 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
REVIEW OF THE DEEP-SEA ANGLERFISHES (LOPHIIFORMES: CERATIOIDEI) OF SOUTHERN AFRICA M. Eric Anderson1 and Robin W. Leslie2 Code Number: FB01002 INTRODUCTION The deep-sea anglerfishes of southern Africa were last reviewed in the book Smiths’ Sea Fishes (Pietsch, 1986a), with seven species recorded. P.C. Heemstra (1995 second edition) added two species of the family Linophrynidae previously recorded from off South Africa. Since 1986 three papers have treated anglerfishes from our waters (Balushkin and Trunov, 1988; Bertelsen and Krefft, 1988; Bertelsen and Pietsch, 1996), but we found several unrecorded specimens in collections and other records that were overlooked by Pietsch in his accounts. Here we provide information on a total of 32 deep-sea anglerfish species from our area. Ceratioids are best known for their dark, usually globular bodies, huge mouths, often with fang-like depressible teeth, and luminescent angling lures (a modified dorsal-fin spine in females), all adaptations to life in the deep pelagic zones. Males are obligatory sexual parasites in the families Ceratiidae, Linophrynidae and Neoceratiidae, and facultative parasites in the Caulophrynidae and the oneirodid genus Leptacanthichthys (Pietsch, 1976). Munk & Bertelsen (1983) described the integration of blood vessels and tissues in a male and female linophrynid, concluding that males derive nutrients from the female while attached and mature. Ceratioids have been recorded in all oceans to both subpolar regions, and are classified in 11 families with 35 genera. Ten families are represented in our area, the exception being the monotypic Neoceratiidae. This rare fish (Neoceratias spinifer) has been taken just outside our area (Bertelsen, 1951), and should be expected when its bathypelagic habitat has been adequately sampled. Larval ceratioids are generally epipelagic and descend into the deeper adult habitat during metamorphosis. Bertelsen (1951, 1984) provides descriptions and illustrations of most known larval forms. Little was known about ceratioids until the major oceanographic expeditions of the late nineteenth and twentieth centuries. Landmark papers on these fishes include Regan, 1926 (on the Danish North Atlantic expeditions of the ship DANA), Parr, 1927 (on the American Atlantic expeditions of the PAWNEE), Regan and Trewavas, 1932 (an update of Regan’s 1926 work), Bertelsen, 1951, (a major ichthyological tome featuring descriptions of almost every ceratioid species then known), and Pietsch, 1974, (an osteological study and revision of the largest family, the Oneirodidae). Papers by the latter two workers, often as co-authors, make up the bulk of the systematic literature on ceratioids, much of it cited herein. MATERIALS AND METHODS Methods of measurement generally follow Pietsch (1974), except head depth is greatest depth behind sphenotic spine, except in Melanocetidae where it is from top of sphenotic bone to ventral base of quadrate; postorbital length is distance from posterior margin of eye in a direct line to posterior edge of preopercle or to its middle when it is posteroventral to the orbit; interorbital width is least bony distance ("least outside width between frontals"; Pietsch and Van Duzer, 1980); pectoral base width is from articulation of base of uppermost ray to base of lowermost; gill slit length is fleshy distance with opening often manually adjusted to its most relaxed position; caudal peduncle length is from base of last anal-fin ray at its exit from the musculature, not the skin, to posterior edge of lower hypural plate. The fishing "lure," or illicium, in ceratioids is tipped with a luminescent organ, the esca, except in the Caulophrynidae, and in the gigantactinid genus Rhynchactis. The illicium of Rhynchactis macrothrix, a species that has been taken just outside our area, is tipped with a slight bulbous swelling, but this is not an esca (Bertelsen and Pietsch, 1998). Often the esca is adorned with spinules, papillae and a variety of filaments and appendages. Illicium length is measured from its base only to the distal end of the bulb of the esca. Other light organs include the hyoid barbels of Linophryne and the dorsal midbody caruncles in ceratiids. Tooth counts are a summation of both sides of each jaw. Free-living metamorphosed males have a unique toothy bone articulating with the anterior ends of both jaws called the upper and lower denticulars; their denticles form into rows and are counted. Scales are often modified into skin spinules (= prickles or dermal spines elsewhere), or greatly enlarged bucklers (Himantolophidae). The bulk of our specimens were collected during routine cruises of the South African Sea Fisheries Research Institute’s vessel AFRICANA since 1984, and CSIR’s vessel MEIRING NAUDÉ during a programme with the South African Museum between 1975 and 1979 (Louw, 1977, 1980). Ship stations in the materials sections of the former are designated "A" and of the latter, "SM." Other specimens from voyages of foreign vessels, particularly from Denmark, Germany, Russia and Spain, are detailed in full. Order of presentation of families follows Bertelsen (1984), repeated by Eschmeyer (1990) and Nelson (1994). Vernacular names of families are after Weitzman (1997). Diagnoses are of the various taxa (families, genera, species) worldwide. The species descriptions are based on southern African specimens only when we have examined the material; otherwise the descriptions are taken from the literature. Damaged specimens missing crucial anatomical parts (like barbels or escae) usually cannot be identified; these are listed in each family account. Synonymies are not extensive here and are mainly confined to the most important and/or illustrative literature. The major revisionary literature is identified in each section and should be consulted for complete synonymies and other illustrative or descriptive accounts. Our southern Africa coverage area is expanded from that of Smith and Heemstra (1986) because we feel that their coverage is restrictive with regard to deep-pelagic fishes, as it is biased against usually broad, open-ocean distributions. This step is taken because the southern Africa region is a major transit zone between the two areas of highest ceratioid diversity, the North Atlantic and the Indo-West Pacific. Our area is still restricted somewhat and extends from southern Angola (15ES) west to the Greenwich meridian, south to 40ES, east to 40EE and north to 15ES off Mozambique. Several species are known to occur just outside this limit and are not included here. Abbreviations for museums holding specimens are: BOC: Bingham Oceanographic Collection, Yale University, USA; BMNH: British Museum (Natural History), London, U.K.; IIPB, Instituto de Investigaciones Pesqueras de Barcelona, Spain; IOAN: P.P. Shirshov Institute of Oceanology, Moscow, Russia; ISH, Institut für Seefischerei, Hamburg, Germany; RUSI, J.L.B. Smith Institute of Ichthyology, Grahamstown; SAM, South African Museum, Cape Town; UW, University of Washington, Seattle, USA; ZIN (ZISP or ZIL elsewhere), Zoological Institute, St. Petersburg, Russia; ZMUC, Zoologisk Museum, University of Copenhagen, Denmark. Other abbreviations are: A, anal-fin rays; C, caudal-fin rays; D, dorsal-fin rays; P, pectoral-fin rays; BT, bottom trawl; MWT, midwater trawl (usually an Engels-308 or an RMT-8, both fished open); SL, standard length. KEY TO FAMILIES (Females) This key, slightly modified from that of Bertelsen and Pietsch (1983), is based only on those genera of the families presently known from our waters. 1a. No bulbous light organ (esca) on tip of illicium (cephalic lure); fleshy hair-like cirri on head, jaws and body; longest rays of dorsal and anal fins more than 60% SL; D 14-22; A 12-19 Caulophrynidae 1b. Esca present on illicium tip; no hair-like cirri; longest rays of dorsal and anal fins much less than 60% SL; D 3-17;A 3-7 2 2a. Soft dorsal-fin rays 12-17 Melanocetidae 2b. Soft dorsal-fin rays 3-8 3 3a. Two or three bulbous swellings (caruncles) on midline in front of dorsal fin Ceratiidae 3b. No caruncles on dorsal midline 4 4a. A second cephalic ray present immediately posterior to base of illicium, with a distal luminous gland (this withdrawn in large specimens, its presence indicated by a pore in illicial base); minute sensory papillae in single rows around head, dorsum and caudal peduncle Diceratiidae 4b. Second cephalic ray not present; no sensory papillae in rows 5 5a. Upper jaw extending well in front of lower jaw; esca with a distal appendage bearing 3 hooks Thaumatichthyidae 5b. Upper and lower jaws equal or subequal 6 6a. Illicium emerging on snout tip; caudal peducle length more than 20% SL Gigantactinidae 6b. Illicium emerging behind snout tip, in front of or above eyes; caudal peduncle length 2-17% SL 7 7a. Skin with dermal spines or plates 8 7b. Skin naked 9 8a. Skin covered with large bony plates (bucklers), each with a large median spine Himantolophidae 8b. Skin covered with fine spines; large oval pit anterior to eye in specimens greater then 42 mm SL Centrophrynidae 9a. Hyoid barbel present (Linophryne only); branchiostegal rays 4-5; D and A with 3 rays (rarely 2 or 4) Linophrynidae 9b. Barbel absent; branchiostegal rays 6; D 4-7; A 4-6 Oneirodidae KEY TO FAMILIES (Free-living males) 1a. Nostrils opening laterally on snout; D 5-6; A 4-5; P 14-18 Himantolophidae 1b. Nostrils opening forward at snout tip 2 2a. Hyoid barbel present; symphysial spine on dentary; D 6-7; A 5-6; P 15-17 Centrophrynidae 2b. No hyoid barbel or symphysial spine; D 6; A 4 P 17 Oneirodidae FAMILY CAULOPHRYNIDAE DIAGNOSIS: Females with globose bodies; esca a simple rod, with 5 minute appendages, the illicium more than 200% SL (Robia), or filamentous, illicium less than SL (Caulophryne); dorsal and anal-fin rays extremely elongate; D 6 (Robia) or 14-22 (Caulophryne); A 5 (Robia) or 12-19 (Caulophryne); P 15-18; C 8; body and head with hair-like cirri (seismosensory neuromasts at tips of these); branchiostegal rays 6 (rarely 5). Other characters, mostly osteological, given by Pietsch (1979). Males known from three specimens, two attached to females and one in metamorphosis (Bertelsen, 1951, fig. 11C; Pietsch, 1979, fig. 23). Maximum length 14 cm SL. Males parasitic. Meso- and bathypelagic from about 275 to below 1500 meters. Two genera, Robia Pietsch, 1979, with one species, and Caulophryne Goode & Bean, 1896, with 4 species, one of which is known in our area from a single female. Last reviewed by Pietsch (1979) who recognized 3 species. Balushkin and Fedorov (1985) described a fourth, Caulophryne pietschi, from the southwestern Pacific (Campbell Plateau). Caulophryne Goode & Bean, 1896 Caulophryne polynema Caulophryne jordani polynema: Bertelsen, 1951: 33, 37. fig. 12, (as subspecies of C. jordani). Grey, 1956: 233 (syn., distr.). Lloris, 1986: 249, fig. 130 (descr.; off Namibia). DIAGNOSIS: Illicium with translucent filaments along entire length, more than 50 in specimens larger than 32 mm SL; illicial length less than 35% SL; upper jaw with 30 or more teeth in specimens larger than 20 mm SL; D 19-22; A 17-19. DESCRIPTION: (Data from Lloris, 1986; specimen damaged). Illicial filaments translucent, most filaments arising from posterior surface. Esca a posterolateral appendage with a flattened, bilobed, opaque distal tip and a terminal appendage also with an opaque tip. Escal appendages highly filamentous, these conveying light to their tips in life (Pietsch, 1979, fig. 21). Jaws subequal, with two rows of hair-like cirri on lower jaw, coursing dorsally along preopercular margin. Cirri also present on snout, suborbital region, top of head and along dorsum to caudal peduncle. Dorsal-fin base 24% SL; anal-fin base 15% SL; illicial length 29% SL; pectoral-fin 18% SL; snout 16% SL; eye diameter 3% SL. Counts: D 22 (24 shown in Lloris, 1986, fig 130); A 18; P 16 (14 shown); C 8. DISTRIBUTION: In northern and southeastern Atlantic; off Hawaii and southern California, USA. Two specimens from opening-closing midwater trawls were captured between 900 and 1250 m; all others are from open nets (Pietsch, 1979). MATERIAL: IIPB 383/1981 (female; 100 mm SL); off Orange Banks, Namibia; 28E02'S, 14E35'E; BT, 0-585 m; MUESTREOS (of BENGUELA programme); 7 Nov. 1980 (Lloris, 1986). Not seen. FAMILY MELANOCETIDAE DIAGNOSIS: Females with gelatinous, globose body; mouth steeply oblique; jaws with elongate, depressible fangs anteriorly; illicium and esca without denticles; escae simple, with conical tip or single, short appendage, anterior and posterior crests in larger M. johnsonii; hyoid barbel absent; minute epidermal spines (skin spinules) present in some species; D 12-17; A 3-5; P 15-23; C 9; branchiostegal rays 6; vertebrae 20. Other (osteological) characters are given by Pietsch and Van Duzer (1980: 67). Males without jaw teeth; eyes ovoid, directed laterally, with aphakic space (pupil larger than lens); lower denticular bone with medial, forward-projecting group of teeth and a right and left, upwardly-directed lateral group; upper denticular bone with a few semicircular series of teeth fused with a median series of 3-9 enlarged dermal spines (Bertelsen, 1951, fig. 14); nostrils lateral, nasal lamellae 9-24, increasing with size; D 12-16; A 4; C 9. A single genus, Melanocetus Günther, 1864, with 6 species, 2 in our area. Last reviewed by Pietsch and Van Duzer (1980). Males free-living, not yet known from our area. Adults meso- and bathypelagic, from about 250 m to below 3000 m. Females of M. johnsonii reach 18 cm SL. Recorded prey includes fishes and crustaceans. Munk et al. (1998) described the development of the escal light organ in two species showing that the luminescent bacteria are not present in the organ until a duct connecting the light gland to the exterior develops, strongly suggesting that the bacteria enter the light organ from the sea. Melanocetus rotundatus Gilchrist, described from off South Africa, was considered "almost certainly" a junior synonym of M. johnsonii (Penrith, 1967; Pietsch and van Duzer, 1980). The two syntypes of M. johnsonii have been lost (Winterbottom, 1974; Gon and Skelton, 1997). REMARKS: Pietsch and Van Duzer (1980) examined numerous male specimens, but concluded that since the work of Bertelsen (1951), they could add no new diagnostic characters to adequately identify any male with a species based on females. However, Bertelsen (1986) identified and illustrated a 28 mm male, apparently covered with skin spinules, as M. johnsonii, and provided a key to males of both M. johnsonii (with concentration of subdermal