 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
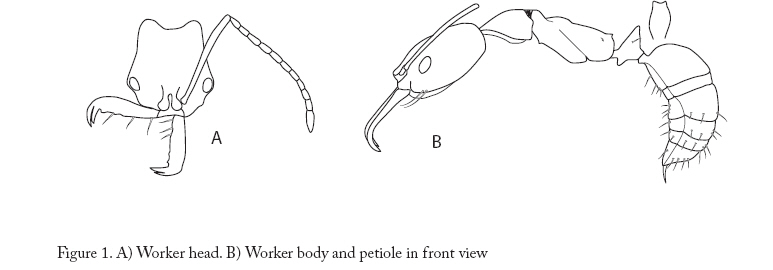
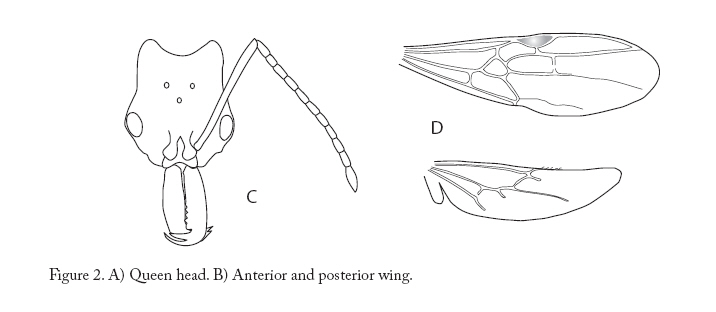
Entomotropica, Vol. 23, No. 1, Abril/April, 2008, pp. 97-102 A new species of Anochetus (Hymenoptera: Formicidae: Ponerini) from Argentina and Paraguay, associated with a leaf cutter ants Una especie nueva de Anochetus (Hymenoptera: Formicidae: Ponerini) asociada a hormigas cortadoras de hojas de Argentina y Paraguay María Constanza González-Campero1, Luciana Elizalde2. 1Instituto Superior de Entomología (INSUE). Fac. Ciencias Naturales e IML. Miguel Lillo Nº 205 T4000JFE -. San Miguel de Tucumán, Argentina, Phone 54-381-4232965. cgonzalezcampero@csnat.unt.edu.ar Code Number: em08004 Abstract A new species of the ant genus Anochetus is described from Argentina and Paraguay. Anochetus miserabilis González-Campero & Elizalde sp. nov. is included within the “emarginatus” group, which is distributed in northern South America and the Antilles. With the inclusion of this new species, the distribution of the group is extended to southern South America. Observations of its biology and peculiar association with a leaf cutter ant species are given. In addition, illustrations and a key for the worker caste of the species of the genus present in Argentina and Paraguay are included. Additional key words: Anochetus miserabilis sp. nov., Acromyrmex hispidus, ant association, emarginatus group, taxonomy. Resumen Se describe una nueva especie del género Anochetus para la Argentina y Paraguay. Anochetus miserabilis González-Campero & Elizalde sp. nov. se incluye dentro del grupo “emarginatus”, el cual se distribuye en el norte de Sudamérica y las Antillas. Con la inclusión de esta nueva especie, la distribución de “emarginatus” se extiende hasta el sur de Sudamérica. Se incluyen observaciones de su biología y su curiosa asociación con hormigas cortadoras de hojas. Se presentan, además, ilustraciones y una clave para obreras de Anochetus de la Argentina y Paraguay. Palabras claves adicionales: Anochetus miserabilis sp nov, Acromyrmex hispidus, asociación de hormigas, grupo “emarginatus”, taxonomía. Introduction Anochetus (Mayr) belongs to the Ponerinae subfamily (Bolton 2003). These ants inhabit every tropical and subtropical region in the world, building their nests generally in fallen trunks, rotting logs or in leaf litter. Twenty-three species have been cited for America, from the Mexican tropics and some Caribbean islands to northern Argentina (Lattke 2003). This genus has scarcely been studied for the last few years, being the last generic review proposed in 1978 by Brown, where every caste is described and keys for workers from the Neotropical region, Africa, Spain, Madagascar, Australia, Asia and the Pacific region are given. His work includes the most complete review of the genus, with a discussion on the taxonomy of every species and data on its biology. At present, there are only three species cited for Argentina: Anochetus altisquamis Mayr, A. neglectus Emery (Kusnezov 1963; Kempf 1972; Cuezzo 1998) and A. diegensis Forel (Leponce et al. 2004). The only species cited for Paraguay is A. neglectus (Wild 2006). In this work we describe a new species of Anochetus from Argentina and Paraguay, and provide some observations of their biology, illustrations and a key at species level. Materials and methods Samples were obtained from field collections carried out in the provinces of Chaco and Formosa (Argentina) and in the Alto Paraguay Department (Paraguay). Using Brown’s key (1978), comparisons with the samples deposited at the Collection Instituto-Fundación Miguel Lillo (IFML, Tucumán, Argentina) and resources from internet (Longino 2006) specimens were identified. The specimens (holotype and paratypes) will be deposited at the IFML Collection. Measurements The samples were studied under a stereomicroscope at a 40 X magnification. The following measurements (mm) were determined: Head Length (HL). Maximum head length in front view, on a straight line from the posterior cephalic margin to the anterior border