 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
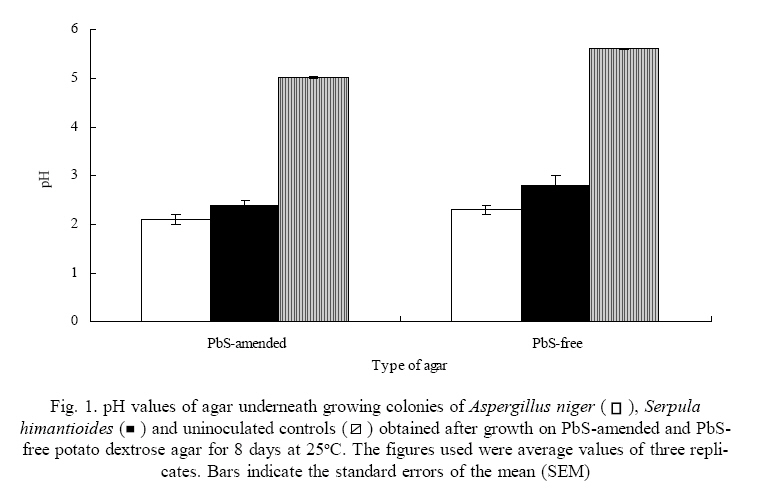
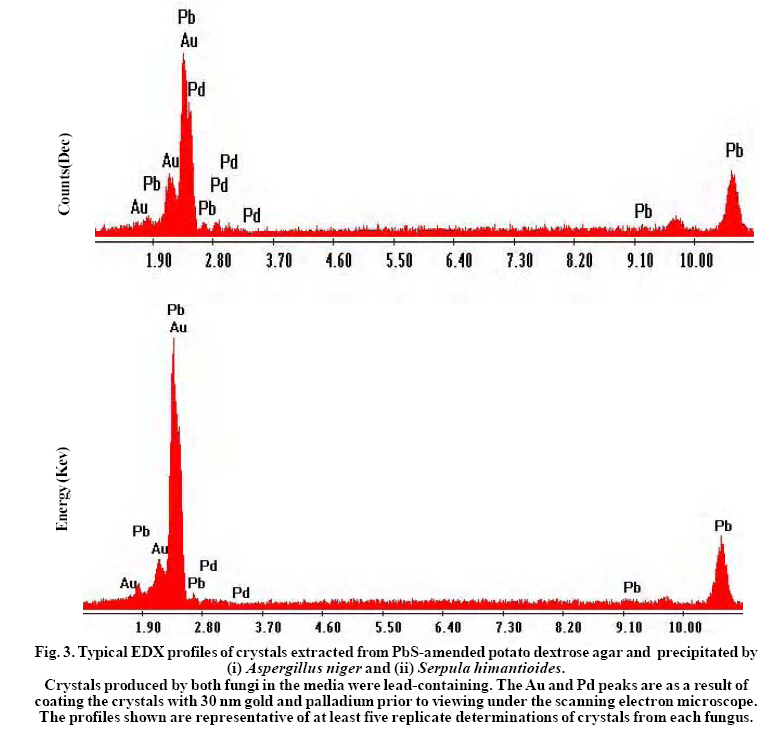
International Journal of Environmental Research, Vol. 3, No. 4, 2009, pp. 477-482 Biological Immobilization of lead from lead sulphide by Aspergillus niger and Serpula himantioides Adeyemi, A.O. Division of Environmental and Applied Biology, College of Life Sciences, University of Dundee, Dundee, DD1 4HN, Scotland, UK and Present address: P. O. Box 83034, Vista Centre, 1830 Bank Street, Ottawa, K1V 1A3, Ontario, Canada *Corresponding author E-mail: dekov@hotmail.com Received 12 April 2009; Revised 17 July 2009; Accepted 27 July 2009 Code Number: er09052 ABSTRACT The abilities of Aspergillus niger and Serpula himantioides to immobilize lead (Pb) from lead sulphide (PbS) were investigated. Upon growth on potato dextrose agar (PDA) amended with PbS which contained 10mM Pb, no visible solubilization activity was detected. A. niger produced biomass in greater quantity compared to S. himantiodes in both experimental and control media. With a ratio of 1.13, A. niger growth was stimulated on PbS-amended media while that of S. himantiodes, with a growth ratio of 0.46 was inhibited. Despite its smaller biomass, S. himantiodes accumulated over five times more Pb than A. niger with 85.7 ± 4.9 µmol of Pb per g of dry weight while A. niger contained 14.6 ± 1.3 µmol of Pb per g of its dry weight. Both fungi immobilized Pb through the formation of crystalline precipitates and energy dispersive x-ray analysis revealed the precipitates to be Pb containing. This transformation is of importance in the weathering of metalbearing rocks as mineral neogenesis. Key words: Galena, Lead, Aspergillus niger, Serpula himantioides, Immobilization INTRODUCTIONLead and sulphur are the elemental constituents of the mineral known as galena (lead sulphide – PbS). Lead (Pb) is a well known toxic metal which is common in the soils and rocks of the earth and deposits usually occur in the form of galena (Greninger et al., 1975; Blaskett and Boxal, 1990; Davies, 1995). Its widespread occurrence in the environment, especially in soils comes from anthropogenic sources such as metal smelting industrial emissions, vehicular exhaust gases and weapons testing in military training (Alloway, 1995; Davies, 1995; Buatier et al., 2001; Cao et al., 2003; Shahidi Bonjar, 2007). Fungi are capable of growing in many environments and the survival and growth of many species is possible in high toxic and metal-laden environments. Several varied mechanisms exist by which they are able to deal with metals in such environments, e.g. extracellular precipitation of secondary minerals, metal binding to fungal cell walls, intracellular sequestration and complexation, compartmentation or volatilization (Gadd, 1993, 1999). The extracellular precipitation of secondary minerals in particular by fungi, often occurring as a result of oxalate/oxalic acid production which immobilizes metals in the process, is of environmental significance especially regarding survival, bio-deterioration, pathogenesis, soil weathering, mineral formation and metal detoxification (Sayer and Gadd, 1997; Jarosz-Wilkolazka and Gadd, 2003). In this paper, the abilities of two fungi – Aspergillus niger and Serpula himantioides to immobilize Pb from PbS were investigated. MATERIALS & METHODSAspergillus niger (ATCC No. 201373) and Serpula himantioides (obtained from Dr N. White, University of Abertay Dundee, UK) were used for the study. They were grown on Potato dextrose agar (PDA; 39 g l-1, Merck, Darmstadt, Germany) prior to and during the experiment. Experimental PDA was prepared and amended with an amount of commercially available PbS (Sigma Aldrich, St Louis, Missouri, USA) which would give 10mM Pb in the final agar plate if the compound was fully soluble. Precut dialysis membranes discs (Medicell Ltd., London, UK) prepared by boiling twice for 10 minutes in ddH2O and rinsed thoroughly on each occasion were autoclaved at 121oC for 15 minutes and placed under aseptic conditions onto the surface of the agar in each 90mm Petri dish prior to inoculation with 7mm diameter discs of A. niger and S. himantioides cut from the margin of five day old grown mycelia on PDA. The inoculated plates were incubated in the dark