pigment on caudal peduncle) and M. murrayi (no pigment subdermally on caudal peduncle). Detecting the shape of the vomer in females of the two species in our area requires a probe or some dissection. Small specimens of M. murrayi have a much less bowed vomer than that shown in Pietsch and Van Duzer (1980, figs 2, 5). KEY TO SPECIES (Females) 1a. Anterior margin of vomer almost straight; number of lower jaw teeth about 35-75 in specimens over 25 mm SL; pectoral base width 11-18% SL; skin spinules covering body and head (only to top of head in small specimens) M. johnsonii 1b. Anterior margin of vomer concave; number of lower jaw teeth about 65-140 in specimens over 25 mm SL; pectoral base width 6-9% SL; skin spinules few, restricted to caudal peduncle M. murrayi Melanocetus johnsonii Melanocetus johnsonii Günther, 1864: 301, pl. 25 (off Madeira). Goode & Bean, 1896: 494, fig. 406 (after Günther, 1864). Fitch & Lavenberg, 1968: 127, fig. 70. Paulin et al., 1989: 140. Melanocetus johnsoni: Brauer, 1906: 318. Regan, 1926: 4-6, 8-10, 18, 33, fig. 10. Parr, 1927: 29. Norman, 1930: 354. Regan & Trewavas, 1932: 27-29, 49-52, figs. 19-22, 72, 73. Koefoed, 1944: 3,pl. 1, fig. 1. Bertelsen, 1951: 7, 40, 41, 43-46, 48-53, figs. 13, 15, 17-19. Grey, 1956: 235 (syn., males). Maul, 1961: 90, fig.1. Maul, 1962: 36, fig. 2 (male). Struhsaker, 1962:841. Pietsch & Van Duzer,1980: 71, figs. 1, 3, 6, 16B, 17, 25 (fam. rev.). Pietsch & Seigel, 1980: 381. Bertelsen & Pietsch, 1983: 81, fig. 2. Masuda et al., 1984: 108, pl. 93I. Pietsch, 1986a: 375, text fig. Lloris, 1986: 242, fig. 124. Paxton et al., 1989: 287. Bertelsen, 1991: 492. DIAGNOSIS: Esca bilobed, with posterior and usually anterior crests; number of lower jaw teeth 32-78. anterior margin of vomer almost straight; minute skin spinules cover most of body; interorbital width 14-29% SL; width of pectoral base 11-18% SL; escal bulb width 4-9% SL; illicium length 32-61% SL. See also Pietsch and Van Duzer (1980). DESCRIPTION: Esca with distal prolongation; anterior and posterior crests in largest specimens. Head deeper than long, mouth vertical, eye minute. Measurements in percent SL: head length 37-48; head width 35-51; head depth 65-77; postorbital 15-32; upper jaw 52-72; lower jaw 53-75; snout 9-21; eye diameter 5-8; interorbital width 18-26; longest upper jaw tooth 7-15; longest lower jaw tooth 12-18; illicium 36-48; esca length 8-12; esca esca 4-8; gill slit 17-24; pectoral base width 10-16; pectoral-fin 16-31; predorsal 59-67; preanal 82-84; body depth at anal-fin origin 39-49; dorsal-fin base 29-32; anal-fin base 7-10; longest dorsal ray 20-40; longest anal ray 16-35; least caudal peduncle depth 16-20; peduncle length 6-16. Counts: D 13-16; A 3-4; P 17-19; upper jaw teeth 32-87; lower jaw teeth 34-62; vomerine teeth 5-8. DISTRIBUTION: In all three major oceans from about 50EN to 50ES (Pietsch and Van Duzer, 1980, fig. 30). REMARKS: Three specimens from our area with greatly distended stomachs were examined for prey items. These were: 1) SAM 34768 (51 mm SL), with a juvenile macrourid, Nezumia micronychodon, and two lanternfish, Lampanyctus sp. Each of these prey exceeded 10 cm; 2) SAM 31207 (57 mm SL), with a 62 mm lampanyctine myctophid; and 3) SAM 31582 (53 mm SL), with 5 Maurolicus walvisensis, 23-32 mm SL, 1 Bolinichthys longipes, 87 mm SL, and 1 Electrona carlsbergi, 40 mm SL. MATERIAL (all females): SAM 31207 (57 mm SL); off Cunene R. mouth; 17E57'S, 11E18'E; BT, 0-1000 m; AFRICANA (no sta.); 28 June 1986. SAM 29456 (26 mm SL); off Rocky Point, Namibia (ca. 19ES); BT; SFRI crab survey, sta. 51; 26 Aug. 1982. SAM 34768 (51 mm SL); northern Namibia; 19E58'S, 11E43'E; BT, 0-500 m; FRIDTJOF NANSEN sta. 473; 1 Oct. 1990. SAM 34769 (38.5 & 65 mm SL); NW of Lüderitz, Namibia; 25E38'S, 13E32'E; BT, 0-644 m; FRIDTJOF NANSEN sta. 1386; 2109 hrs; 30 Oct. 1992. SAM 31581 (13.5 mm SL); Cape Canyon; 33E31.8'S, 17E25.7'E; MWT, 0-756 m; sta. A6994-060; 6 Mar. 1988. SAM 31582 (34 & 53 mm SL); Cape Canyon; 33E38.1'S, 17E24.1'E; MWT, 0-1000 m; sta. A6996-060; 7 Mar. 1988. RUSI 47245 (50 mm SL); Cape Canyon; 34E16'S, 17E29'E; BT, 0-1150 m; sta. A17092-127; 11 Jan. 1995. SAM 31589 (25 mm SL); Cape Point Valley; 34E53.0'S, 18E09.1'E; MWT, 0-920 m; sta. A7014-060; 11 Mar. 1988. SAM 31591 (13 mm SL); Cape Point Valley; 34E56.7'S, 18E13.1'E; MWT, 0-912 m; sta. A7032-060; 13 Mar. 1988. RUSI 61330 (15 mm SL); Agulhas Bank; 35E57'S, 19E24'E; MWT, 0-720 m; FRIDTJOF NANSEN sta. AN 0036; 21 Jan. 2000. SAM 34506 (16.5 & 19 mm SL); off St. Francis Bay; 34E55.8'S, 24E41.4'E; MWT, 0-952 m; sta. A14101-107; 18 Oct. 1992. SAM 28406 (11.5 mm SL); off East London; 33E55.9'S, 27E25.3'E; MWT, 0-982 m; sta. SM 187; 31 May-1 June 1978. SAM 29235 (17 mm SL); off Kei R. mouth; 32E30.8'S, 29E13.4'E; MWT, 0-916 m; sta. SM 218; 23 Jun. 1979. SAM 27809 (10 mm SL); off Durban; 30E17.1'S, 31E25.2'E; MWT, 0-750 m; sta. SM 148; 17 May 1977. SAM 27806 (12 & 13 mm SL); off Durban; 30E21.3'S, 31E15.7'E; MWT, 0-830 m; sta. SM 138; 14 May 1977. SAM 27805 (3; 12-14 mm SL); off Durban; 31E30.4'S, 30E04.0'E; MWT, 0-1129 m; sta. SM 145; 16 May 1977. SAM 29234 (12 mm SL); off Durban; 31E33.1'S, 30E18.2'E; MWT, 0-1050 m; sta. SM 195; 19 Jun. 1979. Melanocetus murrayi Melanocetus murrayi Günther, 1887: 57, pl. 11A (central Atlantic). Regan, 1926: 32, 33 (in part). Parr, 1927: 27. Regan & Trewavas, 1932: 27, 49-50, fig. 22C, 23, 71. Bertelsen, 1951: 40, fig. 16. Grey, 1956: 234 (syn., distr.). Maul, 1962: 37, fig. 3 (male). Pietsch & Van Duzer, 1980: 81, figs. 2, 4, 5, 7 –15, 16a (fam. rev.). Pietsch & Seigel, 1980: 381. Bertelsen & Pietsch, 1983: 81, fig. 3. Bertelsen, 1984: fig. 169D (larval male). Bertelsen, 1986: 1376, text figs. Paxton et al., 1989: 287; Bertelsen, 1991: 493. DIAGNOSIS: Females with anterior margin of vomer concave, more deeply so in larger specimens; skin spinules sparse, only on caudal peduncle; interorbital width 9-18 % SL; width of pectoral base 6-9 % SL; escal bulb width 2-5 % SL; illicium length 23-37 % SL; esca conical to barely lobate in largest specimens; number of lower jaw teeth 46-142. See also Pietsch and Van Duzer (1980). DESCRIPTION: Esca with conical distal end and no crests. Head deeper than long, mouth vertical, eye minute. Measurements in percent SL: head length 35; head width 40; head depth 68; postorbital 21; upper jaw 59; lower jaw 64; snout 10; eye diameter 5; interorbital width 26; longest upper jaw tooth 8; longest lower jaw tooth 10; illicium 35; esca length 8; esca width 4; gill slit 15; pectoral-base width 14; pectoral-fin 21; predorsal 64; preanal 78; body depth at anal-fin origin 43; dorsal-fin base 26; anal-fin base 6; longest dorsal ray 31; longest anal ray 31; least caudal peduncle depth 18; peduncle length 17. Counts: D 14; A 4; P 18; upper jaw teeth 59; lower jaw teeth 57; vomerine teeth 4. DISTRIBUTION: All three major oceans and primarily tropical-subtropical. One record above 60EN off SE Greenland (Pietsch and Van Duzer, 1980, fig. 30). MATERIAL: SAM 29233 (female, 30 mm SL); off Port St. Johns, South Africa; 31E 34.0'S, 30E20.6'E; MWT, 0-2166 m; sta. SM 194; 19 Jun. 1979. FAMILY HIMANTOLOPHIDAE DIAGNOSIS: Females with globose bodies, flesh gelatinous in small specimens, thickening in largest; large specimens covered with round, bony plates with a central spine ("bucklers"), these sparsely distributed in small individuals; mouth oblique, cleft reaching to, or in front of, eye; teeth depressible, retrorse (angled backward), not fang-like, numerous, in close-set rows anteriorly; vomerine teeth absent; snout and chin densely covered with sensory papillae; ilicium and esca with or without denticles; basal part of lure (pterygiophore of illicium) not protruding on snout; escae bulbous, with anterior, dorsal and/or posterior appendages, their morphology specifically variable; hyoid barbel absent; D 5-6; A 4; P 14-18; C 9; branchiostegal rays 6; vertebrae 19. Males: metamorphosed specimens without jaw teeth, denticular "teeth" become fused into solid denticular bone mass with increasing size, with 16-31 hooked denticles on upper bone; enlarged dermal spines above upper denticular bone in snout midline; nostrils lateral; posterior nostril greatly enlarged, with 10-17 lamellae; eyes spherical in metamorphic specimens, becoming more ovoid, with a narrow anterior aphakic space (pupil larger than lens) in adults; skin spinules covering head and body; sphenotic spines absent. Counts as for females. A single genus, Himantolophus Reinhardt, 1837, with 18 species, 5 in our area. Last reviewed by Bertelsen and Krefft (1988) on the basis of all the then known specimens. Males free living, two specimens from our area. Adults primarily mesopelagic, few from nets fished below 1000 m. Larvae epipelagic. Females feed mainly on cephalopods and other fishes, but also eat shrimp and krill (euphausiids). Found in all oceans between subtropical convergences (about 40EN to 40ES), largest females sometimes in higher latitudes. Females of H. groenlandicus reach 60 cm. INDETERMINATE (all females): SAM 27083 (202 mm SL); off Durban; from whale stomach; July 1974; P. Best (digested; no esca). RUSI 61315 (35 mm SL); Agulhas Bank; 35E59'S, 19E26'E; BT, 0-938 m; FRIDTJOF NANSEN sta. AN 0035; 21 Jan. 2000 (illicium torn away). RUSI 61329 (19 mm SL); Agulhas Bank; 35E57'S, 19E24'E; MWT, 0-720 m; FRIDTJOF NANSEN sta. An 0036; 21 Jan. 2000 (esca undeveloped). KEY TO SPECIES This key to females in our area, based on escal morphology, is adapted from Bertelsen and Krefft (1988). See figures 2-7 for terminology of escal appendages. 1a. Posterior escal appendages (PA) present 2 1b. Posterior (PA), anterior (AA), and posterolateral escal appendages (PLA) absent; illicial appendages (IA) absent H. cornifer 2a. Distal escal appendage (DA) divided at its base; spines present on esca or escal appendages in specimens > 35 mm SL; length of DA 0.9-20% SL; posterolateral appendages (PLA) usually present 3 2b. DA undivided at base (but divided slightly distal of base); no spines on esca or escal appendages; length of DA 6-74 % SL; PLA absent 4 3a. Anterior escal appendage (AA) present; illicial appendages (IA) present; posterior appendage (PA) undivided at base H. groenlandicus 3b. AA absent; IA absent; PA divided at base H. appelii 4a. Each main branch of dorsal escal appendage (DA) bifurcated at tip .H. pseudalbinares 4b. DA branches simple, without bifurcations H. albinares Himantolophus albinares Himantolophus albinares Maul, 1961: 111, figs. 11-15 (off Madeira). Maul, 1962: 8, figs. 1, 2. Trunov, 1981: 62, fig. 5. Bertelsen, 1986: 1378, text fig. Bertelsen & Krefft, 1988: 54, figs. 19, 20. DIAGNOSIS: Females: posterior escal appendage (PA) bifurcating near base, no side branches from main branches, tips furcate; distal escal appendage (DA) undivided at base, undivided part 1-9% SL; and with simple (unbranched) tips; no spines or posterolateral appendages (PLA) on esca; illicium length 25-ca. 60% SL; illicial stem with 2-9 simple or branched appendages. Males unknown. DESCRIPTION: Posterior escal appendage (PA) whitish or with scattered melanophores, bifurcated near base, with 2-4 filamentous branches at tips. Distal escal appendage (DA) also deeply bifurcated near its base. Dorsalmost illicial appendages (IA) at proximal end of esca. Skin naked in specimens less than about 35 mm SL. Anterior facial papillae well developed except in juveniles less than about 30-40 mm SL. Head deeper than long, more evenly rounded in young. Jaws subequal, mouth terminal, oblique. Eye small, decreasing in size relative to SL with growth. Smallest specimens with a dorsal hump just anterior to dorsal fin; hump whitish with central, darkly pigmented area; remnant of hump traceable in some larger specimens. Bertelsen and Krefft (1988: tab. 10) give some morphometric characters of the illicium. Colour generally brownish black with occasional white patches on caudal peduncle, along bases of unpaired fins, snout and chin. Counts: D 5; A 4; P 15-17. DISTRIBUTION: North and South Atlantic. Apparently mesopelagic, this species has been taken as shallow as 170-330 m by midwater trawl as well as in the relatively shallow Madeira longline fishery (Bertelsen and Krefft, 1988). MATERIAL: IOAN uncat. (female, 141 mm SL); Valdivia Bank off Namibia; ca. 24ES, 06EE; no other data (Trunov, 1981). Not seen. Himantolophus appelii Aegeonichthys appelii Clarke, 1878: 145, pl. 6 (off New Zealand). Waite, 1912: 194, pl. 10 (second spec., from line fisherman, New Zealand). Himantolophus appelii: Regan, 1926: 41. Regan & Trewavas, 1932: 60, fig. 89. Bertelsen, 1951: 59, 66 (as H. appeli). Grey, 1956: 241 (as H. appeli). Bertelsen & Pietsch, 1983: 82, fig. 4 (addl. mat’l.). Bertelsen & Krefft, 1988: 47-50, 75, 76, 80, 81, figs. 16, 35C; Paxton et al., 1989: 289. Paulin et al., 1989: 139-140. Himantolophus groenlandicus (not of Reinhardt, 1837): Smale et al., 1995: 70, pl. 36. DIAGNOSIS: Females: posterior escal appendage (PA) divided at base, with 1-7 side branches on each main branch (Fig. 3); distal appendage (DA) divided at base; anterior appendage (AA) absent; posterolateral appendages 0-5; spines present on illicium, esca and escal appendages in specimens greater than 30 mm SL; no distinct white patches on body; caudal fin rays white or faintly dusky in specimens less than about 100 mm SL. Males with 11-13 olfactory lamellae; diameter of posterior (enlarged) nostril 6-7% SL; snout length 15-18% SL; 20-25 upper and 22-32 lower denticular "teeth;" eyes of largest males with slight, anterior aphakic space (pupil larger than lens). DESCRIPTION: Females: Smallest (24 mm SL) with posterior escal appendage (PA) just developed, with 1-2 side branches; distal appendage (DA) a mere nubbin; no posterolateral appendage (PLA). Larger females with 1-6 side branches on posterior escal appendage (Fig. 3A); posterolateral appendage absent in largest fish (150 mm SL) (Fig. 3B). Skin naked in smallest specimens; bucklers develop after a length of about 25 mm SL. Head massive, deeper than long; anterior facial sensory papillae just developed in 45 mm SL specimen. Jaws subequal. Eye small, relatively larger (7-8% SL) in smallest two specimens. Measurements in percent SL: head length 38-45; head width 18-40; head depth 59-76; postorbital 18-27; upper jaw 32-35; lower jaw 35-41; snout 12-18; eye diameter 5-8; interorbital width 14-20; longest upper jaw tooth 4; longest lower jaw tooth 5-6; illicium 36-39; esca length 7.5-11; esca width 5-6; gill slit 11-15; pectoral base width 11-13; pectoral-fin length 17-21; predorsal 79-86; preanal 84-90; body depth at anal-fin origin 21-35; greatest body depth 