of the mandibular condyles. Head Width (HW). Maximum width of the cephalic capsule, measured at the eye level. Mandibular Length (ML). Maximum head length, measured at the same plane as HL, from the mandibular apex to its insertion at the head. Scape Length (SL). Maximum scape length in a straight line, without including the basal constriction or neck near the condyle. Eye Length (EL). Higher length of the compound eye in a lateral view. From these measurements, the following indexes were obtained: Cephalic Index (CI). HW/HL*100. Mandibular Index (MI). ML/HL*100. Scape Index (SI). SL/HW*100. Key for Anochetus workers from Argentina and Paraguay (modified from Brown’s key, 1978) 1 Large species with long mandibles (ML > 1.1mm), combined length of head and closed mandibles (HL+ML) > 2.5 mm; inner border with 10 or more preapical teeth (in addition to 3 apical teeth).......................miserabilis sp. nov. - Species with HL + ML < 2.5 mm; inner border of mandibles without preapical teeth, or if present, in a number smaller than 10..............2 2 Petiolar node in frontal view with apical margin rounded, smooth and thin, or slightly emarginated in the middle, but in this case the corners are always broadly rounded.................. ...............................................altisquamis Mayr - Petiolar node in frontal view with apical margin distinctly concave, the 2 free corners forming angles or produced as teeth.............................3 3 Small species, HL+ML < 1.75 mm; the second segment of antennal funniculus less than twice as long as broad........................neglectus Emery - Larger species, HL+ML>1.75 mm; the second segment of antennal funniculus at least twice as long as broad..............................diegensis Forel Anochetus miserabilis González-Campero & Elizalde, sp. nov. Material examined: Holotype Worker. Argentina. Chaco province. Reserva El Loro Hablador (25º 28’ 51.36”S, 61º 54’ 44.59”W – 163 m a.s.l.), 10.xi.2004, L. Elizalde col., IFML. Paratypes: Argentina. Chaco province. Parque Nacional Chaco (26º 49’ 40.42”S 59º 41’ 1.94” W – 67 m a.s.l.), v.2004, L. Elizalde col., 1 worker and 2 queens IFML; Reserva El Loro Hablador (25º 30’ 14.85”S 61º 54’ 44.26” W – 163 m a.s.l.), 10.xi.2004, L. Elizalde col., 2 workers, IFML. Formosa province. Parque Nacional Pilcomayo (25º 07’ 15.93”S 58º 10’ 28.93”W – 72 m a.s.l.), v.2004, L. Elizalde col., 1 worker, IFML. Paraguay. Alto Paraguay department. Parque Nacional Defensores del Chaco, (20º 33’ 3.12”S 59º 50’ 18.85”W – 127 m a.s.l.), 12.vi.2003, L. Elizalde col., 3 workers, IFML. Etymology: miserabilis is derived from the Latin word “miserable” and it refers to the behavior of this species of visiting the refuse dump of the ant Acromyrmex hispidus. Diagnosis (worker): HL + ML > 2.5 mm; mandibles with 3 apical teeth and 10 -12 preapical denticles; pronotum and propodeum finely punctate; anterior and posterior faces of the petiole smooth and shining, without sculpture. Worker measurements: Holotype (Paratypes n = 7), HL 1.87 (1.87-2.02); HW 1.65 (1.65-1.72); ML 1.30 (1.12-1.30); SL 2.00 (1.90-2.00); EL 0.32 (0.32-4.00); CI 88.00 (83.75-89.61); MI 69.33 (59.21-69.33); SI 121.21 (110.14-121.21). Description Head, mesosoma and gaster reddish; legs lighter. Head: In a full-face subrectangular; finely striated longitudinally, parallel rugae running mesad trough at eye level, posteriorly divergent, not reaching occipital corners (Figure 1A); lateral cephalic margin slightly concave and widened at eye level; occipital margin concave; occipital angle rounded and smooth; medial furrow well-marked not reaching vertex; nuchal carina rounded and continuous; eye located laterally on anterior half of head; mid ocellus present and well-developed (except in the samples from Paraguay); scape smooth and shining, projecting beyond posterior head margin more than 1/8 its length; length of: flagellomers 1-4 and 6-10 twice its width, flagellomere 5 three times its width and flagelomere 11 is 4 o 5 times its width; mandible linear, without basal angle; inner mandibular margin