 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
International Journal of Environmental Research, Vol. 3, No. 1, Winter, 2009, pp. 35-44 The Physiological Ecology of Cardamom (Elettaria cardamomum M) in Cardamom Agroforestry System Murugan, M.1*, Shetty, P. K.1, Ravi , R.2 and Subbiah, A.3 1 School of Natural Sciences and Engineering, National Institute of Advanced Studies,
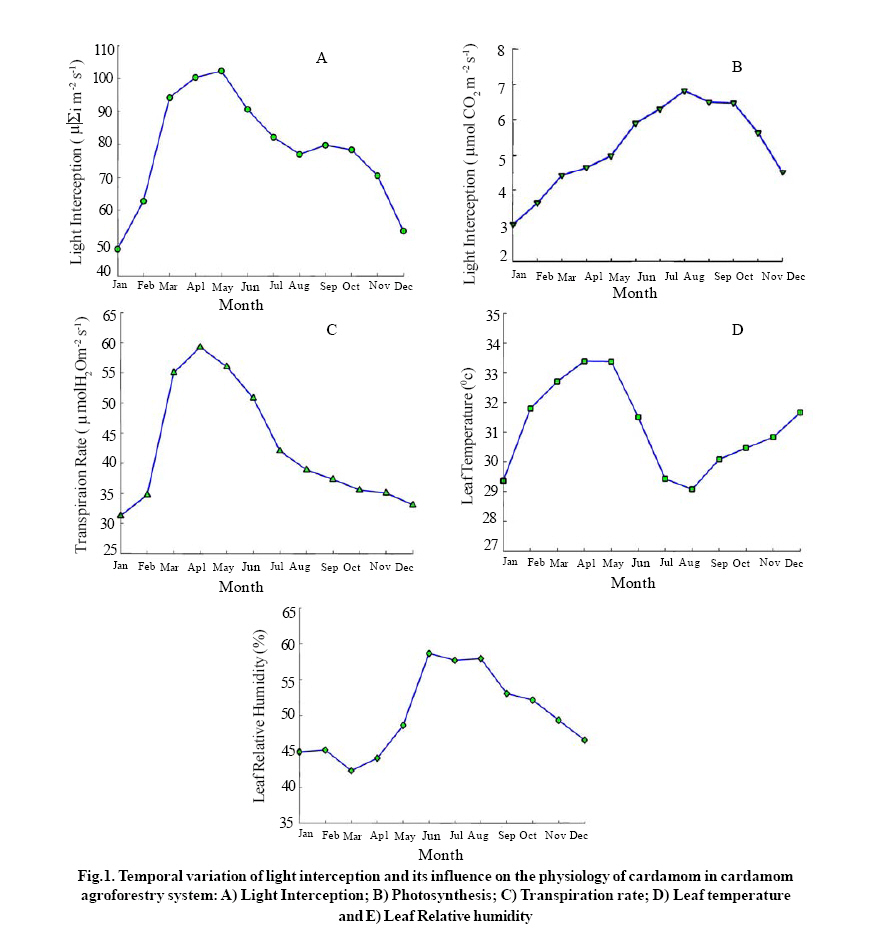
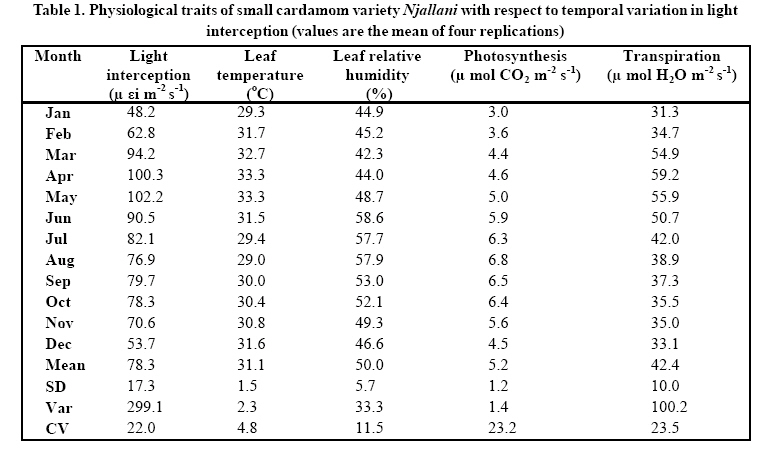
Indian Institute of Science campus, Bangalore-560 012, India Received 14 June 2008; Revised 15 Sep. 2008; Accepted 25 Sep. 2008 Code Number: er09004 ABSTRACT Since 1895 cardamom has been cultivated in the cardamom hills of Western Ghats, India, which form a part of global biodiversity hot spots. These tropical forests in the last couple of decades have been subjected for severe periodical shade lopping and selective felling for maximizing cardamom production. Photosynthesis, transpiration, leaf temperature and leaf relative humidity was studied in response to change in intercepted light condition in each month during the year 2007. The light interception significantly varied from one month to another and increasing trend was noticed from January ( close to 48 μ εi m-2 s-1) up to the month of May ( close to100 μ εi m-2 s-1 ) and thereafter decreasing trend followed until December (50 μ εi m-2 s-1 ). Increased light interception increased photosynthesis, transpiration and leaf temperature. Photosynthesis rate was maximum (close to 7 μ mol CO2 m -2 s-1) in the month of August in which month the interception of light was 77 μ εi m-2 s-1. Increasing light interception beyond 77 μ εi m-2s-1 did not increase photosynthetic rate of the crop. The highest transpiration rate of 59 μ mol H2O m-2 s-1 was observed when the light interception and leaf temperatures were 100 μ εi m-2 s-1, 33.4 ºC respectively. Summer months have recorded higher transpiration rate than those of monsoon months. Results showed good correlation between light interception and the rates of photosynthesis, transpiration, as well as leaf temperature. Key words: Light interception, Photosynthesis, Transpiration, Leaf temperature, Correlation, Cardamom INTRODUCTION Small cardamom (Elettaria cardamomum M) (hereafter called cardamom) is one of the oldest spices known to mankind. The southern most part (Agasthiya malai-Kalakkad Mundanthurai range) of the Western Ghats, India is considered the home of this crop. Originally found as shade loving understory crop of herbaceous non-woody shrub in the erstwhile tropical rainforest (present cardamom hills, known as cardamom hill reserve or CHR) (Wilkinson and Elevitch, 2000). Since last 120 years, cardamom has been cultivated in cardamom hills where sun loving crops are not allowed so as to preserve cardamom ecosystem environment. The practice of shade lopping (cutting branches of natural forest trees) has been continued primarily for the purpose of allowing more sunlight to minimize the pathogenicity of disease causing organisms during the rainy seasons of a year. Until 1980s shade regulation was not widespread among planters (cardamom farmers). The positive response of cardamom (higher yield) to increased levels of sunlight was found out later on. Therefore now, majority of the planters believe that increasing the frequency of shade lopping had increased the crop yields of small cardamom.This has led the way for regular shade lopping and now the entire cardamom hill reserves maintain a shade level ranging at55-60%. After the advent of high yielding varieties of cardamom such as Njallani, Palakkodi shade lopping becomes regular and widespread owing to their good performance under more open condition. Shade lopping which always ends in degradation of tropical forest can aggravate local warming in cardamom agroforestry system therefore ecosystem