58-78; dorsal-fin base 8-10; anal-fin base 5-9; longest dorsal ray 17-32; longest anal ray 15-27; least caudal peduncle depth 14-18; peduncle length 5-9. Counts: D 5; A 4; P 16-17; upper jaw teeth 73-160; lower jaw teeth 80-166. Males: Body fusiform. Head ovoid, longer than deep. Jaws subequal, toothless. Eye rounded, with small aphakic space. Measurements in percent SL: head length 39-40; head width 20; head depth 33-34; postorbital 25-26; upper jaw 14; lower jaw 18-19; greatest diameter of posterior nostril 6.5-6.8; snout 15; eye diameter 8; pectoral base width 8; pectoral-fin length 20-22; predorsal 74; preanal 77-78; body depth at anal-fin orgin 30; greatest body depth 42-43; dorsal-fin base 9; anal-fin base 6-7; longest dorsal ray 25-28; longest anal ray 18; caudal peduncle depth 16-17; peduncle length 8. Counts: D 5; A 4; P 16; nasal lamellae 12-13; upper denticular teeth 21-23; lower denticular teeth 22-25 (see below). DISTRIBUTION: Generally along the southern subtropical convergence (except Africa) from South America east to New Zealand. Apparently mesopelagic, this species has been taken as shallow as 195-205 m in midwater trawls; also from sperm whale stomachs (Bertelsen and Krefft, 1988). REMARKS: Bertelsen and Krefft (1988: 75) tentatively referred the two males, SAM 30981 (incorrectly as SAM 27805), to this species. This was done on the basis of minor differences in their number of olfactory lamellae, denticular teeth, snout length, and posterior nostril and eye diameters with two Australian males of virtually the same size, but greater sexual maturity. However, in their closing remarks on SAM 30981 (and key and distribution map, fig. 45), they as much as admit that the juvenile condition of these specimens probably accounts for at least the shorter snouts and smaller nostril than the more mature Australian specimens. Bertelsen and Krefft’s tentative identification is further confusing, as they included only the one species in the H. appelii species group! In our view this removes any doubt that SAM 30981 are anything but H. appelii. Our recounts of the denticular teeth in SAM 30981 (above) do not correspond to those given by Bertelsen and Krefft, as these areas are now damaged. Smale et al. (1995: 70, pl. 36) described and illustrated two otoliths of this species, apparently for the first time (but as H. groenlandicus). They were removed from SAM 31615. MATERIAL: Females: SAM 31615 (150 mm SL); Cape Canyon; 33E38.1'S; 17E24.1'E; MWT, 910-990 m (bottom depth 2000 m); sta. A6996-060; 7 Mar. 1988. RUSI 54509 (130 mm SL); Cape Canyon; 33E36'S, 17E07'E; BT, 0-977 m; sta. A18562-139; 24 Jan. 1997. RUSI 54996 (45 mm SL); Agulhas Bank; 36E42'S, 20E15'E; BT, 0-810 m; sta. A18938-144; 17 Apr. 1997. UW 22179 (232 mm SL); Agulhas Bank; 35E10.0'S, 23E11.0'E; BT; 0-580 m; sta. A12108-095; 1 Sept. 1991 (not seen). SAM 34505 (24 mm SL); off Algoa Bay; 34E11.7'S, 26E44.5'E; MWT, 0-750 m; sta. A14030-107; 11 Oct. 1992. Males: SAM 30981 (25, 26 mm SL); off Port St. Johns; 31E30.4'S, 30E04.0'E; MWT, 0-1129 m; sta. SM-145; 16 May 1977. Himantolophus cornifer Himantolophus cornifer Bertelsen and Krefft, 1988: 64, figs. 27, 28 (Coral Sea). DIAGNOSIS: Females with anterior (AA), posterior (PA) and posterolateral (PLA) escal appendages and illicial appendages (IA) absent; distal escal appendage (DA) 9-72% SL, bifurcated at or near base; each main branch of DA with a basal, posterior side branch 3-24% SL; DA with 1-3 distal bifurcations; spines on illicium, esca and DA in specimens over about 55 mm SL; illicial length 21-35% SL; facial papillae prominent, semiglobular; white patches on face, around unpaired fins and caudal peduncle (these becoming obscure with age). Males unknown. DESCRIPTION: Single female. Distal escal appendage (DA) with one of its two main branches severed 1.5 cm above bifurcation at escal base; remaining DA with basal side branch and 4 distal branches (Fig. 5). Three escal pores, with well developed lobe covering two; few escal spines on interior of lobe. Bucklers well developed; on ventral, lateral and dorsal surfaces of body, caudal peduncle, pectoral base and in axil; bucklers absent on head except ventrally. Head massive, globose, deeper than long; anterior facial papillae well developed. Mouth oblique; lower jaw longer than upper, protruding well beyond upper. Eye minute, smaller than longest dentary tooth. Measurements in percent SL: head length 34; head width 19; head depth 61; postorbital 21; upper jaw 20; lower jaw 25; snout 13; eye diameter 2.5; interorbital width 10; longest upper jaw tooth 2; longest lower jaw tooth 3; illicium 21; esca width 11; DA 59 (DA apparently lost some of its length due to digestion inside whale; see below); basal side branch of DA 24; base of DA to side branch 10; gill slit 10; pectoral base width 9; pectoral-fin 13; predorsal 80; preanal 92; body depth at anal-fin origin 31; greatest body depth 65; dorsal-fin base 12; anal-fin base 9; longest dorsal ray 17; longest anal ray 15; least caudal peduncle depth 16; peduncle length 4. Counts: D 5; A 4; P 17; upper jaw teeth 256; lower jaw teeth 234. DISTRIBUTION: Tropical waters of the northwestern and central Atlantic, Western Indian Ocean (Natal), Coral Sea and central Pacific. Meso- and bathypelagic; caught in closing nets at 450 m and at 1850-1900 m (Bertelsen and Krefft, 1988). MATERIAL: SAM 34508 (female, 208 mm SL); Salmon Bay, South Africa; 29E25'S, 32E33'E; stomach of sperm whale (beach stranding); 2 Apr. 1973; P. Best. Himantolophus groenlandicus Himantolophus groenlandicus Reinhardt, 1837: 116, pl. IV (off Greenland). Regan, 1926: 40, fig. 23 (in part). Regan & Trewavas, 1932: 59, fig. 88A, pl. I, fig. 1 (in part). Smith, 1949: 429, fig. 1230. Bertelsen, 1951: 60, figs. 24C, 24D (in part). Grey, 1956: 239-240. Smith, 1965: 429, fig. 1230. Leim & Scott,1966: 426. Lloris, 1986: 246-247, fig. 128. Pietsch, 1986a: 376, text fig. Nielsen & Bertelsen, 1992: 62, text fig. Himantolophus reinhardti Lütken, 1878: 320, pls. I, II (off Greenland). Himantolophus ranoides Barbour, 1942: 83, pls. XIII, XIV (off E. Canada). DIAGNOSIS: Females: anterior escal appendage (AA) present; distal escal appendage (DA) bifurcated near base, 0.9-4.2% SL; posterior escal appendages (PA) undivided at base, with 2-6 branches distally in specimens over 60 mm SL; 2-11 total posterolateral escal (PLA) and illicial appendages (IA), arranged more or less in pairs and either simple or with up to 9 branches per appendage; escal lobes blunt; illicium length 28-54% SL; spines develop on illicium, esca and appendages in specimens at about 35-50 mm SL; bucklers develope at about 50 mm SL. Males probably of this species were tentatively placed in the "H. brevirostris" group (Bertelsen and Krefft, 1988: 80). DESCRIPTION: Single female. Esca with simple anterior appendage (AA), distal appendage (DA) bifurcated near midlength; posterior appendage (PA) with 3 branches; posterolateral appendage with 4 branches. Bucklers well developed, on all body surfaces, caudal peduncle, pectoral base and in axil; bucklers absent on dorsal surface of head. Head massive, deeper than long; anterior facial papillae well developed. Mouth oblique, lower jaw projecting beyond upper. Eye minute, rounded. Measurements in percent SL: head length 33; head width 21; head depth 56; postorbital 14; upper jaw 27; lower jaw 31; snout 15; eye diameter 2; interorbital width 14; longest upper jaw tooth 3; longest lower jaw tooth 6; illicium 43; esca length 11; esca width 8.5; gill slit 10; pectoral base width 10; pectoral-fin 14; predorsal 82; preanal 92; body depth at anal fin origin 24; dorsal fin base 16; anal fin base 10; longest dorsal ray 17; longest anal ray 10; least caudal peduncle depth 15; peduncle length 6.5. Counts: D 5; A 4; P 15; upper jaw teeth 177; lower jaw teeth 133. DISTRIBUTION: North and South Atlantic; western Indian Ocean off South Africa and possibly farther north (Bertelsen and Krefft, 1988, fig. 38). Recorded trawl depths range 250-1800 m. MATERIAL: SAM 23120 (female, 216 mm SL); Kommetjie beach, Cape Peninsula, South Africa (stranded); 34E08.65'S, 18E19.40'E; 20 July 1961. Mistakenly reported as SAM 23129 by Bertelsen and Krefft (1988: 37). Himantolophus pseudalbinares Himantolophus pseudalbinares Bertelsen & Krefft, 1988: 59, fig. 23 (off Cape St. Francis, South Africa). DIAGNOSIS: Anterior escal appendage (AA) absent; dorsal escal appendage (DA) undivided at base, with bifurcated, whitish tips, DA 27% SL (holotype); posterior escal appendage (PA) bifurcated at base; illicial appendages (IA) present; posterolateral escal appendages (PLA) absent. DESCRIPTION: Tip of esca whitish, with paired swellings. No spines on escal bulb, appendages or illicium. Posterior escal appendage (PA) proximally pigmented, distal tips of 2 main branches bifurcate. Dorsal escal appendage (DA) dark, shallowly bifurcated at tips; tips white. Illicial appendages (IA) arranged in two pairs, with a fifth small filament. Bucklers present on body and pectoral lobe. Facial papillae well developed, more lightly pigmented than rest of head and body. Body without whitish patches. Measurements in percent SL: length illicium 52; escal width 9; undivided portion of DA 7; PA 13. Counts: D 5; A 4; P 16. DISTRIBUTION: Known from only one specimen caught by a Soviet fisheries vessel southwest of Port Elizabeth, RSA. MATERIAL: ZIN 49711 (female, 82 mm SL; holotype); off Cape St. Francis; 35E01.0'S, 24E36.8'E; BT; 1280-1300 m; FIOLENT cr. 3-63. Not seen. FAMILY DICERATIIDAE DIAGNOSIS: Females with a short, second cephalic ray (lure with esca)immediately behind main lure, often hidden beneath a pore in largest specimens; escal bulb with minute denticles on proximal half; body short, globose; mouth large, oblique, cleft extending posteriorly well beyond minute eye; lower jaw with well developed symphysial spine; skin spinules present, dense; sphenotic spines well developed; vomerine teeth present; branchiostegal rays 6; D 5-7; A 4; P 13-16; C 8-9. Diagnostic osteological characters given by Uwate (1979). The double anglers are a small family, just five species in two genera: Diceratias Günther, 1887 and Bufoceratias Whitley, 1931. One species in our area. The preoccupied generic replacement name Phrynichthys Pietschmann, 1926 has been incorrectly used for our species, and the correct (next available) name Bufoceratias is used here, apparently for the first time (see Remarks). The double anglers are meso- or bathypelagic, but apparently B. wedli may become benthic at large sizes. Bertelsen (1951: 70) considered Laevoceratias liparis Parr, 1927 the male of a double angler, but Bertelsen et al. (1981) showed it to be the male of some Gigantactis species. A single male of an uncertain species has been reported from the tropical western Pacific (Bertelsen, 1983). Last reviewed by Uwate (1979). Balushkin and Fedorov (1986) added Diceratias trilobus to the family. Females of B. wedli reach 178 mm SL. Bufoceratias Whitley, 1931 Bufoceratias wedli Phrynichthys wedli Pietschmann, 1926: 88 (off Madeira). Uwate, 1979: 142, figs. 3, 6, 7B, 9, 10B, 11B, 13 18. Bertelsen, 1986: 1381, text fig. Lloris, 1986: 244, figs. 125-127. Pietsch, 1986a: 376, text fig. Machida & Yamakawa, 1990: 60, figs. 1-3. Bertelsen, 1991: 496. Bertelsen 1995: 141, text fig. Paroneirodes wedli: Regan & Trewavas, 1932: 58. Bertelsen, 1951: 70. Grey, 1956: 243. Trunov, 1974: 145. Phrynichthys sp.: Yamakawa, 1984: 288, 289, 385. DIAGNOSIS: Illicial length more than 80% SL; esca with anterior, posterior and posterolateral appendages, these often multifurcate; a luminescent terminal papilla between anterior and posterior appendages; head, dorsum and caudal peduncle with rows of dark seismosensory papillae; vomerine teeth 7-15; upper jaw teeth 21-65; lower jaw teeth 16-44. DESCRIPTION: Head and body globose, flesh gelatinous. Skin spinules densely arrayed, covering whole body including illicium, absent laterally on jaws and esca. Escae with simple posterolateral and posterior appendages (former termed "lateral appendages" by Uwate, 1979). Jaws long and strong, upper extending posteriorly well beyond minute eye, lower jaw projecting slightly beyond upper. Pectoral lobe short; gill opening near its middle. Base of illicium on rear of head or near midbody, illicial pterygiophore beneath flesh. Measurements in percent SL: head length 39-53; head width 29-34; head depth 69-83; postorbital 21-30; upper jaw 37-43; lower jaw 42-54; snout 11-15; eye diameter 5-7; interorbital width 13-15; longest upper jaw tooth 4-6; longest lower jaw tooth 5-8; illicium 153-172; illicial base to anteriormost tip of upper jaw 46-58; esca length 7-14; esca width 4-7; gill slit 10-19; pectoral base width 7-12; pectoral-fin 16-23; predorsal 80-88; preanal 85-95; body depth at anal fin origin 29-42; greatest body depth 73-88; dorsal-fin base 11-12; anal-fin base 7-9; longest dorsal ray 22-27; longest anal ray 17-26; least caudal peduncle depth 17-19; peduncle length 4-7. Counts: D 5-6; A 4; P 13-15; C 8-9; upper jaw teeth 42-54; lower jaw teeth 28-37; vomerine teeth 8-11. DISTRIBUTION: Both sides of the tropical Atlantic, south in our area to Namibia; East China Sea (Uwate, 1979; Yamakawa, 1984; Machida and Yamakawa, 1990). Meso- and bathypelagic, taken in trawls operated from 390 to 1750 m (Uwate, 1979). Bertelsen's (1991) statement "old specimens probably benthic" is corroborated by SAM 29918, taken in a crab trap (below). REMARKS: Pietschmann (1926) erected the genus Phrynichthys for this species, a name preoccupied (in fishes) by Agassiz (1846). A replacement name, Bufoceratias Whitley (1931) was disregarded by Uwate (1979), Bertelsen (1986, 1991) and Pietsch (1986a). Paroneirodes Alcock, 1890, erected as a monotypic genus for an Indo-Pacific diceratiid, P. glomerulosus, was used by Regan and Trewavas (1932) and Bertelsen (1951) to contain glomerulosus and wedli as congeners. However, glomerulosus was synonymized by Uwate (1979) with Diceratias bispinosus Günther, 1887, thus the earlier Paroneirodes cannot be used as a generic replacement name for our species. MATERIAL (all females; off northern Namibia): ZMUC P922153 (117 mm SL); 17E25'S, 11E11'E; BT, 0-787 m; SFRI crab survey, sta. 31; 13 Aug. 1982. ZMUC P921130 (97 mm SL); 18E33'S, 11E25'E; BT, 0-610 m; ERNST HAECKEL, sta. 236; 28 Nov. 1972. SAM 29918 (117 mm SL); 19E59.0'S, 11E28.9'E; beehive crab trap, 800 m; SFRI; 13 Oct. 1982. SAM 29459 (110 mm SL); 20E58.6'S, 12E25.7'E; BT, 0-498 m; BENGUELA sta. 28; 20 Aug. 1982. SAM 28637 (69 mm SL); BT; SFRI; 20 July 1979 (no other data). Also, 5 specimens 30-134 mm SL from Namibia in bottom trawls fished