with 3 long sensorial hairs (length > 0.75 mm), 12 preapical denticles and 3 apical teeth of which the central tooth is always shortest. MESOSOMA: metanotal suture deeply concave in profile (Figure 1B); metanotal area with longitudinal grooves in dorsal view; pronotum and propodeum finely punctate with no erect hairs; mesonotum transversally striated; posterior dorsal angle of propodeum well-marked without tooth or spine; mesopleuron with weak sculpture in upper half, smooth and shining in lower half (Figure 1 B); metapleural lobe well-defined slightly projecting over metacoxa; propodeal declivity straight in lateral view; in posterior view with weak transverse striation, propodeal spiracle elongated, its length twice its width; coxae and tibiae smooth and shining; pro and metatibia with 2 apical spurs, the outer one being simple with half length of the inner one’s; inner spur pectinated; mesotibia with two small and simple apical spurs of about 0.05 mm; procoxa with 5 (3-8 in paratypes) scattered erect hairs of > 0.1 mm; meso and metacoxas with 3 (2-5 in paratypes) erect hairs of > 0.1 mm. PETIOLE: with node higher than propodeum, apex bicuspid (Figure 1B), anterior face forming angle of 45º with main axis of the petiole, posterior face convex; anterior and posterior faces smooth and shining; subpetiolar process projects as well developed lobe anteriorly. GASTER: Tergite I smooth and shining, without hairs, posteriorly following terga each smooth with 4 or more scattered erect hairs of more than 0.15 mm. Every sternite with 4 or more scattered erect hairs longer than 0.15 mm. Queen measurements: 2 queens. HL: 1.92 (1.92); HW: 1.72 (1.75); ML: 1.25 (1.20); SL: 1.90 (1.90); EL: 0.40 (0.42). Similar to worker in color of head, legs and body. HEAD: in dorsal view with fine longitudinal striation; in lateral view smooth and shining; 3 ocelli present, antennae with 12 segments; length of all flagellomeres approximately 2.5 times their width; well-developed compound eyes located on the anterior half (Figure 2A). MESOSOMA: pronotum rugose, mesonotum longitudinally striate, mesopleuron with weak sculpture in dorsal half, smooth and shining on ventral half; propodeum rough; smooth and shining coxa; procoxa with 5 scattered erect hairs of more than 0.1 mm; meso and metacoxa with 3 erect hairs of more than 0.1 mm; pro and metatibia with 2 apical spurs, outer one simple, its length half the width of inner one which is pectinated; mesotibia with 2 simple apical spurs; anterior wing with Radial, Medial and CuA closed; medial and cubital veins do not reach the wing margin; r-m3 and cu-a veins, incomplete leaving Cu1 and Cu2 cells open (Figure 2 B). PETIOLE: similar to worker. GASTER: tergite I smooth with no hairs; tergite II, slightly striate with no hairs; from terguite III posterad, smooth with 6 or more scattered erect hairs of more than 0.15 mm. Every sternite with 4 or more scattered erect hairs of more than 0.15 mm Male: unknown. Biology Workers of A. miserabilis were collected associated with Acromyrmex hispidus Santschi (Formicidae: Myrmicinae: Attini) in Chacoan region of Paraguay and Argentina. The Gran Chaco covers over 1.000.000 square kilometers (386.000 square miles) of Argentina, Bolivia, Brasil and Paraguay. Argentina holds 60 percent of this region with a variety of habitats that includes humid and dry forests, wetlands, gallery forest and savannahs. Workers of A. miserabilis were frequently found walking around refuse dumps of A. hispidus. These refuse dumps are located about 50 centimeters from the mound of the attine nest and communicate with it through underground paths, also used by Anochetus miserabilis. Workers of both species were totally indifferent to each other. Generally, refuse dumps constitute a significant source of organic matter that serve as food for many insects which, in turn, could be used as a prey by Anochetus miserabilis. The workers of A. miserabilis were seen hunting cricket nymphs and termites, both in the foraging trails of the attines and in the refuse dump itself. Most observed nests of this