climate change is inevitable. If the fluctuation of precipitation pattern and increase in hours of sun light and clear skies by degradation of mesoscale forest and local land use change, there would be possibility that cardamom plants are repeatedly exposed to stress and thereby sustainability of cardamom agroforestry can be highly questionable in near future. Systematic studies on the physiology of small cardamom which is the fundamental process for economic yield have been lacking. This kind of study is more relevant and essential under current and future climatic change scenario of cardamom agroforestry system. The frequency of extreme weather will increase in cardamom agroforestry system (Murugan et al., 2008 a ). Light intensity can be crucial for the yield of understorey crop plants in tropical mountain ecosystem (Mooney et al., 1980). The knowledge of the physiological characteristics of understorey crop plants is essential to our understanding of the functioning and effective management of agroforestry systems. The tropical humid forest systems like cardamom agroforestry system, is one of the most productive systems in terms of monetary returns in the world. Considering the role of agriculture in the social and economic progress of developing countries, the vulnerability of agricultural systems to the impacts of climate change has received considerable attention from the scientific community (Fisher et al., 2002; Kurukulasurya and Rosenthal, 2003). Much of the available literature confirms that the overall impacts of climate change on agriculture especially in the tropics will be highly negative, particularly in the mountain agroecosystems. The change in macro and microclimate in cardamom hills has been studied and it was evident that the warming is unequivocal and rainfall pattern is much altered since 1960 (Murugan et al., 2000; 2003). The practice of shade regulation in tropical perennial agroforestry systems (cardamom, coffee, vanilla and cacao) has been practiced since last many decades but the intensity of shade lopping varied with crops and places. Within cultivation of small cardamom, the lopping of branches is regular (once in six months to one year) in Indian cardamom hills while two to three years in Srilanka. Therefore the much more complex tropical forest has been converted to a more open, simple and uniform system and hence leading to degradation of cardamom agroforestry system in India. As a whole, shade can create microclimates with lower seasonal means in ambient temperature and solar radiation. Beer et al., (1998) have observed that shade trees buffer high and low temperature extremes by as much as 5ºC in coffee and cacao plantations. Selective felling of trees in all agroforestry systems has also been noticed for various reasons which can increase soil temperature by 4ºC and reduce relative air humidity by 12% in coffee and cacao plantations. Agroforestry systems can bring about favorable changes in the microclimatic conditions by influencing radiation flux, air temperature and wind speed all of which will have a significant impact on modifying the rate and duration of photosynthesis, transpiration and stomatal conductance (Monteith et al.,1991). However, the response of crops to varied climatic condition induced by shade level depends on the variety and type of crops. The net shade effect was reported to be more positive for C3 crops which are normally light saturated in the open (Ong, 1996). Much of research, to date, has focused on the impacts of shade and light intensity on annual crops and very little on perennial agroforestry systems which are most unique systems in the world. Although the total stomatal pore area may be only 5% of a leaf surface (Willmer and Fricker, 1996), the rate of water vapor loss may reach as high as 70%. Stomata exert major controls on both the