between 183 m and 615 m (Lloris, 1986) (not seen). FAMILY ONEIRODIDAE DIAGNOSIS: Females with single illicium (only remnant of second cephalic ray present), with bulbous esca that may be simple or filamentous; esca without denticles; body globose to moderately elongate, naked except for minute skin spinules in Oneirodes or very evident skin spines in Spiniphryne; sphenotic spines present except in Chaenophryne; jaws equal anteriorly; no hyoid barbel; branchiostegal rays 6; D 4-8; A 4-7; P 13-30; C 9. Osteological characters are found in Pietsch (1974). Males without jaw teeth or epidermal spines on snout; anterior nostrils opening forwards, posterior nostrils lateral, usually larger than eye; anterior end of basal bone above middle of frontal, well removed from upper denticular (Bertelsen, 1951, figs. 6, 30); eyes elliptical, with aphakic space; skin naked. The dreamers are the largest family of deep-sea anglers, with 16 genera and over 60 species, 8 of which have been identified within our region. Pietsch (1974) reviewed the genus Oneirodes and provided a key to the other genera. Males are free living except in Leptacanthichthys which so far has not been taken in our area. Adults are deep meso- or bathypelagic. Larval and male metamorphosis was described by Bertelsen (1951, 1984). Dreamers are found in all oceans including seas of both polar regions. Females reach 30 cm. Recorded prey include fish and a variety of crustaceans. Dolopichthys cornutus Gilchrist & von Bonde, 1924, described from a single female taken at an unspecified locality off South Africa, was designated a nomen dubium by Pietsch (1972a). The type was discarded, the same circumstance as with the types of Melanocetus rotundatus discussed above. INDETERMINATE (all females): Dolopichthys sp.: SAM 28481 (9.8 mm SL, a metamorphosing female with illicium length 1.5 mm and esca torn away); off Durban; 30E17.1'S, 31E25.2'E; MWT, 0-750 m; sta. SM 148; 17 May 1977. Dolopichthys sp.: RUSI 61341 (43 mm SL, illicium torn away); Agulhas Bank; 35E23'S, 18E42'E; BT, 0-950 m; FRIDTJOF NANSEN sta. AN 0042; 22 Jan. 2000. Dolopichthys sp.: SAM 31587 (90 mm SL, badly damaged, no esca); Cape Canyon; 33E40.1'S, 17E22.2'E; MWT; 0-889 m; sta. A7003-060; 8 Mar. 1988. Oneirodes sp.: SAM 34507 (43 mm SL, no esca); eastern Agulhas Bank; 35E10'S, 23E37'E; MWT, 0-1090 m; sta. A14120-107; 20 Oct. 1992. RUSI 61331 (16 mm SL, illicium torn away); Agulhas Bank; 35E57'S, 19E24'E; MWT, 0-720 m; FRIDTJOF NANSEN sta. AN 0036; 21 Jan. 2000. KEY TO GENERA (Females) 1a. Sphenotic spines present 2 1b. Sphenotic spines absent; low ridges across back of head Chaenophryne 2a. Dorsal profile of frontal region of cranium straight (but not parallel to ventral surface of head); skin spines absent Dolopichthys 2b. Dorsal profile of frontal region curved; minute, scattered skin spines present Oneirodes Chaenophryne Regan, 1925 Chaenophryne draco Chaenophryne draco Beebe, 1932: 84, fig. 22 (off Bermuda). Regan & Trewavas, 1932: 85, 89. Bertelsen, 1951: 115. Grey, 1956: 258. Pietsch, 1975: 87, fig. 8. Munk & Bertelsen, 1980: 103-129, figs. 1-20. Pietsch & Seigel,1980: 395. Bertelsen & Pietsch, 1983: 88, fig. 9. Masuda et al., 1984: 106, pl. 92C. Bertelsen, 1986: 1385, text fig. Paxton et al., 1989: 290. Bertelsen, 1991: 498. Chaenophryne parviconus Regan & Trewavas, 1932: 35, 85, 87, figs. 39, 41, 138 (Gulf of Panama). Bertelsen, 1951: 115, fig. 74b, c. Grey, 1956: 258. Chaenophryne atriconus Regan & Trewavas, 1932: 85, 87, fig. 139 (off Cocos Isl., Indian Ocean). Chaenophryne columifera Regan & Trewavas, 1932: 85, 87, 88, fig. 140 (Gulf of Panama). Chaenophryne melanodactylus Regan & Trewavas, 1932: 85, 87, 88, fig 141 (North Atlantic). Chaenophryne macractis Regan & Trewavas, 1932: 85, 88, 89, fig. 142 (SE Pacific). DIAGNOSIS: Esca with an unpaired anterior appendage and no anterolateral or medial appendages; P 16-18, usually 17; ratio of number of upper to lower jaw teeth highest in genus at a given size (1.08-1.61 in specimens 20 mm SL or greater; Pietsch, 1975, fig. 5; this report). DESCRIPTION: Single, slightly damaged female with illicium broken off. Esca with unpaired anterior appendage, posterior portion torn away. Head skinned; massive, as deep as long; frontal greatly convex anteriorly; parietals with low, lateral bulge. Jaws equal, rictus of mouth extending to below mid-orbit, but eyes torn away. Pectoral-fin lobe as high as long. Ratio of upper to lower jaw teeth (totals): 1.61. Measurements in percent SL: head length 48; head width 14; head depth 47; postorbital 48; upper jaw 37; lower jaw 51; interorbital width 15; longest upper jaw tooth 5; longest lower jaw tooth 10; illicium 17; esca length 6; esca width 3; gill slit 20; pectoral base width 7; pectoral-fin 15; predorsal 72; preanal 86; body depth at anal fin origin 30; greatest body depth 56; dorsal-fin base 14; anal-fin base 12; least caudal peduncle depth 11; peduncle length 5. Counts: D 7; A 5; P 16; vomerine teeth 4; upper jaw teeth 50; lower jaw teeth 31. DISTRIBUTION: Tropical and temperate regions of Atlantic, Indian and Pacific oceans. Meso- and bathypelagic. REMARKS: Munk and Bertelsen (1980) studied the microanatomy of the esca of this species, describing the luminous and light-guiding structures in detail. Escae of three specimens of C. draco were illustrated, from which we have redrawn one (Fig. 10). Although our specimen’s esca is damaged, we identify it as C. draco on the basis of its unpaired anterior escal appendage and upper-to-lower tooth ratio of 1.61 (Pietsch, 1975). MATERIAL: SAM 31590 (female, 93 mm SL); Cape Point Valley; 34E56.0'S, 18E12.1'E; MWT; 0-894 m; sta. A7028-060; 13 Mar. 1988. Dolopichthys Garman, 1899 DIAGNOSIS: Body relatively long, not globose; sphenotic spines well developed; rictus (corner) of mouth extending from in front of to beyond eye; pectoral-fin lobe about as long as high, rays confined to distal margin; lower jaw with symphysial spine; illicial pterygiophore (proximalmost structure of lure, or "basal bone" in earlier literature) emerging anterior to sphenotic spines; dorsal margin of frontal bones nearly straight (not bowed); small, rounded papillae of seismosensory system around head, body and caudal fin (Marshall, 1979, fig. 146); posterior margin of opercle bifurcate. Vomerine teeth present or absent. Branchiostegal rays 6; D 5-8; A 4-6; P 17-22; C 9 Other, mostly osteological, characters found in Pietsch (1972a), a revision of the genus. Females reach 190 mm SL. Generic diagnosis for males found in Bertelsen (1951: 99-100), but no male has ever been identified to species (Bertelsen, 1951, 1984, 1986). REMARKS: Three species are known from our area, but a fourth, Dolopichthys allector, has been taken just outside in the south central Atlantic (Pietsch, 1972a, fig. 11). KEY TO SPECIES (Females) 1a. Vomerine teeth 4 or more, usually more than 6; D 5-7, usually 6; A 4-6, usually 5; lower jaw with more than 130 teeth in specimens over 60 mm SL, more than 115 teeth in specimens 25-60 mm SL; posterior escal appendage not bifurcated; no bifurcating papilla on anterior midline of esca. 2 1b. Vomerine teeth 2-4 in specimens less than 35 mm SL, none in larger fish; D 7-8, usually 7; A 6; lower jaw with less than 130 teeth in specimens over 60 mm SL, less than 115 in smaller specimens; posterior escal appendage bifurcated; a small, bifurcated papilla on escal anterior midline D. jubatus 2a. Esca with a posteriorly or posterodorsally directed papilla in dorsal midline; lower jaw with more than 300 teeth in specimens over 70 mm SL, more than 150 in specimens 25-70 mm SL, more than 85 in specimens 18-25 mm SL D. pullatus 2b. Esca with an anteriorly or anterodorsally directed papilla in dorsal midline; lower jaw with less than 300 teeth in specimens over 70 mm SL, usually less than 150 in specimens 25-70 mm, less than 85 in smaller specimens D. longicornis Dolopichthys jubatus Dolopichthys jubatus Regan & Trewavas, 1932: 79, fig. 126 (Atlantic and Indian oceans). Pietsch, 1972a: 17 (lectotype), figs. 1, 3, 7, 11, 12. Bertelsen & Pietsch, 1977: 186. Bertelsen, 1986: 1389, text figs. Bertelsen, 1991: 501. Dolopichthys danae: Regan, 1926: 29 (No. Atlantic; in part, second spec.). Dolopichthys megaceros (not of Holt & Byrne, 1908 [= Oneirodes eschrichtii Lütken, 1871]): Regan, 1926: 29. Dolopichthys longicornis (not of Parr, 1927): Norman, 1930: 354. Bertelsen, 1951: 96, fig. 53. DIAGNOSIS: Vomerine teeth never more than four, lost by a length of about 35 mm SL; posterior escal appendage bifurcated at lengths of 15-25 mm SL; small, unpigmented, bifurcated papilla on anterior edge of esca in midline; illicial length 31-45% SL; D usually 7, A 6. DESCRIPTION (Damaged juvenile female, 15.2 mm SL): Esca with bifurcated posterior appendage and bifurcated anterior papilla (Fig. 11A). Head longer than deep, roughly triangular. Jaws equal anteriorly, rictus extending to vertical in front of eye. Symphysial spine of lower jaw well developed. Sphenotic spines moderately protruding. No skin spines. Posterior margin of opercle strongly bifurcated. Measurements in percent SL: head length 42; head width 30; postorbital 18; upper jaw 34; lower jaw 43; snout 28; eye diameter 7; interorbital width 10; longest upper jaw tooth 3; longest lower jaw tooth 3; illicium 31; esca length 7; esca width 5; pectoral base width 8; pectoral-fin 10; greatest body depth 44; dorsal-fin base 15; anal-fin base 14; longest dorsal ray 29; least caudal peduncle depth 10; peduncle length 12. Counts: D 7; A 6; P 18; vomerine teeth 3; upper jaw teeth 33; lower jaw teeth 47. DISTRIBUTION: All three major oceans in tropical to cold temperate waters. Adults meso- and bathypelagic. MATERIAL: SAM 28480 (female, 15.2 mm SL); off Durban; 30E17.1'S, 31E25.2'E; MWT; 0-750 m; sta. SM 148; 17 May 1977. Dolopichthys longicornis Dolopichthys longicornis Parr: 1927: 18, fig. 6 (Bahamas). Regan & Trewavas, 1932: 66, 79. Bertelsen, 1951: 100 (figs. incorrect). Grey, 1956: 253 (syn., some refs. incorrect). Bussing, 1965: 222. Pietsch, 1972a: 12, fig. 5. Marshall, 1979: 449 (fecundity). Pietsch & Seigel, 1980: 394. Bertelsen, 1986: 1390, text figs. Bertelsen, 1991: 501. Dolopichthys mucronatus Regan & Trewavas, 1932: 79, fig. 124 (South China Sea). DIAGNOSIS: Vomerine teeth 4-10; lower jaw teeth 31-286 in specimens over 16 mm SL; dorsal midline of esca with an anteriorly or anterodorsally directed papilla; D usually 6; A usually 5. DESCRIPTION: Posterior escal appendage (PA) laterally compressed, crescentic, almost equaling escal length, darkly pigmented in specimens over 98 mm SL. Two small unpigmented papillae on either side of esca just in front of PA. Papillae also in dorsal and anterior midline of esca; papillae simple and unpigmented in specimens less than 98 mm SL, bifurcated and medially pigmented in larger fish. Measurements in percent SL (12 specimens, 16-159 mm SL, from Pietsch, 1972a): head length 32-46; head width 16-40; head depth 26-46; lower jaw 28-54; upper jaw (premaxilla) 23-32; illicium 25-36. Counts: D 5-7, usually 6; A 4-6, usually 5; P 17-21; vomerine teeth 4-10. DISTRIBUTION: All three major oceans in tropical and temperate waters. Adults meso- and bathypelagic. REMARKS: Large female D. longicornis may be identifed on the basis of vomerine and lower jaw tooth counts alone. In these counts, specimens less than 25 mm may be confused with D. pullatus, in which case escal morphology will be required. Thus, small specimens with escal damage from our area may not be identifiable. See Pietsch (1972a, fig. 3) for regression of jaw tooth counts. MATERIAL (both females): ISH 1205/71 (98 mm SL); central Cape Basin; 37E08'S, 05E23'E; BT; 0-2200 m; WALTHER HERWIG sta. 412/71; 21 Mar. 1971. ISH 1098/71 (83 mm SL); central Cape Basin; 39E19'S, 03E15'W; BT; 0-2000 m; WALTHER HERWIG sta. 406/71; 19 Mar. 1971. Neither seen. Dolopichthys pullatus Dolopichthys pullatus Regan & Trewavas, 1932: 79, fig. 123, pl. 3, fig. 1 (Molucca Sea, Indonesia). Pietsch, 1972a: 6, figs. 1, 3, 4. Pietsch, 1974, figs. 23, 35, 39F, 44, 51D, 56C. Bertelsen & Pietsch, 1977: 185. Pietsch & Seigel, 1980: 394. Bertelsen & Pietsch, 1983: 87, fig. 8. Paxton et al., 1989: 291. Paulin et al., 1989: 139. Bertelsen, 1991: 501. Dolopichthys longicornis (not of Regan and Trewavas, 1932; in part): Bertelsen, 1951: 96, 100, 101, figs. 53D, 57. Dolopichthys allector (not of Garman, 1899; in part): Pietsch, 1972a: 35, fig. 24. DIAGNOSIS: Vomerine teeth 4-14; lower jaw teeth 44- ca. 600; dorsal midline of esca with a posteriorly or posterodorsally directed papilla; D usually 6; A usually 5. DESCRIPTION (Single female, 63 mm SL): Pigmented, anteriorly directed papilla near anterior margin of esca; another, smaller papilla at base of unpigmented, blade-like posterior escal appendage (PA). Head as long as deep, roughly triangular. Jaws equal anteriorly, rictus extending beyond orbit. Symphysial spine of lower jaw well developed. Sphenotic spines large, protruding. No skin spines. Posterior margin of opercle deeply bifurcated. Measurements in percent SL: head length 41; head width 25; head depth 41; upper jaw 34; lower jaw 42; interorbital width 24; longest upper jaw tooth 2; longest lower jaw tooth 2; illicium 29; esca length 4; esca width 2.5; gill slit 9; pectoral-base width 4; pectoral-fin 13; predorsal 71; preanal 73; body depth at anal-fin origin 13; greatest body depth 44; dorsal-fin base 16; anal-fin base 11; longest dorsal ray 18; longest anal ray 16; least caudal peduncle depth 7; peduncle length 15. Counts: D 7; A 5; P18; upper jaw teeth 184; lower jaw teeth 282; vomerine teeth 5. DISTRIBUTION: All three major oceans in tropical and temperate waters. Adults meso- and bathypelagic. REMARKS: May be confused with D. longicornis at small sizes. See Remarks under that species (above). MATERIAL: SAM 29238 (female, 63 mm SL); off Port St. Johns, South Africa; 31E33.1'S, 30E18.2'E; MWT; 0-1050 m; sta. SM 195; 19 Jun. 1979. Oneirodes Lütken, 1871 DIAGNOSIS: Females with skin spinules present, scattered; skin not extending past caudal base; rictus of mouth extending beyond eye; vomerine teeth present; pterygiophore of illicium emerging on snout, its anterior end exposed; lower jaw with well developed symphysial spine; sphenotic spines well developed; pectoral lobe shorter than longest pectoral rays; posterior margin of opercle deeply bifurcate; A 4, rarely 5. Other, osteological, characters found in Pietsch (1974). Females reach 213 mm SL. Males nonparasitic, with area between anterior nostrils and between posterior nostril and eye pigmented; area between anterior and posterior nostrils unpigmented; caudal peduncle