at 25oC over an eight day period. Radial growth and that of any clear zones underneath or around the colonies (indicative of solubilization) over this period were measured as the mean of two perpendicular radii of growing colonies and recorded on a daily basis until the mycelia reached the edge of the Petri dish or the experimental period had elapsed. After eight days, fungal biomass from each membrane was harvested into foil cups and oven dried at 70oC to complete dryness. pH values of the agar across the dish at 20 mm intervals were then measured. Lead content in the biomasses of both fungi was determined using a Perkin Elmer AAnalyst 400 Atomic Absorption Spectrophotometer. Biomass samples from each fungus were digested in a heating block at 90oC for twenty hours with 2 ml concentrated HNO3. Known lead standards in a range of concentrations were prepared from a standard solution obtained from BDH, Poole, UK. Digested biomass solutions were analyzed for Pb content after appropriate dilutions with ddH2O. In order to extract the precipitates (i.e. crystals) formed by the fungi, the dialysis membranes upon which fungal mycelia grew on were first removed from the surfaces of the mineral-containing agar at the end of the duration of the experiment. The agar was collected with a spatula into a crystallizing dish and sterile distilled water was added to the dish. The dish was put in a microwave oven at full capacity (950W) for 2-3 minutes until the agar dissolved completely, leaving behind the crystals in the resulting solution. The solution was pipette out of the dish and the process repeated twice. Crystals which settled at the bottom of the dish were then transferred into an Eppendorf tube and placed in desiccators until fully dry crystals were obtained. Samples of the resulting crystals were mounted on double sided carbon adhesive tape on 9 mm diameter aluminum stubs and sputter coated with 30 nm gold (Au) and palladium (Pd). The crystals were viewed under a Philips XL 30 environmental scanning electron microscope (ESEM). Energy dispersive x-ray (EDX) analysis linked to the ESEM was used to determine the constituent element(s) of the crystals. Throughout the 8 days of growth on PbSamended PDA containing 10mM of Pb, growth rates of A. niger increased by 7.08 ± 0.36 mm day-1 while colony growth rate for S. himantiodes increased by 3.46 ± 0.42 mm day-1 (mean values ± standard error of mean; three replicates each). Growth ratios (Rm:Rc), calculated from the rate of growth of each colony on mineral-amended agar (Rm) compared to that of mineral-free agar (Rc) (Sayer, 1996), indicated that the growth of A. niger with a ratio of 1.13 was stimulated by PbS in the medium while the growth of S. himantiodes with a ratio of 0.46 was inhibited (a Rm:Rc value > 1.0 indicated that the mineral compound stimulated fungal growth while a value < 1.0 is indicative of growth inhibition). No visible solubilisation of PbS was observed underneath either of the growing fungal colonies or elsewhere in the agar plates. Although there was no visible solubilisation, lead accumulation by biomass as described below is both an indicator and mechanism of solubilisation (Burgstaller and Schinner, 1993). Another plausible reason for the lack of visible solubilisation is the role of particle surface area and how it affects fungal ability to solubilise minerals. The average particle size of PbS used in this study was ~ 355 µm, rather large when compared to the average particle size of, for example, Ca3(PO4)2 at ~ 90 µm which A. niger, S. himantioides and a range of other unidentified soil fungi solubilized (A.O. Adeyemi unpubl.).The addition of PbS to PDA decreased the pH of the medium from 5.62 ± 0.01 to 5.03 ± 0.03. Growth of both fungi on PbS-amended PDA further reduced the pH of the medium and the results are depicted in Fig. 1. A. niger generally produced biomass in greater quantities compared to S. himantiodes in both experimental and control media. The obtained dry weight values also reflected PbS stimulation of growth for A. niger and inhibition for S. himantioides. This could also be seen in the lead tolerance values, expressed as an index based on the dry weights of both fungal biomasses obtained during the study. The tolerance index of fungal biomass is described as the dry weight