water and water cycles of the world. Annual precipitation over the land (annual average of 1023 mm) is about 110,000 Km3 or 110×10 15 kg and evaporation and transpiration total about 70×10 15 kg. The greatest rates of transpiration occur in the uniform and warm forested areas between the tropics with 32×10 15 kg yr-1 of water vapor passing through stomata. This is double the water content of the atmosphere (15×10 15 kg yr-1) (Hetherington and Woodward, 2003). Plants with higher stomatal conductance show minimal gain of photosynthesis and minimal changes in leaf temperature for large changes in conductance, reflecting high water usage and little limitation on water supply. By contrast plants with low values of stomatal conductance may occur in water limited environments, where small reductions in stomatal conductance significantly increase water use efficiency. Differing capacities and resource availability (light), forest species have low actual and inherent capacity. There seems to be as great a range of variability in photosynthetic rates among tropical communities. This is probably due to dramatic range of light resource changes. Information on the response of cardamom to more open condition (more light condition) and the interrelationships among the various physiological parameters such as photosynthesis, transpiration, stomatal conductance, leaf temperature and relative humidity etc. have not been studied and quantified for cardamom agro-forestry system. Therefore, the primary objective of this work was to study the temporal variation of light interception and its effects on the physiological characteristics of cardamom in cardamom agroforestry system. Under the fast changing cardamom hill reserve climate such ecophysiological studies would be of immense use to manage and to safe guard the cardamom agroforestry system in a rational way. MATERIALS & METHODS Cardamom belongs to zingiberaceae and has C3 photosynthetic pathway. The crop is a shrub by habitat and has several tillers consisting of pseudo stems with leaves on the upper part. The inflorescence appears on the rhizome from the point where pseudo stem shoots up. At present, the variety Njallani has been occupying more than 80% of the total cultivated area in the cardamom hills, Kerala owing to its positive response to natural resources like light and water as well as applied plant nutrients. Observations on physiological parameters were made from index leaf (4th leaf from the top) of yielding plants (2-3 year old) at ten different sites in cardamom hot spots of A (Vandenmedu) and B (Pampadumpara) on monthly intervals during the year 2007. All the experimental sites have had a frequency of shade lopping once in a year maintained. Shade lopping was carried out in the month of May and shade level at the time of lopping was nearly 50%. The hot spots “A” and “B” are the highest productivity areas of cardamom in the world. The experiment plants in all the sites were treated uniformly as per the standard practices of Kerala Agricultural University. No chemical spray or fertilizer application to enhance photosynthetic capacity of the crop has been given at least one week prior to our measurement. All of the observations were taken on clear days between 12 noon to 2.30 pm and same leaves were used for all the physiological parameters observed in this experiment. The data of each parameter given in the (Table.1). are the averages of three replications and the total number of observation sites was ten. The cardamom hill reserve (334 sq km) is surrounded by three national parks namely; the Periyar tiger reserve (PTR) on the south, the Eravikolum national park (ENP) on the north and the Mathikettan national park (MNP) on the eastern side. The river Periyar forms western boundry. The climate is of tropical humid type. The annual rainfall fluctuates from 