without subdermal pigment, except in O. melanocauda (known only from larvae taken outside our area); olfactory lamellae 6-12; upper denticular teeth 6-17; lower denticular teeth 7-28. REMARKS: A single male Oneirodes sp. is known from our waters. Apparently, within the genus Oneirodes, only the male of the larval taxon O. melanocauda has been identified (Bertlsen, 1951; Pietsch, 1974). We provide here a brief description. Head longer than deep, roughly triangular. Jaws subequal, toothless. Body elongate, laterally compressed. Posterior nostril very large, longer than eye. Eye rounded, with small aphakic space. A series of 7 small dermal humps before dorsal fin in midline. Subopercle large and broadly rounded dorsally; inner surface of subopercle unpigmented. Skin between nostrils white, between eye and posterior nostril pigmented (brown in alcohol); no subdermal pigment on caudal peduncle. Measurements in percent SL: head length 32; head depth 29; postorbital 28; upper jaw 18; lower jaw 19; greatest diameter of posterior nostril 9; snout 15; eye diameter 6; pectoral base width 7; pectoral-fin 15; predorsal 68; preanal 70; body depth at anal-fin origin 20; greatest body depth 23; dorsal-fin base 11; anal-fin base; caudal peduncle depth 11; peduncle length 19. Counts: D 5; A 4; P 17; nasal lamellae 10; upper denticular teeth 10; lower denticular teeth 15; vomerine teeth absent. MATERIAL: Oneirodes sp.: SAM 28760 (male, 13.6 mm SL); off Port St. Johns, South Africa; 31E 40.2'S, 30E03.9'E; MWT; 0-1333 m; sta. SM 201; 20 Jun. 1979. KEY TO SPECIES (Females) 1a. Esca with well developed lateral appendages (LA) 2 1b. Esca without lateral appendages; anterior (AA) and medial (MA) appendages filamentous, posterior appendage (PA) simple; terminal papilla (TP) enlarged (Fig. 12B) O. eschrichtii 2a. Medial appendages (MA) well developed (Figs. 12C & D) 3 2b. MA absent; many low, simple filaments in dorsal midline of escal bulb, number increasing with growth. O. anisacanthus 3a. AA with an unpigmented, bulbous tip with 2 terminal filaments and no filaments in middle area; PA longer than escal length (Fig. 12C) O. kreffti 3b. AA with an unpigmented, conical tip, a lateral bulb, and 2-3 unbranched terminal filaments; middle area of AA with few simple or branched filaments; PA much shorter than escal length (Fig. 12D) O. whitleyi Oneirodes anisacanthus Dolopichthys anisacanthus Regan, 1925: 562 (North Atlantic). Regan, 1926: 29. Regan & Trewavas, 1932: 72, fig. 105, pl. 2, fig. 2. Maul, 1961: 129, figs. 19-21 (in part). Dolopichthys heteracanthus Norman, 1930: 353 (in part; not of Regan, 1926). Oneirodes anisacanthus Pietsch, 1974: 37, 54-57, figs. 73, 74. Bertelsen & Pietsch, 1977: 174, fig. 2. Bertelsen, 1986: 1394, text figs. Bertelsen, 1991: 504. DIAGNOSIS: Lateral escal appendages (LA) well developed, elongate over 50 mm SL; anterior escal appendage (AA) longer than posterior appendage (PA) at all sizes, internally pigmented proximally in larger specimens; median escal appendages (MA) absent; D 6-7; A 4; P 14-19; upper jaw teeth 21-53; lower jaw teeth 26-54; vomerine teeth 5-8. DESCRIPTION: Damaged female. Head massive, roughly triangular. Jaws strong, rictus of mouth extending well beyond eye. Body stout, rapidly tapering posteriorly. Esca with elongate LA, originating somewhat anteriorly; PA short, swollen distally; AA lightly pigmented proximally about 1/3 its length. Eye minute, but head and body skinned. Sphenotic spines well developed. Opercle deeply bifurcate, posterior margin of subopercle gently rounded. Pectoral lobe slightly longer than high. Measurements in percent SL: head length 44; head width 27; upper jaw 32; lower jaw 46; interorbital width 8; longest upper jaw tooth 4.5; longest lower jaw tooth 6; illicium 26; esca length 6; esca width 4; gill slit 7; pectoral base width 7; pectoral-fin 21; predorsal 75; preanal 84; body depth at anal-fin base 10; dorsal-fin base 11; anal-fin base 10; longest dorsal ray 22; longest anal ray 20; least caudal peduncle depth 11; peduncle length 13. Counts: D 5; A 4; P 14; upper jaw teeth 34; lower jaw teeth 32; vomerine teeth 7. DISTRIBUTION: Both sides of the North Atlantic from Iceland to the west coast of South Africa. Adults meso- and bathypelagic. MATERIAL: SAM 31583 (female, 51 mm SL); Cape Canyon; 33E38.1'S, 17E24.1'E; MWT; 0-990 m; sta. A6996-060; 7 Mar. 1988. Oneirodes eschrichtii Oneirodes eschrichtii Lütken, 1871: 56, pl. 11 (off Greenland). Pietsch, 1974: 37, 44-52, figs. 65-70, frontispiece. Bertelsen & Pietsch, 1977: 174. Pietsch & Seigel, 1980: 383, fig. 3. Masuda et al., 1984: 106, pl. 346. Bertelsen, 1986: 1395, text figs. Munk, 1988: 93, figs. 1-19. Bertelsen, 1991: 504. Munk, 1998: 176, figs. 2-4. Oneirodes megaceros Holt & Byrne, 1908: 93-95 (NE Atlantic). Regan, 1926: 29 (in part). Dolopichthys obtusus Parr, 1927: 16-18, fig. 5 (off Bermuda). DIAGNOSIS: No lateral escal appendages (LA); anterior escal appendage (AA) with short filaments, increasing in number with size; posterior escal appendage (PA) simple (unbranched); median escal appendages (MA) of numerous, hair-like filaments; D 5-7; A 4; P 15-19; upper jaw teeth 24-50; lower jaw teeth 29-56; vomerine teeth 4-8. DESCRIPTION: Damaged female. Head massive, roughly triangular, as long as deep. Body globose, tapering rapidly at origins of unpaired fins. Skin spinules present, sparse over patches of remaining skin. Antertor escal appendage (AA) base thick, pigmented, with 4 distal filaments; MA highly filamentous, PA rather short, slender, unbranched; TP with enlarged, unpigmented base. Eye minute, orbit distorted due to loss of tissue. Sphenotic spines well developed. Opercle deeply bifurcate. Posterior margin of subopercle gently convex dorsally, becoming straight at midsection. Pectoral lobe stout, about as long as deep. Measurements in percent SL: head length 42; head width 21; head depth 43; postorbital 20; upper jaw 35; lower jaw 42; interorbital width 7; longest upper jaw tooth 4; longest lower jaw tooth 5; illicium 46; esca length 7; esca width 3; gill slit 7; pectoral base width 9; pectoral-fin 21; predorsal 72; preanal 82; body depth at anal-fin origin 25; greatest body depth 53; dorsal-fin base 11; anal-fin base 6; longest dorsal ray 22; longest anal ray 19; least caudal peduncle depth 11; peduncle length 16. Counts: D 7; A 4; P 15; upper jaw teeth 32; lower jaw teeth 30; vomerine teeth 5. DISTRIBUTION: All three major oceans in tropical to cold temperate waters. Adults meso- and bathypelagic. REMARKS: Munk (1988, 1998) described the ultrastructure of the escal light organ and guides in O. eschrichtii. See Pietsch (1974) or Bertelsen (1991) for full synonymies of this species, especially the numerous names used by Regan and Trewavas (1932). MATERIAL: SAM 29461 (female, 54 mm SL); Namibia; sta. 39; SFRI crab survey; Aug. 1982 (no other data). Oneirodes kreffti Oneirodes kreffti Pietsch, 1974: 37, 57-58, figs. 75, 76 (W. of South Africa). Bertelsen & Pietsch, 1977: 175, fig. 3 (not fig. 2). Bertelsen & Pietsch, 1983: 84, fig. 5. Pietsch, 1986a: 375, fig. 106.1. Paxton et al., 1989: 291. DIAGNOSIS: Anterior escal appendage (AA) with distal filament bifurcated at base, arising from terminal bulbous swelling; median escal appendage (MA) elongate, filamentous, with numerous branches; lateral escal appendage (LA) elongate, simple; posterior escal appendage (PA) simple; D 5-6; A 4; P 16-18; upper jaw teeth 31-55; lower jaw teeth 39-50; vomerine teeth 6-8. DESCRIPTION: Four females known, 21-101 mm SL. Head large, somewhat triangular. Mouth large, rictus extending to below eye. Symphysial spine of lower jaw strong. Body not globose, tapering rapidly posterior to unpaired fin bases. Esca with LA the longest appendage; branchings of MA numerically increasing with size; each MA may bifurcate basally; PA decreasing in length with increasing size. Eye minute, rounded. Sphenotic spines well developed. Subopercle with posterior margin rather straight (Pietsch, 1974, fig. 76). Pectoral lobe about as long as high. Measurements in percent SL: head length 39-64; head depth 44-51; lower jaw 46-52; upper jaw (premaxilla) 31-36; illicium 22-24. DISTRIBUTION: Cape Basin and southern Indian Ocean; probably circumglobal in the southern hemisphere, at least. Apparently mesopelagic, one specimen was taken in a closing net at 150-635 m. REMARKS: Originally described on the basis of three specimens, 21-53.5 mm SL, a fourth at 101 mm SL was reported by Bertelsen and Pietsch (1977) from SE of Mauritius. Pietsch (1986a) mistakenly gave the latter as a 75 mm specimen from off Sydney, Australia, perhaps confusing O. kreffti with the species below. MATERIAL (both females): ISH 1536/71 (50 mm SL, holotype); central Cape Basin; 30E04'S, 05E22'E; MWT; 0-500 m; WALTHER HERWIG sta. 431/71; 31 Mar. 1971. ISH 1463/71 (53 mm SL, paratype); central Cape Basin; 33E00'S, 07E50'E; MWT; 0-2000 m; WALTHER HERWIG sta. 427/71; 30 Mar. 1971. Neither seen. Oneirodes whitleyi Oneirodes whitleyi Bertelsen & Pietsch, 1983: 85, fig. 7 (off eastern Australia). Paxton et al., 1989: 291. DIAGNOSIS: Anterior escal appendage (AA) with black internal tube, distal tip with 2-3 unbranched filaments and a conical, bulbous protuberance; middle area of AA with few simple or bifurcate filaments; lateral escal appendages (LA) branching distally; medial escal appendages (MA) with numerous filaments; posterior escal appendage (PA) simple, very short, mostly unpigmented; D 5-6; A 4; P 16-17; upper jaw teeth 16-52; lower jaw teeth 24-41; vomerine teeth 4-8. DESCRIPTION: Three females. Head massive, roughly triangular, as long as deep. Jaws subequal, lower projecting beyond upper; rictus of mouth extending to below eye. Symphysial spine of lower jaw well developed. Body short, rapidly tapering posteriorly at origins of unpaired fins. Skin spinules microscopic. Esca with AA tip conical, a bifurcating filament at its end and a bulbous filament below this; 2-4 unpigmented filaments in middle of AA, some bifurcate; MA unpigmented, with numerous filaments; LA unpigmented, branching distally, about 60% escal length in smallest, 3 times escal length in largest; PA digitiform, with thin line of black internal pigment; largest specimen with intact PA, ca. 1/3 escal length. Sphenotic spines rather low, blunt. Opercle deeply bifurcate. Posterior margin of subopercle very gently convex, almost straight at mid-height. Pectoral lobe longer than high. Measurements in percent SL: head length 35-50; head width 21-22; head depth 49-53; upper jaw 34-36; lower jaw 38-47; interorbital width 9-13; longest upper jaw tooth 4-4.5; longest lower jaw tooth: 5; illicium 53-56; esca length 4-8; esca width 3-5; pectoral base width 9-10; pectoral-fin 18-21; predorsal 74-82; preanal 83-86; body depth at anal-fin origin 18-29; greatest body depth 49-62; dorsal-fin base 9-13; anal-fin base 7-8; longest dorsal ray 12-22; longest anal ray 9-22; least caudal peduncle depth 12-13; peduncle length 9-17. Counts: D 5-6; A 4; P 16-17; upper jaw teeth 23-52; lower jaw teeth 28-41; vomerine teeth 2-8. DISTRIBUTION: West coast of South Africa and eastern Australia; probably circumglobal in the southern hemisphere. Mesopelagic. MATERIAL (all females): SAM 31580 (31 mm SL); Cape Canyon; 33E48.4'S, 17E22.2'E; BT, 0-900 m; sta. A6988-060; 5 Mar. 1988. RUSI 61310 (126 mm SL); Cape Point Valley; 34E43'S, 17E58'E; BT, 0-924 m; FRIDTJOF NANSEN sta. AN0055; 24 Jan. 2000. RUSI 61316 (25 mm SL); Agulhas Bank; 35E59'S, 19E26'E; BT, 0-938 m; FRIDTJOF NANSEN sta. AN0035; 21 Jan. 2000. FAMILY THAUMATICHTHYIDAE DIAGNOSIS: Females with upper jaw protruding well in front of lower; upper jaws (premaxillae) widely separated at symphysis, connected by broad, elastic membrane which allows jaw to rotate up and outward; esca with 1 or 3 denticles; esca on long illicium (Lasiognathus) or inside mouth (Thaumatichthys); upper ramus of opercle poorly ossified, with 1-6 long, thin extensions; branchiostegal rays 6; D 5-7; A 4-5; P 14-20; C 9. Other, mostly osteological, characters are found in Bertelsen and Struhsaker (1977). This rare family contains 2 genera, Lasiognathus Regan, 1925, with 4 species, and Thaumatichthys Smith and Radcliffe, 1912, with 3. Lasiognathus is known in our area from a 129 mm SL female, the largest for the genus so far collected. Lasiognathus adults are meso- and bathypelagic, but Thaumatichthys is benthic at upper slope to abyssal depths. Larvae are epipelagic. Very few thaumatichthyid males are known; none have been referred to Lasiognathus. The few known males of Thaumatichthys are all immature, thus it is unknown if sexual parasitism occurs. The genus Lasiognathus was reviewed by Nolan and Rosenblatt (1975) and Bertelsen and Pietsch (1996). Bertelsen and Struhsaker (1977) revised Thaumatichthys and briefly mentioned a 4 mm larval male, which is probably T. pagidostomus from off Mozambique. As it cannot be positively identified, we do not treat Thaumatichthys here. Lasiognathus Regan, 1925 DIAGNOSIS: Females with head long (more than 60% SL) and narrow; pterygiophore of illicium exposed between frontal bones (whole lure inserted dorsally on head, i.e., in normal position); esca with 3 bony hooks emanating from distal appendage; skin spinules absent; D 5; A 5; P 16-20; vomerine teeth absent. Males unknown. Lasiognathus intermedius Lasiognathus intermedius Bertelsen & Pietsch, 1996: 406, fig. 4 (Agulhas Bank, South Africa). DIAGNOSIS: Distal escal appendage (DA) with slender prolongation emerging from base of middle escal hook; prolongation without serrations or filaments; posterior escal appendage (PA) compressed distally; no fleshy ridge along anterior surface of escal bulb; illicial length 41-53% SL. DESCRIPTION: Head dorsoventrally depressed, snout elongate and more depressed anteriorly. Upper jaw greatly produced beyond lower, cleft of mouth reaching eye or nearly so. Sphenotic and articular spines well developed. Illicium lying a in deep groove on head. Esca with 3 pigmented hooks (middle hook the larger) emanating from a distal appendage (DA) and a fleshy, unpigmented prolongation distal to hooks (Fig. 13). Pectoral-fin lobe relatively short and broad. Scales and denticles (skin spinules) absent. Premaxillae separated anteriorly, connected by membrane (no ligament). Teeth in upper jaw fang-like, shorter in lower jaw (most of these missing in our damaged specimen). Measurements in percent SL: head length 61; upper jaw 38; lower jaw 32; snout 52; eye diameter 2; longest upper jaw tooth 6; longest lower jaw tooth 2.5; illicium 53; esca length 2; esca width 1; length DA 4.5; length of DA prolongation 2; middle median hook 3; gill