive excretion, thought to be the evolutional origin of sweet, watery secretions by plants, was not an acceptable explanation. It is possible that the original target of these EFNs were other species of ants present in undisturbed tropical humid forests, which did play a defensive role and that in highly disturbed environments the specialized interactions may break down or become obsolete. The author is also finding that increasingly rare pollination events in orchids in Soconusco are being carried out by a few, common and generalist pollinators (Damon, unpublished). The interaction clearly lacks a feedback mechanism or synchrony between the two organisms (Hölldobler and Wilson 1990) and resource limited plants continue to secrete extrafloral nectar despite the presence of unrewarding, opportunistic ant species which may, therefore be considered as nectar robbers. Acknowledgements We are grateful to Luis Quiroz Robledo of the Institute of Ecology A.C. in Xalapa, Veracruz, for the identification of the ants collected during this study. References
© 2005- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em05009f1.jpg] |
| |||||||||
{kind=link}