1670-3000 mm year-1 and annual mean temperature is around 23 C. The altitude of cardamom hills varies from 900-1250 mean sea level.Rates of photosynthesis and transpiration in the fully matured dark green leaf (4th leaf from the top which is considered as reference or index leaf for cardamom) of a tiller in a clump were measured in each site using a photosynthesis measurement system (LICOR, Inc., Steady State Porometer, Mini PAM (Photosynthesis Analysis Meter)). The measurements were replicated thrice for all experiments under this study. Per cent interception of light in the canopy was calculated (middle point of a slope) by comparison with a Photometer (Luxor USA) placed in the open field situation. The measurement was made between mid-day and 2.00 pm as per the method suggested by Nelliat et al., (1974).
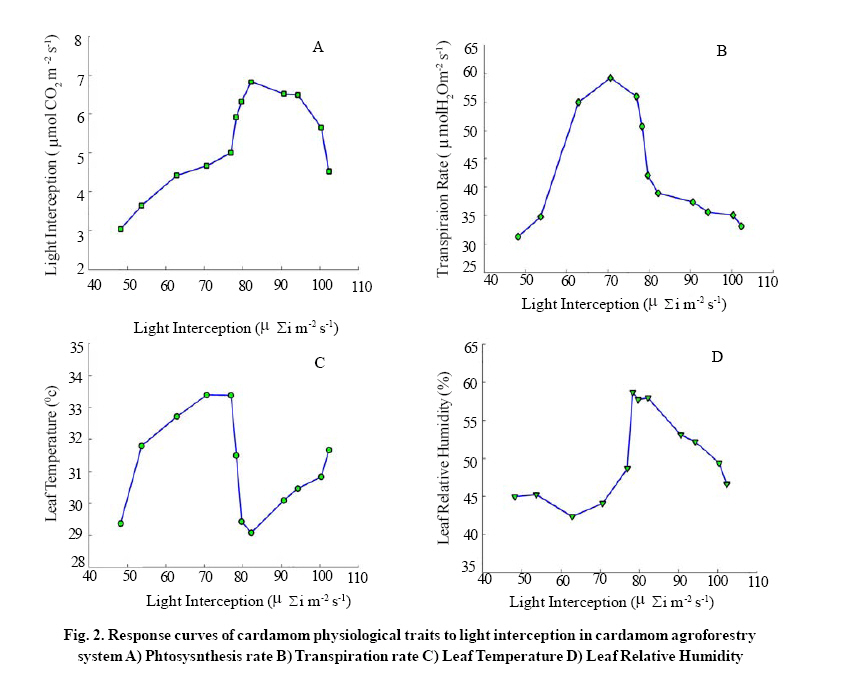
Leaf temperature was measured from the same leaf used for photosynthesis using a thermocouple fixed in the Steady State Porometer LI-1600, (LICOR, Inc., Nebraska, USA) and expressed in °C. The relative humidity inside the cuvette containing leaf sample was estimated using a Steady State Porometer (LI-1600, LICOR, Inc., Nebraska, USA) and expressed in percentage. The transpiration loss of water was assessed using a Steady State Porometer (LICOR, Inc., Nebraska, USA) and expressed in μg H2O. The quantum yield of photochemical energy conversion was estimated using the photosynthesis analysis meter (MINI-PAM). The photosynthesis yield was expressed in terms of chlorophyll fluorescence (Fv / Fm). The resistance offered by stomata for the exit of water from the leaf surface was measured using a Steady State Porometer (LICOR, Inc., Nebraska, USA) and expressed in s cm-1.Statistical analysis was performed for each parameter studied using MATLAB 7.0. Associations among physiological characters were examined by simple correlation analysis and the correlation coefficient values between variables are given in (Table 2). The response curves shown in (Fig.1) were prepared by sorting the light interception values against the other variables. RESULTS & DISCUSSION Light interception in the cardamom canopy varied significantly between months reaching maximum in the month of May (102.2με im-2s-1) thereafter declined gradually up to the month of December (53.7 μ εi m-2 s-1 ), January recorded the lowest value of 48.2 μ εi m-2 s-1 . Table 1 shows the temporal variation in light interception by cardamom plants during a year. The change in light interception could be attributed to: a) cardamom canopy architecture and specific leaf area b) rate of rejuvenation of forest trees c) level of current year shade lopping; the interception of light for September (79.7 μ εi m-2 s-1 ) and October (78.3 μ εi m-2 s-1 ) is well above than for August (76.9 μ εi m-2 s-1 ). These result suggest that the light interception during September and October could be purely seasonal than any other reasons like denseness of forest canopy. Usually shade lopping is done during the months of April and May. This brings about more sunlight falling on the cardamom canopy, during these two months the number of matured tillers as well as leaves per clump is also high leading to more interception of light by cardamom canopy. Forest trees are allowed to rejuvenate till next season comes and this gradual regeneration of forest canopy blocks incoming radiation thereby decreases light interception. The variation in light interception between any two months was found to be 22% (Table 1). Good correlation has been noticed between light interception and transpiration rate (R = 0.91, P=0.05) as well as leaf temperature (R = 0.56, P=0.05) (Table 2; Fig.2B and 2 C). Originally shade lopping was done in anticipation that it would suppress the intensity of rot diseases (capsule rot, sheath rot and clump rot), in our observation