slit 3; pectoral-base width 2; pectoral-fin 7; predorsal 81; preanal 82; dorsal-fin base 5; anal-fin base 4; longest dorsal ray 13; longest anal ray 11; least caudal peduncle depth 6; peduncle length 13. Counts: D 5; A 5; P 17; upper jaw teeth 78; lower jaw teeth (?); vomerine teeth absent. DISTRIBUTION: Seven specimens known from the northwestern and southeastern Atlantic and eastern tropical Pacific. All from open nets fished into bathypelagic depths. MATERIAL: SAM 32819 (female, 127 mm SL, holotype); eastern Agulhas Bank; 35E09.8'S, 23E37.0'E; MWT; 975-1090 m in water 1055-1126 m; A14120-107; 20 Oct. 1992. FAMILY CENTROPHRYNIDAE DIAGNOSIS: Females with subopercle as long as opercle, and with anterior spine on lower half; opercle bifurcate, upper ramus reduced; jaws equal anteriorly, teeth not enlarged; well developed symphysial spine on lower jaw; large oval pit just before eye in specimens over 40 mm SL; a short fleshy hyoid barbel present in larvae and juveniles less than 50 mm SL; nostrils paired, tubular; no sphenotic spine; pectoral lobe shorter than longest pectoral ray; skin spinules present; branchiostegal rays 6; D 6-7; A 5-6; P 15-16; C 9; upper jaw teeth 32-153; lower jaw teeth 23-96; vomerine teeth 0-9. Males with globose head, more triangular in metamorphosed specimen; hyoid barbel reduced to a nubbin in metamorphosed specimen; subopercle as long as opercle, with anterior spine on lower half; anterior nostrils directed forward, somewhat larger than posterior nostrils; few larval teeth in lower jaw of both metamorphosing specimens; skin pigmented between anterior and posterior nostrils and on snout between anterior nostrils, except in larva; eyes rounded, without aphakic space; olfactory lamellae 7-8; upper denticular teeth 3; lower denticular teeth 4; fin rays as for females. This family contains a single species, Centrophryne spinulosa, originally described in the Oneirodidae. Females reach 230 mm SL (Pietsch, 1976), but only 4 males are known, one larva (from our area), 2 metamorphosing specimens and a free-living, metamorphosed individual. Sexual parasitism may thus not occur, but in a bizarre case of mistaken mate identity, Pietsch and Nafpaktitis (1971) found a male Melanocetus johnsonii that had attached itself to the upper lip of a female Centrophryne spinulosa. Adult Centrophryne are meso- and bathypelagic, larvae epipelagic. Descriptive information on this family is found in Bertelsen (1951, 1983) and Pietsch (1972b, 1976). The larval male from our area was only briefly mentioned by Bertelsen (1951) and Pietsch (1972b), but was examined by the senior author in June 1998 for the description below. Centrophryne spinulosa Centrophryne spinulosa Regan & Trewavas, 1932: 84, pl. 4, fig. 2 (Gulf of Panama and north of New Guinea). Bertelsen, 1951: 126, figs. 84- 87. Grey, 1956: 261 (syn., males). Pietsch & Nafpaktitis, 1971: 322, figs. 1-4. Pietsch, 1972b: 17, figs. 1-25. Pietsch, 1976: 786. Pietsch & Seigel, 1980: 395. Bertelsen, 1983: 313, figs. 2-3. Bertelsen, 1984: 329, fig. 168F. Bertelsen, 1986: 1401, text figs. Bertelsen, 1991: 509. Rhynchoceratias leucorhinus (not of Regan, 1925): Regan, 1926: 44 (in part). DIAGNOSIS: As for family (above). DESCRIPTION: Larval male, 7.5 mm SL. Head and body globose owing to encapsulating, gelatinous flesh. Hyoid barbel fleshy, unpigmented, about as long as eye. Mouth terminal, jaws subequal, larval teeth present in both jaws. Denticular bones and nasal lamellae not developed. Eye rounded, no aphakic space. Pigment pattern described by Bertelsen (1951) not evident today (specimen alizarin stained and stored in 70% glycerol). Measurements in percent SL: head length 32; head depth 45; postorbital 17; upper jaw 16; lower jaw 19; snout 13; eye diameter 10; pectoral base width 13; pectoral-fin 16; predorsal 73; preanal 76; body depth at anal-fin origin 27; greatest body depth 62; dorsal-fin base 13; anal-fin base 9; longest dorsal ray 20; longest anal ray 21; caudal peduncle depth 12; peduncle length 16. Counts: D 7; A 5; P 16. DISTRIBUTION: Tropical and warm temperate regions of all three major oceans. MATERIAL: ZMUC P92153 (male, 7.5 mm SL); Mozambique Channel; 25E19'S, 36E13'E; S150 net; 50 m.w.o. (= approx. 30 m depth); DANA sta. 3964, cr. 5; 15 Jan. 1930. FAMILY CERATIIDAE DIAGNOSIS: Females with 2-3 caruncles on dorsal midline in front of dorsal fin; jaws equal anteriorly, mouth oblique in adults, teeth not enlarged; eye minute; sphenotic spines absent; skin spinules strong, close-set; second cephalic ray and esca present, becoming rudimentary in adults; posterior end of illicial pterygiophore ("basal bone") emerging from dorsal midline in front of caruncles, and retractable beneath skin (Bertelsen, 1951, fig. 4); illicium about half head length (Ceratias), or minute remnant about twice escal length or less (Cryptopsaras); esca without denticles; branchiostegal rays 6; D 3-5; A 4; C 8 (Cryptopsaras) or C 9 (Ceratias, but lowermost a small remnant; 8 rays visible without dissection). Free-living male juveniles with 2 upper denticular teeth, fused at base, and articulating with illicial pterygiophore (basal bone); 2 pairs of lower denticular teeth; eyes large, directed laterally, with aphakic space; caruncles absent; skin naked and unpigmented (spiny and pigmented in attached parasitic males). Counts as per females. Other, mainly osteological, characters of both sexes given by Pietsch (1986b). The seadevils are a family of only four species in two genera, Ceratias Kröyer and Cryptopsaras Gill, and all four occur in our area. Females of Ceratias holboelli are the largest deep-sea angler, reaching 145 cm TL (Pietsch, 1986a); males are parasitic and reach 118 mm. Females have a second cephalic ray (lure) with an esca. The posterior esca is withdrawn under the skin and loses its connection with the second ray at metamorphosis (Bertelsen, 1951). Females also have additional bacterial light organs on their backs (caruncles) which are situated at the tips of modified dorsal-fin rays (the caruncles degenerate in Ceratias at about 40 cm SL, but are retained by Cryptopsaras). Caruncles secrete a whitish-yellow luminescent fluid which appears to be an anti-predator, startle substance (Marshall, 1979). Early stages in the development of the light organs of Cryptopsaras were studied by Munk and Herring (1996). In Ceratias an accessory escal gland develops around the illicium and part of the esca. It is not light-generating, but may supplement the function of the lure in some way, such as secreting a scent (Munk, 1992). One of the commonest ceratioids, Cryptopsaras has been taken in nets that reached from 75 to 4000 m, the majority of specimens, however, were taken between 500 and 1250 m. Pietsch (1986b) provided a review of the family. Larval development and metamorphosis were described by Bertelsen (1951; 1986). Fishes and crustaceans are the main prey of seadevils. INDETERMINATE: The following females of Ceratias lack the illicial apparatus and variously have or lack vomerine teeth. They are thus indeterminable (Pietsch, 1986b): SAM 34771 (97 mm SL); NW of Lüderitz, Namibia; FRIDTJOF NANSEN sta. 1386; 25E38'S, 13E32'E; 0-644 m; 30 Oct. 1992. RUSI 2723 (510 mm SL); off Saldanha Bay; commercial trawler, 16 Oct. 1972. RUSI 38217 (215 mm SL); off St. Helena Bay; sta. A13033; 32E24'S, 16E14'E; 0-1012 m; 15 Feb. 1992. SAM 29607 (334, 373, 453 mm SL); whale stomach at Donkergat Whaling Station; P. Best, 1966. SAM 31584 (40 mm SL); Cape Canyon; sta. A6997-060; 33E43.5'S, 17E21.5'E; 0-983 m; 7 Mar. 1988. SAM 31586 (105 mm SL); Cape Canyon; sta. A6999-060; 33E40.0'S, 17E25.8'E; 0-923 m; 8 Mar. 1988. SAM 33146 (129 mm SL); S. of Childs Bank; sta. A9827-079; 31E14.6'S, 15E35.5'E; 0-796 m; 26 Jan. 1990. KEY TO GENERA (Females) 1a. Two club-like caruncles in dorsal midline anterior to dorsal fin; C 9; illicial length much greater than escal length Ceratias 1b. Three caruncles in dorsal midline; C 8; illicium reduced to small remnant almost entirely enveloped by escal tissue Cryptopsaras Ceratias Krøyer, 1845 DIAGNOSIS: Illicial length 14-38% SL; two luminescent caruncles in dorsal midline (minute in specimens over 40 cm SL); subopercle without anteriorly-directed spine; caudal-fin rays 9, the lowermost a small, fused remnant. As with most ceratioid genera, species-level characters other than escal morphology are difficult to find. Pietsch (1986b) found over 60% of the Ceratias he studied had lost their lures, thus impeding identification. Key below taken from Pietsch (op. cit.). Free-living juvenile males presently unknown in our area. KEY TO SPECIES (Females) 1a. Esca with one or no distal appendage (DA); vomerine teeth present or absent 2 1b. DA paired; vomerine teeth present C. tentaculatus 2a. Esca with a DA; vomerine teeth almost always present in specimens less than about 80 mm SL, rarely present in larger individuals; illicial length 15-38% SL (see Pietsch, 1986b, fig. 3 for regression) C. holboelli 2b. Esca without DA; illicial length 14-29% SL; vomerine teeth always absent C. uranoscopus Ceratias holboelli Ceratias Holbölli Krøyer, 1845: 638, 648 (off southern Greenland). Bertelsen, 1943: 185, figs. 1-4. Ceratias holboelli: Günther, 1887: 53. Regan, 1926: 34, fig. 20. Regan & Trewavas, 1932: 96. Bertelsen, 1951: 133, figs. 88-92, frontispiece. Grey, 1956: 261. Penrith, 1967: 185. Pietsch, 1976: 781, 787. Bertelsen & Pietsch, 1983: 89, fig. 10. Bertelsen, 1984: 329, fig. 168E. Masuda et al., 1984: 108, pl. 93F. Pietsch,1986a: 374, text fig. Pietsch, 1986b: 481, 484, 485, figs. 2, 3, tab. 1. Bertelsen, 1986: 1403, text fig. Paulin et al., 1989: 137. Paxton et al., 1989: 294. Bertelsen, 1991: 510. Munk, 1992: 33, figs. 2-3. Nielsen & Bertelsen, 1992: 62, text fig. Ceratias holbølli: Koefoed, 1944: 10, pl. 2, fig. 2. Reganichthys giganteus Bigelow & Barbour, 1944: 10, pls. 4-6 (off Portland, Maine, USA). DIAGNOSIS: Esca with single distal appendage which may develop side branches (filaments) at large sizes; vomerine teeth usually lost by about 80 mm SL; illicial length 14-38% SL, longer than in other species at comparable sizes over about 65 mm SL (Pietsch, 1986b, fig. 3). Males unknown. DESCRIPTION: Metamorphosed females with stout head, becoming deeper than long with increasing size. Head and body fusiform. Jaws subequal, upper jaw slightly overhanging lower when mouth closed; mouth strongly oblique, more horizontal in larvae; rictus of mouth not reaching below eye. Eye minute, rounded. Skin spinules small, close-set, needle-like in young, becoming sparse and thorn-like in largest adults. Proximal half to two-thirds of esca pigmented, distal part ovoid, becoming stouter with increasing size. Escal pore at apex of bulb, on a raised papilla. Counts: D 4 (rarely 3); A 4; P 16-19; vomerine teeth 2-6 in specimens less than 80 mm SL, 0-4, usually lost, in larger fish. DISTRIBUTION: Tropical to subpolar regions of all three major oceans. Meso- and bathypelagic. MATERIAL: IOAN uncat. (female); off Durban; 29E54'S, 31E51'E. No other data (Pietsch, 1986b). Not seen. Ceratias tentaculatus Mancalias tentaculatus Norman, 1930: 355, fig. 45 (South Atlantic, NE of Bouvet Isl.). Regan & Trewavas, 1932: 100, fig. 158D. Bertelsen, 1943: 201. Clarke, 1950: 13, figs. 4B, 5 as syn. of C. holboelli). Mancalias bifilis Regan & Trewavas, 1932: 100, fig. 158C, pl.6, fig. 1 (off New Zealand). Clarke, 1950: 13, figs. 1B, 4A, 5 (syn. of Ceratias holboelli). Ceratias holboelli tentaculatus: Bertelsen, 1951: 133. Ceratias holbölli (not of Krøyer, 1845): Clarke, 1950: 1-32, figs. 1A, 2B, 3, 4C. Ceratias holboelli (not of Krøyer, 1845): Penrith, 1967: 185. Ceratias tentaculatus: Bertelsen & Pietsch, 1984: 43, fig. 1. Pietsch, 1986a: 374, text figs. Pietsch, 1986b: 481, fig. 1. Paulin et al., 1989: 137. Pietsch, 1990: 210, text figs. Smale et al., 1995: 70, pl. 36A (tentative; RUSI 38217). DIAGNOSIS: Distal escal appendages (DA) paired; illicial length 19-38% SL; vomerine teeth present. DESCRIPTION: General description as for C. holboelli (above). Escal distal appendages (DA) usually proximally pigmented; our largest specimen without appendage pigmentation or bifurcations (Fig. 14B). Measurements in percent SL: head length 22-36; head width 12-17; head depth 37-41; postorbital 6; upper jaw 16; lower jaw 19; snout 14-20; eye diameter 2-6; interorbital width 4; longest upper jaw tooth 1; longest lower jaw tooth 1; illicium 31-38; esca length 1; esca width 0.5-6; gill slit 6-7; pectoral base width 5; pectoral-fin 4; predorsal 72; preanal 74; body depth at anal-fin origin 18; greatest body depth 36-43; dorsal-fin base 11; anal-fin base 9; longest dorsal ray 22; longest anal ray 20; least caudal peduncle depth 8-9; peduncle length 16. Counts: D 4; A 4; P 17; upper jaw teeth 20-47; lower jaw teeth 32-52 (buds in SAM 29954); vomerine teeth 1. DISTRIBUTION: Circumglobal in southern hemisphere. Meso- and bathypelagic. MATERIAL (all females): UW 22321 (460 mm SL); off Childs Bank; sta. A11416-090; 30E48'S, 15E10'E; MWT; 0-758 m; 10 Mar. 1991. SAM 24829 (468 mm SL); Donkergat whaling station (Saldanha Bay); whale stomach; Mar. 1966; P. Best. SAM 31592 (27 mm SL); Cape Point Valley; sta. A7033-060; 34E46.6'S, 18E02.5'E; MWT; 0-702 m; 13 Mar. 1988. SAM 34715 (552 mm SL); so. of Danger Point, Cape; ca. 35ES, 19EE; commercial trawler; 0-350 m; 14 Dec. 1997. SAM 29954 (10.1 mm SL); off Durban; sta. SM 138; 30E21.3'S, 31E15.7'E; MWT; 0-830 m; 14 May 1977. BMNH 1932.5.3.37 (15 mm SL); off Delagoa Bay, Mozambique; 25E18'S, 36E13'E (not seen). Ceratias uranoscopus Ceratias uranoscopus Murray, in Thomson, 1877: 67, fig. 20 (off NW Africa). Günther, 1887: 52, 54, pl. XI. Pietsch, 1986b: 481, 485-487, figs. 3-5. Bertelsen, 1991: 510. Mancalias uranoscopus: Gill, 1878: 227 (type desig. of Mancalias Gill). Goode & Bean, 1896: 490. Regan, 1926: 37, fig. 21. Parr, 1927: 30. Typlopsaras shufeldti Gill, 1883: 284 (Middle Atlantic Bight, USA). Ceratias shufeldti: Günther, 1887: 52, 54. Mancalias shufeldti: Goode & Bean, 1896: 490, pl. 119, fig. 401. Mancalias xenistius Regan & Trewavas, 1932: 99, fig. 158B, pl. 6, fig. 2 (South China Sea). Mancalias sessilis Imai, 1941: 245, figs. 12, 13 (Sagami Bay, Japan). Mancalias krøyeri Koefoed, 1944: 11, pl. 3, fig. 4 (off Canary Isls.). DIAGNOSIS: No escal appendages; illicial length 14-29% SL; vomerine teeth always absent. DESCRIPTION: General description as for C. holboelli but esca without appendages. Measurements in percent SL: head length 24-31; head width 11-18; head depth 32-41; postorbital 7-13; upper jaw 18-21; lower jaw 21-24; snout 14-18; interorbital width 4-7; longest upper jaw tooth 2; longest lower