of mycelium on amended agar/dry weight of mycelium on control agar, expressed in percentage terms (Vodnik et al., 1998; Colpaert et al., 2000; Fomina et al., 2005). The tolerance index of A. niger was 116 % while that of S. himantiodes was 50 %. There was Pb accumulation in the biomasses of both fungi after 8 days of growth although S. himantioides was more effective than A. niger. Despite its smaller biomass, it accumulated over five times more Pb than A. niger – S.himantioides contained 85.7 ± 4.9 µmol of Pb per g of dry weight while A. niger contained 14.6 ± 1.3 µmol of Pb per g of its dry weight. Pb accumulation by both fungi therefore indicated the possible use of fungal biomass as sorbent material and this may provide some potential for metal removal in contaminated environments. Oxalate-like crystals were precipitated by both fungi and observed in the agar and underneath the growing colonies of both fungal species after 3 days growth with a light microscope (results not shown). After 8 days of growth and when examined under an electron microscope, the morphologies of crystals produced by both fungi were similar and they generally appeared as clusters of several needle-like crystals (see Fig. 2). Energy dispersive x-ray analysis profiles shown in Fig. 3 revealed both sets of crystals to be Pbcontaining. In addition, the spectra of both sets of crystals were identical to those produced by fungi when inoculated on natural galena rock surfaces in a previous experiment (Adeyemi and Gadd, 2005). While the solubilization of other metal bearing minerals by fungi has been proven (Sayer et al., 1997), this report further demonstrates the capability of fungi to immobilize Pb from PbS through the formation of insoluble metal containing oxalate-like precipitates. Immobilization of metals from metal-bearing compounds by fungi sometimes in the form of oxalates or oxalate-like precipitates is often dependent on their abilities to produce organic acids (Cunningham and Kuiack1992; Sayer et al., 1997; Gharieb et al., 1998). These acids, often low molecular weight, for example, citric and oxalic acids interact through the provision of protons and organic anions which sequester metal cations (Altomare et al., 1999; Gadd 2000) to result in mineral transformation. Whether produced in combination or singularly, both protons and organic acids tend to create an acidified environment which often leads to the dissolution of metal-containing compounds, a transformation which is one of the widely acknowledged means of mineral weathering by fungi (Illmer and Schinner 1995; Sayer et al., 1995; Ehrlich 1996; Sayer and Gadd 1997; Gharieb et al., 1998; Fasim et al., 2002; Welch et al., 2002). Depending on which organic acids are formed, this microbial mediated process has potential for metal recovery and reclamation from contaminated solid wastes, soils and low grade ores (Gadd 1999) since citric acid usually results in the release of elements while oxalic acid result in immobilization (Gadd 2000). This work has further demonstrated that immobilization by fungi through the formation of oxalate-like crystal precipitates may play a role in the toleration of high concentrations of toxic metals, as seen with A. niger which despite the presence of 10mM of Pb in agar, showed an increase in growth rate and ratio and lead tolerance index. This is particularly significant with regards to the environmental concerns surrounding Pb toxicity as fungi may be able to mediate conversion of metal-bearing minerals into less toxic forms through immobilization. Furthermore, this process of insoluble oxalate-like crystal precipitate formation may hold immense potential for bioremediation of contaminated soil and groundwater especially as it may lead to a decrease in the bioavailability of toxic species It is also important in the microbial weathering of rock minerals through the promotion of mineral neogenesis. ACKNOWLEDGEMENTThe author is indebted to Geoffrey Gadd for provision of fungal cultures, laboratory space and access to facilities used for this work. REFERENCES
Copyright 2009 - International Journal of Environmental Research The following images related to this document are available:Photo images[er09052f1.jpg] [er09052f3.jpg] [er09052f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}