despite the recent severe shade lopping (which eliminated natural canopy shapes of forest trees and now all trees have only small crown with tall bole shape), the intensities of these diseases have gone up and to manage these diseases alone planters go for at least 4-6 sprays of fungicides in a year. After the severe modification of shade in cardamom hills, some of the insect pest population (root grubs, white flies and thrips) has been increased many folds and now planters take up minimum 16-20 rounds of chemical spray to manage these pests. Up to the period of 1995 only 6-7 spays of pesticides were in recommendation to suppress most of the cardamom pests and diseases. Therefore the speculation and trade-offs of shade lopping in cardamom hills have been increasing and it needs thorough investigation in terms of agroecological perspectives. The change in leaf temperature recorded for the months March, April and was significant than for the rest of the months. Monthly mean leaf temperature was 31.1ºC and the summer months have recorded higher leaf temperature. Maximum and minimum leaf temperatures of 33.3ºC and 29.0ºC were reported respectively for April and August. The trend of temporal variations of leaf temperature followed similar to that of canopy light interception. The leaf temperature extremes (higher leaf temperature of 33.3ºC) have had good correlation with transpiration (R = 0.72, P=0.05) than leaf relative humidity and leaf photosynthesis (Table 2; Fig.2A and 2 D). The response curve of cardamom leaf temperature for various light interception levels is shown in Fig.2. The lowest leaf temperature (29.0ºC) in the month of August resulted in maximum photosynthesis. These results suggest that leaf temperature have greater role in transpiration and photosynthesis which are fundamental for crop growth and yield. The conditions of humidity at leaf surfaces are of importance for managing cardamom agroforestry system because cardamom is extremely sensitive to microclimatic change which in turn favors development of disease causing organisms. The temporal fluctuation in leaf relative humidity is given in Table 1. The response curve of cardamom leaf relative humidity for various light interception levels is shown in Fig.2D. Mean monthly leaf relative humidity is reported to be 50.0%. March and June are the respective months for the lowest (42.3%) and highest (58.6%) values. Good negative correlation was observed between leaf relative humidity and leaf temperature (Table 2). However in mountain ecosystems the surrounding atmospheric air temperature and air movement will decide the leaf relative humidity and leaf temperature. Higher positive correlation was observed between leaf relative humidity with photosynthesis (R = 0.83, P=0.05). The photosynthetic rate ranged from 3.0 μ mol CO2 m -2 s-1 (January) to 6.8 μ mol CO2 m -2 s-1 (August) and the mean monthly rate was found to be 5.2 μ mol CO2 m -2 s-1 (Table 1). The response curve of cardamom photosynthetic rate for various light interception levels is shown in Fig.2A. The rate of photosynthesis increased with passage of months up to August then declined gradually up to December. When the light interception was minimum in January (48.2 μ mol CO2 m -2 s-1) the photosynthesis was the lowest (3.0 μ mol CO2 m 2 s-1). Photosynthesis increased with increase in light interception up to 76.9 μ εi m-2 s-1 (optimum light interception level for cardamom). Further increase in light interception to 102.2 μ εi m-2 s-1 did not increase the photosynthetic rate but reduced it to 5.0 μ mol CO2 m -2 s-1. These results give us exact information on light saturated photosynthetic rate of cardamom. Maintaining shade level at which the cardamom canopy light interception is near optimum (76.9 μ εi m-2 s-1) level could be beneficial for cardamom agroforestry system. This has