leaf cutter species, which is very abundant in the Chaco, presented Anochetus associated with them. On one ocassion, when breaking an upper part of a mound of Acromyrmex hispidus, workers and winged queens of A. miserabilis were found but no brood. No nest of A. miserablis was found nearby. Discussion Anochetus miserabilis is easy to distinguish from the three species cited for Argentina (Anochetus neglectus, A. altisquamis and A. diegensis), due to its longer head (CI: 88.00). In addition, A. diegensis differs from the new species in presenting erect hairs on the mesosoma and mandibles with a single preapical tooth close to the apical teeth. If we passed A. miserabilis through the key of Brown (1978), we arrived at couplet 5, the emarginatus group, nevertheless it does not fit any of those species. A. miserabilis is placed within this group (Brown 1978) which includes neotropical species of large size and slender mesosoma; with mandibles serially dentate; petiolar node short or long, bicuspidate or bidentate. The emarginatus group was distributed in northern South America and the Antilles previous to this study. With the inclusion of Anochetus miserablis in the emarginatus group, the range of this group is extended to the south, towards Argentina and Paraguay. The emarginatus complex includes 4 species (Brown 1978): - A. testaceus (Forel): thorax ferruginous yellow; almost all the vertex, the pronotum and the half upper part of the petiole are smooth and bright; petiolar tooth, not very sharp. - A. emarginatus (Fabricius): thorax and petiole ferruginous to dark color; legs and head, pale yellow; pronotum variable, rough or transversally grooved to smooth and bright; frontface of the petiole, generally rough and opaque; petiolar tooth short. - A. striatulus (Emery): body dark reddish brown, including the head (head angles slightly pale); legs yellowish brown; pronotal disc and petiolar node finely grooved; posterior center of the pronotal disc smooth and bright; petiolar tooth sharp and long. - A. micans (Forel): body light ferruginous; pronotum striated at least in the front anterior half; the sculpture and the petiolar teeth varying with the locality. Analyzing local variations in Costa Rica, Longino (2006) proposed that A. micans and A. striatulus are probably synomyms of A. emarginatus. Anochetus miserabilis differs from A. emarginatus, A. striatulus and A. micans by having a completely smooth and shining petiole. A. testaceus has the pronotum smooth and shining, while Anochetus miserabilis has a finely punctate pronotum. According to Brown (1978), A. emarginatus varies in color and sculpture with the locality where it was collected, although the color of its head is always lighter than that of the mesosoma and gaster, unlike Anochetus miserabilis that presents a uniform color throughout its range. This constitutes the first record of an association between species of Anochetus and leaf cutter ants, and to our knowledge, the first association between a Ponerinae and an Acromyrmex. It is possible that this association is a case of parabiosis, where two ant species can live in the same nest and can share the foraging trails, but they keep the brood separated (Höldobler et al.1990). There is a parabiotic association between species of Ponerinae ants, Odontomachus mayi and Pachycondyla goeldii, and the garden ants Crematogaster (Orivel et al. 1997). In that study there was no interspecific aggression between the ants, and the authors concluded that the tolerance was not explained by a change in the chemical profile of the cuticular compounds. The new association found between Anochetus and Acromyrmex, could be an interesting system to study how these interspecific tolerances are reached. Acknowledgements We thank Fabiana Cuezzo for her valuable comments on an early version of the manuscript. This research was supported by a grant from CONICET to L.E. and FONCYT PICT 12605 program of M.C.G.G References
© 2008, Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em08004f1.jpg] [em08004f2.jpg] |
| |||||||||
{kind=link}
{kind=link}