remarkable significance and implication on shade lopping and regulation in cardamom agroforestry system. Severe and frequent shade lopping to allow more sunlight to fall on the cardamom canopy to get more yields will not increase the photosynthetic rate. High photosynthetic rate observed during first (June, July, August and September) and second monsoon (October, November and December) months can maximize growth and yield of cardamom. That is the reason why the first and second monsoon months harvests are the peak harvest for cardamom. Therefore the high rates of photosynthesis during first and second monsoon months can be ascribed to photosynthetic source- sink relationship. Surprisingly after the peak harvest (third harvest), the sink capacity becomes weaker which influences the photosynthetic capacity to be slow down, and the impact of which has been reflected as reduced per capsule weight in the subsequent harvests. In majority of the rain fed cardamom plants the photosynthetic sink (cardamom capsules) is absent during summer months. Similar observation in other crops has been reported elsewhere (Stanghellini and Bunce, 1994; Stitt, 1991). A range of variability in photosynthetic rates among members of tropical plant communities under light limiting condition has been reported by Mooney, et al., (1980). Paradoxically, the lower photosynthetic rate has negative implication on the yield of cardamom; the combined effect of maximum day time temperature and lower photosynthetic rate can be one of the causes of immature capsule shedding during summer months. Higher atmospheric temperature exceeding 34 ºC (day time maxima) observed in the month of April 2008 resulted in heavy shedding of immature capsules in majority of rain fed plantations has been evident from many plantations. From these findings it is understood that lower photosynthetic rates would have negative impact on yield of cardamom. The transpiration rate of cardamom in response to temporal variation in light interception is showed in Table 1. Transpiration rate and its relationship with other physiological parameters are given in Table 2. Leaf temperatures have positive correlation with transpiration rate (R = 0.72, P=0.05). The response curve of cardamom transpiration rate for various light interception levels is shown in Fig.2B. Transpiration rate was lowest (31.3 μ mol H2O m-2 s-1) in January and reached maximum (59.2 μ mol H2O m-2 s-1) during April. Thereafter the rate of transpiration decreased gradually up to December (33.1 μ mol H2O m-2 s-1). Higher level of transpiration rate has been evinced during summer months (March, April, May and June).The probable reasons for this observation could be the higher level of light interception and leaf temperature during the summer month’s period. If transpiration rate is directly dependant on leaf temperature and leaf temperature is much influenced by surrounding air temperature, then the transpiration rate can be much higher under future warming condition. Therefore the crop demand for water during summer months can exceed stock level. This can lead to reduced yield of cardamom thereby sustainability becomes unattainable. Stomatal diffusive resistance was maximum (6.1 s cm-1 ) when the transpiration rate was also maximum (7.1 μg of H2O cm-1 s-1). No correlation could be established with chlorophyll fluorescence against stomatal diffusive resistance and transpiration rate in cardamom (data not given in this paper). These findings will have significance in drawing implications on the sustainability of cardamom agroforestry system under fast changing weather and climatic condition. CONCLUSION The paucity and need of information on physiological ecology of cardamom is of great concern in view of the rate of destruction of this peculiar forest ecosystem (cardamom hill reserves) in Western Ghats, India. This paper touched upon the study of canopy light interception in cardamom agroforestry system which is essential for managing the crop growth and productivity as well as effective management of CHR. The influence of amount of light intercepted by cardamom clumps on various physiological parameters has been discussed. During the months (April and May) of severe shade lopping the yield of cardamom was decreased considerably in many rain fed plantations; in addition to this, the CHR had also experienced multiple trade-offs e.g. soil erosion and soil moisture regime (Mathew et al., 2004), loss of bio diversity and microclimate change (Murugan et al., 2008 a ), increased intensity of insect pests and disease attack , frequent application of pesticides (Shetty et al., 2007), and soil quality and soil nutrient dynamics. As a whole shifts in ecosystem dynamics of CHR have occurred because of agricultural modification of forest shade lopping, and this necessitates further scientific investigation on ecosystem resilience. Agricultural related regime shifts in sensitive ecosystems like CHR is important in present context when agriculture is treated as not only production base but also provider of environmental services as well (Gordon et al., 2008 a ). Higher transpiration rates reported in this paper for the summer months can have negative implications on yield of cardamom and this could be attributed to enhanced incident light intensity and leaf temperature. Increase in transpiration rate can decrease the yield of cardamom significantly during summer months (April and May) for example in our observation, the number of flowers and capsules formed as well as the percentage of capsule set in per plant (30% in summer and 80% in August or September) had been decreased significantly during summer months. Synchronous warming (both soil and atmospheric air warming) in cardamom hills (Murugan et al.,2008 b )is evident which can adversely impact soil nutrient dynamics particularly major nutrients (nitrogen, phosphorus and potassium) thereby yield reduction can be unavoidable. Therefore sustainability of climate sensitive cardamom agroforestry system could be in danger when future climate is more unstable and uncertain as predicted by models. Although the light interception of cardamom clumps depends on the level of shade lopping, it is understood that more efficient canopy architecture is essential to avoid self shading of cardamom. Self shading is one of the main barriers of yield improvement. Therefore increasing the frequency and severity of shade lopping from the current rate may not increase the photosynthetic capacity of cardamom and thereby may not increase yield; instead it would decrease it and invite ecological risks also. Development of high yielding cardamom varieties and types which can intercept more light (under optimum shade level of 55-60%) with high water use efficiency would be of good strategy to manage the cardamom agroforestry system in a rational way. In the end, present knowledge level on physiological ecology of cardamom is so limited and this information will be of immense use for CHR system indeed. ACKNOWLEDGEMENTS This paper forms a part of work done by the senior author Muthusamy Murugan towards his doctoral research programme in agroecology. The work was supported by National Institute of Advanced Studies (NIAS), Bangalore in the form of senior research fellowship. The authors express their sincere thanks and gratitude to NIAS. The anonymous reviewers were sincerely thanked for their insight review comments. REFERENCES
© Copyright 2009 - Graduate Faculty of Environment University of Tehran The following images related to this document are available:Photo images[er09004t2.jpg] [er09004t1.jpg] [er09004f2.jpg] [er09004f1.jpg] |
| |||||||||


{kind=link}
{kind=link}
{kind=link}
{kind=link}