jaw tooth 2-3; illicium 21-24; esca length 4-5; esca width 1-2; gill slit 6-9; pectoral base width 4-7; pectoral-fin 5-8; predorsal 69-73; preanal 72-74; body depth at anal-fin origin 13-16; greatest body depth 29-49; dorsal-fin base 7-10; anal-fin base 7-9; longest dorsal ray 13-15; longest anal ray 11-13; least caudal peduncle depth 7-9; peduncle length 16-19. Counts: D 4; A 4; P 16; upper jaw teeth 44-80; lower jaw teeth 39-55; vomerine teeth absent. DISTRIBUTION: Tropical to cold temperate regions of all three major oceans. Meso- and bathypelagic. MATERIAL (all females): SAM 34772 (59 mm SL); off Orange Banks, Namibia; FRIDTJOF NANSEN sta. 1088; 27E44'S, 14E32'E; BT; 0-500 m; 29 Apr. 1992. ISH 1260/71 (74 mm SL); Cape Canyon; 34E12'S, 16E35'E; WALTHER HERWIG (not seen). RUSI 43049 (27 mm SL); Agulhas Basin; RS AGULHAS sta. C110/2; 40E20'S, 19E15'E; MWT; 10 July 1993. RUSI 54927 (47 mm SL); off Great Fish R.; sta. A12127-096; 34E00.8'S, 27E10.6'E; BT; 0-1045 m; 19 Oct. 1991. SAM 29955 (65 mm SL); off Port St. Johns; sta. SM 195; 31E33.1'S, 30E18.2'E; MWT; 0-1050 m; 19 June 1979. Cryptopsaras Gill, 1883 DIAGNOSIS: Females with illicium a small remnant about twice escal length or less, mostly covered by escal tissue; three caruncles on back just before dorsal fin; caudal-fin rays 8; anteriorly-directed spine on subopercle. Males with anterior pair of lower denticular teeth longer than posterior pair. This monotypic genus has often been misspelled or unjustifiably emended to Cryptosparas, Cryptosarus, or Cryptosaras in the literature (Pietsch, 1986b). Cryptopsaras couesii Cryptopsaras Couesii Gill, 1883: 284 (Middle Atlantic Bight, USA). Goode & Bean, 1896: 491, pl. 119, fig. 402. Parr, 1927: 29. Bertelsen, 1951: 138, figs. 3, 93-97 (as C. couesi). Smith, 1949: 428, fig. 1229. Grey, 1956: 264. Fast, 1957: 237, fig. 1 (males). Smith, 1965: 428, fig. 1229. Penrith, 1967: 186. Pietsch, 1976: 786, fig. 6. Pietsch & Seigel, 1980: 396. Bertelsen & Pietsch, 1983: 91, fig. 11. Bertelsen, 1984: 329, fig. 168C, D (larvae; as C. couesi). Masuda et al., 1984: 108, pl. 93G. Bertelsen, 1986: 1404, text fig. Pietsch, 1986a: 375, text fig. Pietsch, 1986b: 488, fig. 7. Lloris, 1986: 248, fig. 129. Bertelsen, 1991: 511(syn.). Paxton et al., 1989: 294. Paulin et al., 1989: 136. Munk & Herring, 1996: 517, figs. 1-4 (light organs). Ceratias carunculatus Günther, 1887: 52, 55, pl. 11, fig. D (Japan). Ceratias Couesi Brauer, 1906: 317, pl. 15, fig. 7. Ceratias (Paraceratias) mitsukurii Tanaka, 1908: 18, pl. 2 (with parasitic male; Japan). Cryptosparas couesii (lapsus calami): Regan, 1926: 35, pl. 9. Regan & Trewavas, 1932: 96, fig. 154. Koefoed, 1944: 11. Cryptosparas carunculatus: Regan & Trewavas, 1932: 96, 98, fig. 155. Cryptosparas valdiviae Regan & Trewavas, 1932: 96, 98, fig. 156 (Gulf of Aden). Cryptosparas pennifer Regan & Trewavas, 1932: 96, 98, fig. 157 (off New Zealand). Cryptosparas normani Regan & Trewavas, 1932: 96, 98 (Cape Basin). Cryptopsaras atlantidis Barbour, 1941: 11, 14, pls. 3, 6 (No. Atlantic). Cryptosaras couesii (lapsus calami): Smale et al., 1995: 70, pl. 36B (otoliths). DESCRIPTION: Head massive, deeper than long. Body stout, gradually tapering posteriorly at origins of unpaired fins. Skin spinules needle-like. Esca with distal appendage only on pigmented base; this develops a central filament with few to several side branches. Eye minute, subcutaneous, spectacle smaller than pupil. Lure composed of retractable illicial pterygiophore, minute esca and remnant illicium. Sphenotic spine absent. Subopercle with small, hooked spine on anterior margin ventrally (Bertelsen, 1951, fig. 88B). Pectoral lobe about as long as deep. Gill opening oblique, ventral magin slung forward. Caruncles inserted in triangular pattern; the median the largest. Measurements in percent SL: head length 30-41; head width 11-19; head depth 41-51; postorbital 9-13; upper jaw 19-27; lower jaw 24-31; snout 19-21; eye diameter 2-7; interorbital width 4-9; longest upper jaw tooth: 1-2; longest lower jaw tooth: 1-3; illicium 4-17; esca length 4-6; esca width 3-4; gill slit 8-9; pectoral base depth 5; pectoral-fin 6-9; predorsal 75-78; preanal 81-83; body depth at anal-fin origin 17-29; dorsal-fin base 12-13; anal-fin base 10-11; longest dorsal ray 14-17; longest anal ray 12-17; least caudal peduncle depth 11-15; peduncle length 10.5-13. Counts: D 4; A 4; P 16-17; upper jaw teeth 21-119; lower jaw teeth 19-81; vomerine teeth 6-10. Parasitic males were found attached as follows: SAM 34480, with 40 mm TL male on left cheek under eye; SAM 34481, with 88 and 120 mm TL males on left side by gill slit. DISTRIBUTION: All three major oceans in tropical to subpolar waters. Adults mesopelagic. MATERIAL: Females: IIPB 387/1981 (190 mm SL); off northern Namibia; 26E54'S, 14E08'E; BT; 0-386 m; MUESTREOS; 13 May 1980 (not seen). SAM 34770 (92 mm SL); off Namibia; 21E19'S, 12E52'E; BT; 0-301 m; FRIDTJOF NANSEN sta. 620; 14 Feb. 1991. SAM 29455 (107 mm); off Namibia; BT; BENGUELA sta. 51; 26 Aug. 1982; SFRI crab survey. SAM 34386 (36, 82, 275 mm SL); Cape west coast; commercial BT; ALOE sta. 4; 16 June 1995. SAM 26230 (35 mm SL); SE of Vema Seamount; 33E41'S, 05E54'E; MWT; 0-1000 m; AFRICANA II sta. 3632; 15 Apr. 1965. SAM 23587 (318 mm SL); off Cape Columbine; commercial BT; 0-585 m; Jan. 1963. SAM 24283 (358 mm SL); off Cape Columbine; commercial BT; 4 June 1962. SAM 17772 (156 mm SL); NW of Table Bay; BT; Apr. 1950. SAM 31585 (99 mm SL); Cape Canyon; 33E43.5'S, 17E21.5'E; MWT; 0-983 m; A6997-060; 7 Mar. 1988. SAM 31588 (50 mm SL); Cape Canyon; 33E38.8'S, 17E24.0'E; MWT; 0-814 m; A7004-060; 8 Aug. 1988. SAM 34481 (343 mm SL); off Cape Town; commercial BT; Nov. 1992. SAM 19977 (285 mm SL); off Cape Point; commercial BT; ANEMONE; 1957. SAM 34480 (311 mm SL); off Cape of Good Hope; 34E00'S, 17E20'E; commercial BT; 0-490 m; PROTEA; 10 Nov. 1992. SAM 34482 (198 mm SL); off Cape of Good Hope; 34E30.0'S, 17E45.1'E; BT; 0-916 m; A14662-109; 23 Jan. 1993. SAM 25050 (297 mm SL); 45 n. mi. W. Of Cape Point; BT; 0-585 m; ARUM. SAM 29453 (100 mm SL); 70 n. mi. W. of Cape Point; MWT; 0-600 m; BENGUELA; 24 May 1982. SAM 29236 (9.4 mm SL); off Kei R. mouth; 32E30.8, 29E13.4'E; MWT; 0-916 m; SM 218; 23 Jun. 1979. SAM 29237 (34 mm SL); off Port St. Johns; 32E34.4'S, 29E13.1'E; MWT; 0-670 m; SM 223; 23 June 1979. FAMILY GIGANTACTINIDAE DIAGNOSIS: Females with body and caudal peduncle elongate; illicial pterygiophore ("basal bone") emanating from snout tip; sphenotic spines, vomerine teeth and oral valves absent; jaws subequal; branchiostegal rays 6; D 3-10; A 3-8; C 9 (lowermost caudal-fin ray reduced and embedded in tissue). The whipnose anglers are placed in 2 genera, Gigantactis Brauer, 1902, with 18 species, and Rhynchactis Regan, 1925, with 3. The rare, primarily tropical Rhynchactis is known just outside our area (off Madagascar) from a single female of R. macrothrix Bertelsen and Pietsch, 1998 and is not included here. Six species of Gigantactis have been found in our area. Females of Gigantactis reach about 41 cm SL, and adults of both sexes are primarily bathypelagic. Metamorphosed males are non-parasitic and reach 22 mm SL; none are yet known from our area. Parr (1927) described the first male of Gigantactis but placed it in a new genus, Laevoceratias, family Aceratiidae. Bertelsen (1951) tentatively placed it in the family Diceratiidae. The gigantactinids were last reviewed by Bertelsen et al. (1981) who examined all the then known material and recognized 17 species. Kharin (1984) described G. balushkini from off Japan. Gigantactis Brauer, 1902 DIAGNOSIS: Females with greatest body depth about 25% SL; caudal peduncle slender, depth 5-10% SL in most specimens, peduncle length more than 20% SL; snout pointed; upper jaw slightly overhanging lower; teeth fang-like, retrorse, in several rows in lower jaw; D 4-10; A 4-8; escal bulb present. Other, osteological characters, as well as those for males and larvae, given by Bertelsen et al. (1981). INDETERMINATE: Gigantactis sp., SAM 27810 (2 larval females; 11-17 mm SL); off Durban; sta. SM 148; 30E17.1'S, 31E25.2'E; 0-750 m; 17 May 1977. See Bertelsen et al. (1981: 58). A few characters were given for one of the two larval females, SAM 27810 (Bertelsen et al., 1981, table 18; D and A fin-ray counts reversed, corrected here to D 6, A 5). We confirm the high pectoral-fin count (21) in these specimens. Only two species of Gigantactis have counts that high, G. paxtoni and G. gargantua. All of the latter, known from 7 Pacific and eastern Indian Ocean females, have 6 anal-fin rays. Thus, on the basis of fin-ray counts (Bertelsen et al., table 2) and distributions (above), it is most likely that the larvae of SAM 27810 represent G. paxtoni or perhaps G. macronema (P 17-20). As in other speciose ceratioid genera, few characters are of value in differentiating Gigantactis species other than features of the lure. Maximum illicial lengths are individually variable, but are reached by 30 mm SL. Smaller females will thus have to be identified, if they can be at all, on the basis of the esca and sometimes a few other characters. See Bertelsen et al. (1981) for features identifying the species groups of males. KEY TO SPECIES (Metamorphosed females) 1a. Esca with a distal prolongation (not a dorsal appendage) above bulb 2 1b. Esca with tufts or isolated, long or short filaments; area distal to bulb not prolonged 5 2a. Distal escal prolongation with spinules extending to tip;prolongation black, at least partly, in larger specimens 3 2b. Distal prolongation without spinules 4 3a. Lower jaw teeth in 3 series; a pair of appendages (IA) on illicium below escal bulb (Fig. 15E) G. vanhoeffeni 3b. Lower jaw teeth in 5-6 series; illicial filaments just below esca absent or unpaired G. meadi 4a. Escal base with short median papilla (MP; Fig. 15B); distal prolongation subequal to escal bulb width; escal bulb covered with spinules G. kreffti 4b. Escal base without MP; paired, filamentous posterior appendages (PA) present (Fig. 15D); distal prolongation much longer than escal bulb width; no spinules on esca or distal prolongation G. perlatus 5a. Illicial length less than 120% SL G. elsmani 5b. Illicial length 3 to 4 times SL G. macronema Gigantactis elsmani Gigantactis elsmani Bertelsen, Pietsch & Lavenberg, 1981: 43, tabs. 1, 2, 11, figs. 4E, 41, 42 (So. Atlantic). Bertelsen, 1991: 513. DIAGNOSIS: Illicial length less than 120% SL; escal bulb without distal prolongation, its tip with many short filaments and 2 pairs of long filaments; 2 long, lateral filaments at proximal end of escal bulb; posterior series of lower jaw teeth 5; no prolonged caudal-fin rays. DESCRIPTION (from Bertelsen et al., 1981): Females with body and caudal peduncle elongate. Illicium without filaments, laterally compressed proximally. Esca club-shaped, without distal prolongation. Proximal portion of escal bulb dark, with minute spinules reaching light organ and a long filament on each side. Distal portion of escal bulb bearing about 2 dozen short filaments and 2 lateral pairs of long filaments. Upper jaw teeth in 2-3 series; lower jaw teeth in 5 series, better defined in its posterior half (Bertelsen et al., 1981, fig. 42). Measurements in percent SL: illicial length 93-105 (10 in metamorphosing South African specimen; see below); longest upper jaw tooth 1.1-1.2; longest lower jaw tooth 2.8; longest caudal ray 21-27 (40 in South African specimen). Counts: D 4-5; A 4-5; P 16-18; upper jaw teeth 32; lower jaw teeth 35-56. Males and larvae unknown. DISTRIBUTION: Central Atlantic and north of the Amundsen Sea, Antarctic Ocean. Tentatively recorded from a metamorphosing female taken off South Africa. Three specimens known. MATERIAL: MCZ 51269 (female, 11.5 mm SL); Cape Basin; 34E26'S, 16E11'E; MWT; 710-790 m; ATLANTIS II sta. RHB2251; 2 May 1971. Not seen. Gigantactis kreffti Gigantactis kreffti Bertelsen, Pietsch & Lavenberg, 1981: 29, tabs. 1, 2, 4, figs. 4D, 27-29 (So. Atlantic). DIAGNOSIS: Illicial length less than 120% SL; escal distal prolongation with spines restricted to base (when present) and numerous digitiform filaments laterally; single small papilla (MP) on posteromedial base of escal bulb and another (IP) on illicium just below esca; esca without filaments proximally; lower jaw teeth in 4-5 series in larger specimens; longest lower jaw tooth 2.6-5.2% SL; caudal-fin rays less than 30% SL. DESCRIPTION: Females with body and caudal peduncle elongate (Bertelsen et al., 1981, fig. 27); dorsal-fin origin at about mid-length. Illicium without filaments, proximal portion slightly compressed. Escal bulb roughly pear-shaped, more spherical in juveniles, with short, pale distal prolongation. Prolongation covered with short, pale digitiform filaments, tips of these red in life in 252 mm SL holotype. Filaments increase in number and relative length with size. Spinules on illicium and esca, extending on escal bulb to base of distal prolongation, absent on posteromedial surface. Escal pore enclosed by short tube. Small papillae (MP; IP)) on esca and illicium just below esca (Fig. 15B). Upper jaw teeth in single series, with few teeth laterally. Lower jaw teeth in 5 series posteriorly (Bertelsen et al., 1981, fig. 29). Measurements in percent SL: illicium 69-94; longest upper jaw tooth 1.3-2.5; longest lower jaw tooth 2.6-5.2; longest caudal ray 18-28. Counts: D 7; A 6; P 16-18; upper jaw teeth 7-44; lower jaw teeth 13-47. Males and larvae unknown. DISTRIBUTION: Southeastern Atlantic and off Japan. Four specimens known. MATERIAL: ISH 1262/71 (female, 75 mm SL); Cape Canyon; 34E12'S, 16E35'E; MWT; 0-1550 m; WALTHER HERWIG sta. 417/71; 28 Mar. 1971. Not seen. Gigantactis macronema Gigantactis macronema Regan, 1925: 565 (cen. North Atlantic). Regan, 1926: 38, pl. 11 (illustr. of type; after Regan, 1925). Regan & Trewavas, 1932: 93, 94. Bertelsen, 1951: 150, fig. 101 (illustr. after Regan, 1926). Fitch & Lavenberg, 1968: 135, fig. 74. DIAGNOSIS: Illicium about 3.4 to 4.5 times SL; esca with a short, dusky, spineless distal prolongation bearing 8-20 long filaments; no papillae or proximal filaments on esca; lower jaw teeth in two series posteriorly; caudal-fin rays less than 40% SL; caudal rays free nearly to base. DESCRIPTION: Females with illicium, body and caudal peduncle elongate (Bertelsen et al., 1981, fig. 22). Illicial length independent of SL. Illicial filaments absent except in largest specimen (354 mm SL), in which they cover distal sixth of illicial length; each emanating from a swollen, white base (Bertelsen et al., 1981, fig. 52). Esca darkly pigmented proximal to light organ, with spinules (but none distally). Area surrounding light organ pale, with some dusky pigmentation distally. Dorsal prolongation of esca truncated, boundary with escal bulb obscure in smaller specimens, reducing in length with increasing fish length; prolongation covered with long filaments that increase in number with size and may bifurcate in larger specimens. Escal pore enclosed by short tube. No escal or illicial papillae. Upper jaw teeth in single series; lower jaw with double series posteriorly. Measurements in percent SL: illicium 340-447; longest upper jaw tooth 0.6-1.6; longest lower jaw tooth 1.3-3.4; longest caudal ray 26-37. Counts: D 5-6 A 5-6; P 17-20; upper jaw teeth 3-23; lower jaw teeth 5-31. Males and larvae unknown. DISTRIBUTION: Known from 10 females taken in the North and South Atlantic and central and eastern Pacific. Meso- and bathypelagic. MATERIAL: ISH 1596/71 (232 mm SL); Walvis Ridge; 27E14'S, 02E56'E; MWT; 0-2000 m; WALTHER HERWIG sta. 435/71; 1 Apr. 1971. Not seen. Gigantactis meadi Gigantactis meadi Bertelsen, Pietsch & Lavenberg, 1981: 33, tabs. 1, 2, 6, figs. 4B, 18, 20, 23 (mislabelled G. "vahoeffeni"), 32, 33 (So. Indian Ocean). Kharin, 1984: 116. DIAGNOSIS: Metamorphosed females with filaments on dorsal surface of head posterior to illicial base; filaments on entire posterior margin of illicium; illicial length less than 120% SL; escal bulb with black, spiny distal prolongation; both bulb and prolongation with warty papillae and filaments; lower jaw teeth in 5-6 series posteriorly; caudal-fin rays less than 30% SL. DESCRIPTION: Based on three metamorphosing and one metamorphosed early juvenile female from South Africa. See Bertelsen et al. (1981) for description of larger specimens. Body and caudal peduncle elongate. Head as deep as body, triangular anteriorly in lateral aspect. Cirri on dorsal surface of head in SAM 29239, absent in others (Fig. 17). Scales spinulose, dense but not in contact with one another; only spinule pockets remain over most of body and head in present material. Illicium emerging from snout tip, with few filaments in SAM 29239; filaments absent in others. Illicium longer than head in metamorphosed SAM 27811, shorter than head in others. Escal bulb darkly pigmented except at base, distal prolongation dusky, with few, short filamants except in metamorphosing SAM 27807 and 27808. Few upper jaw teeth in each; tooth buds only in lower jaws of metamorphosing specimens. Measurements in percent SL: head length 22-25; head depth 26-37; postorbital head 5-8; upper jaw 16-18; lower jaw 18-19; snout 11-14; eye diameter 4-5; interorbital width 3; longest upper jaw tooth 0.5-1; longest lower jaw tooth 1.5-2; illicium 17-56; esca length 5-9; esca width 2-3; gill slit 6-11; pectoral base depth 7-8; pectoral-fin 12-13; predorsal 61-73; preanal 75-78; body depth at anal-fin origin 11-15; greatest body depth 22-30; dorsal-fin base 8-9; anal-fin base 8-9; longest dorsal ray 20; longest anal ray 18-20; least caudal peduncle depth 7-10; peduncle length 13-20. Counts: D 6; A 5-6; P 16-17; upper jaw teeth 3-10; lower jaw teeth 0-14; vomerine teeth 0-4. DISTRIBUTION: Circumglobal in southern hemisphere. Meso- and bathypelagic. REMARKS: Two metamorphosing females, SAM 27807 and SAM 27808, were tentatively assigned to G. meadi by Bertelsen et al. (1981), apparently as they represented a significant hiatus in ontogeny between them and the next, more developed, stage, SAM 27811. However, Bertelsen et al. did not report on SAM 29239, intermediate between SAM 27811 and the smaller two. In the development of the head, illicial filaments and escal morphology, we now have no hesitation in identifying all four of the South African specimens as G. meadi. MATERIAL (all females): SAM 29239 (20.5 mm SL); off Kei R. mouth; 32E31.3'S, 29E11.9'E; MWT; 0-1416 m; sta. SM 220; 23 June 1979. SAM 27808 (19.5 mm SL); off Port St. Johns; 31E14.4'S, 30E20.9'E; MWT; 0-1120 m; sta. SM 140; 15 May 1977. SAM 27807 (17.5 mm SL); off Port Shepstone; 30E45.4'S, 30E42.2'E; MWT; 0-830 m; sta. SM 132; 11 May 1977. SAM 27811 (21 mm SL); off Durban; 30E05.5'S, 31E57.0'E; MWT; 0-750 m; sta. SM 157; 18 May 1977. Gigantactis perlatus Gigantactis perlatus Beebe & Crane, 1947: 167, text fig. 13, pl. 2, fig. 3 (Pacific coast of Panama). Bertelsen, 1951: 150. Grey, 1956: 268. Bertelsen et al., 1981: 41, tabs. 1, 2, 10, figs. 1C, 4C, 39, 40. DIAGNOSIS: Illicial length less than 120% SL; lower jaw teeth in 2 irregular series; caudal fin length less than 35% SL; distal prolongation of esca very large (escal length about 5 times its width); esca densely covered with somewhat elongate papillae; escal cirri (short filaments) more numerous distally and in larger specimens; esca with a pair of posterior appendages (PA) at base. DESCRIPTION (from Bertelsen et al., 1981): Females with body and caudal peduncle elongate. Illicial length variable, size-independent, somewhat laterally compressed in larger fish. Esca with a large, conical distal prolongation, increasing in length with increasing standard length. Esca without spines, covered with slightly elongate papillae and scattered, short filaments; (PA) with numerous short branches, becoming more filamentous in larger specimens, rather wing-like in smaller specimens. Fresh coloration of esca pink or whitish in smaller specimens, bright red in largest fish (223 mm SL). Teeth relatively few, in 2 very irregular series in lower jaw. Males unknown. Measurements in percent SL: illicium 74-111 in metamorphosed (over about 30 mm SL) specimens; longest lower jaw tooth 2-6; longest caudal ray 22-31. Counts: D 5-7; A 5-7; P 16-19; upper jaw teeth 2-10; lower jaw teeth 6-18 (no jaw teeth in a 20 mm SL metamorphosing specimen). DISTRIBUTION: Cosmopolitan in tropical and temperate seas. Meso- and bathypelagic. MATERIAL: ISH 1466/71 (female, 152 mm SL); off Vema Seamount; 33E00'S, 07E50'E; MWT; 0-2000 m; WALTHER HERWIG sta. 427/71; 30 Mar. 1971. Not seen. Gigantactis vanhoeffeni Gigantactis vanhoeffeni Brauer, 1902: 296 (off Kenya). Brauer, 1906: 322, pl. 15, figs. 8-9. Regan, 1926: 38. Regan & Trewavas, 1932: 93. Beebe & Crane, 1947: 168. Bertelsen, 1951: 150. Grey, 1956: 267. Pietsch & Seigel, 1980: 396. Bertelsen et al., 1981: 31, figs. 1B, 4A, 5, 6, 8, 12, 19, 21, 30, 31. Masuda et al., 1984: 106, pl. 93A. Kharin, 1984: 116. Bertelsen, 1986: 1406, text figs. Bertelsen, 1991: 514. Nielsen & Bertelsen, 1992: 62, text fig. Gigantactis exodon Regan & Trewavas, 1932: 94, pl. 5, fig. 1 (Caribbean Sea). Bertelsen, 1951: 150-151. Grey, 1956: 268. DIAGNOSIS: Metamorphosed females with illicial length less than 120% SL; illicium with pair of appendages just below escal bulb; escal bulb with dark distal prolongation, both bulb and prolongation with flattened papillae; tuft of short filaments at tip of prolongation in larger specimens; lower jaw teeth in 3 series; caudal-fin rays less than 45% SL. DESCRIPTION: Based on Bertelsen et al., 1981; counts and measurements from our single specimen which now lacks the esca and has a damaged illicium, with its distal end skinned. Females with body and caudal peduncle elongate. Illicium spiny, laterally compressed, more distinctly so in larger specimens, with a pair of small appendages just below esca, becoming bifurcated cirri in some large specimens; short, slender illicial filaments present distally in specimens over 70 mm SL. Esca with spiny distal prolongation, more thread-like in small specimens, greatly thickening and developing tuft of short, thick filaments at tip in larger fish. Few, short filaments on escal bulb near light organ. Escal papillae tack-like, some black, others almost transparent. Paired illicial appendages (IA) and tips of distal illicial filaments bright red in freshly caught specimens. Number of jaw teeth increasing with size, lower jaw with 3 distinct rows of teeth posteriorly, rows rather obscure anteriorly. Males unknown. Measurements in percent SL: head length 31; head depth 22; postorbital length 15; upper jaw 15; lower jaw 21; snout 12; eye diameter 2; interorbital width 4; longest upper jaw tooth 1; longest lower jaw tooth 4; longest upper pharyngeal tooth 4; illicium broken into 4 parts; gill slit 4; pectoral base width 4; pectoral-fin 6; predorsal 58; preanal 62; body depth at anal-fin origin 16; greatest body depth 23; dorsal-fin base 12; anal-fin base 11; longest dorsal ray 17; longest anal ray 13; longest caudal ray 32; least caudal peduncle depth 8; peduncle length 24. Counts: D 6; A 6; P 17; vomerine teeth 0; upper jaw teeth 42; lower jaw teeth 92. DISTRIBUTION: Adults meso- and bathypelagic in temperate and tropical seas. One record off southern Greenland. MATERIAL (females): SAM 35017 (133 mm SL); Agulhas Bank; 36E53'S, 21E33'E; BT; 0-1000 m; A12167-096; 25 Oct. 1991. UW 22178 (132 mm SL); same data as preceding. FAMILY LINOPHRYNIDAE DIAGNOSIS: Females with globose bodies; flesh scaleless and gelatinous; sphenotic and preopercular spines strong; illicium short, its pterygiophore ("basal bone") concealed or just protruding on snout; pectoral radials 3; hyoid barbel present in Linophryne; D 2-4; A 2-4; P 13-19; C 9; branchiostegal rays 4-5. Males obligatory parasites; metamorphosed free-living specimens with denticular teeth short and stout; illicial pterygiophore and upper denticular bone separated; olfactory organs large, anterior nostrils directed laterally; eyes tubular, directed forward somewhat; skin naked; premaxilla and jaw teeth present in 2 genera, reduced or absent in other genera. The linophrynids comprise 5 genera and 47 species. Adults are meso- and bathypelagic, occurring from just above 200 m to 1900 m. Only the largest genus, Linophryne, is represented in our area by 3 species. Recorded prey of netdevils include fishes and crustaceans. Females of Linophryne reach 23 cm SL (L. lucifer; Bertelsen and Krefft, 1965). Males are obligatory sexual parasites (Pietsch, 1976), and Munk and Bertelsen (1983) described the anatomy of the vascular network (plexus) between attached males and female hosts in the netdevil Haplophryne mollis. The functional anatomy of escal light guides and luminescence control in H. mollis and Linophryne coronata were described by Herring and Munk (1994) and Munk (1998). The Danish worker Erik Bertelsen published a series of 8 papers on Linophryne, the last of which (Bertelsen, 1982) contains a key to the 21 species then recognized for the genus. Gon (1992) subsequently named L. andersoni from the central Pacific. Linophryne Collett, 1886 DIAGNOSIS: Females with short, globose or tapering bodies. Head massive, teeth fang-like. Branchiostegal rays 4-5. Sphenotic spines well developed. Skin black, naked. Pelvic bones absent. Pterygiophore of illicium short, concealed under skin or just protruding. Luminous hyoid barbel present. Vomerine teeth present. D 2-4 and A 2-4 (both usually 3); P 13-19; C 9. See Bertelsen (1951, 1982) for diagnoses of males and larvae. KEY TO SPECIES (Females) 1a. Hyoid barbel with 3-5 main branches near base; no line of filaments at tip of esca 2 1b. Barbel elongate, unbranched at base, bifurcated at tip, longer than SL; 4 (or 5?) short, unpigmented filaments at tip of esca L. digitopogon 2a. Distal escal appendage (DA) long and slender, with 6-10 pairs of filaments (in adults); posterior escal appendage (PA) small, papilliform or digitiform; barbel with 5 main branches near base L. densiramus 2b. Distal escal prolongation short, papilliform, without filaments; paired posterolateral escal appendages (PLA) long, digitiform, with highly filamentous tips; barbel with 3 main branches near base L. parini Linophryne densiramus Linophryne densiramus Imai, 1941: 247, figs. 14-17 (Suruga Bay, Japan). Bertelsen, 1980a: 55, figs. 3C, 4D, 5D, 11-12. Bertelsen & Pietsch, 1983: 94, fig. 16. Paxton et al., 1989: 296. Linophryne arborifera (non Regan, 1925): Bertelsen, 1951: 178-183 (in part). Masuda et al., 1984: 108, pl. 93H. DIAGNOSIS: Distal escal appendage elongate, filamentous in adults; posterior escal appendage reduced; barbel multi-stemmed, with 5 main branches near base; illicial length 14.5-24% SL. DESCRIPTION: Females with massive, roughly triangular head. Body tapering posteriorly; body black, fins unpigmented. Distal escal appendage unpigmented; escae of our specimens have only slightly fewer pairs of filaments than illustrated by Bertelsen (1980a, fig. 11). Jaw teeth in 2 series, inner fang-like; vomer with one tooth on each side. Measurements in percent SL: head length 44-56; head width 22-26; head depth 42-51; postorbital length 20-28; upper jaw 37-42; lower jaw 45-53; snout 17-26; eye diameter 5-8; interorbital width 10-14; longest upper jaw tooth 7-9; longest lower jaw tooth 8-14; illicium 20-23; esca length 10-12; esca width 7; distal escal appendage 17-18; barbel 57-77; gill slit 7-9; pectoral base width 8-10; pectoral-fin 14-16; predorsal 74-80; preanal 78-84; body depth at anal-fin origin 23-33; greatest body depth 47-51; dorsal-fin base 5-6; anal-fin base 4-6; longest dorsal ray 17-25; longest anal ray 21-27; least caudal peduncle depth 12-15; peduncle length 2-4. Counts: D 3; A 3; P 16; upper jaw teeth 24-40; lower jaw teeth 19-31; vomerine teeth 2; branchiostegal rays 5. A 17 mm SL male is attached to the 71 mm SL female of RUSI 54760 just anterior to the anus by separate outgrowths from the male’s snout and lower jaw; no denticular teeth remain. Nasal lamellae 10. DISTRIBUTION: Cosmopolitan and mesopelagic in tropical and temperate seas (Bertelsen, 1980a, fig. 17; Chilean record not shown). MATERIAL (females): SAM 31705 (41 mm SL); off Childs Bank; 31E14.6'S, 15E35.5'E; BT; 0-796 m; sta. A9827-079; 26 Jan. 1990. RUSI 54760 (71 mm SL); off Cape Columbine; 33E36'S, 17E07'E; BT; 0-977 m; sta. A18562-139; 24 Jan. 1997. SAM 31593 (45 mm SL); Cape Point Valley; 34E46.2'S, 18E02.9'E; BT; 0-710 m; sta. A7036-060; 14 Mar. 1988. Linophryne digitopogon Linophryne digitopogon Balushkin & Trunov, 1988: 62, fig. 1 (off Childs Bank, South Africa, 0-920 m). Heemstra, 1991: 377, text fig. DIAGNOSIS: No escal appendages, instead, line of 4 (or 5?) short filaments on low papilla at distal end; illicial length 28% SL; hyoid barbel unbranched at base, bifurcated at tip. DESCRIPTION (from Balushkin & Trunov, 1988): Single 48 mm TL female. Head massive; jaws relatively longer (60-70% SL) than other two species in our area. Sphenotic spines well developed. Teeth fang-like. Greatest body depth at about level of pectoral lobe, rapidly tapering posteriorly. Body black; no pigment on distal ends of esca and barbel, skin under eye and base of caudal fin. Tip of esca with low papilla bearing 4 (?) filaments arranged in a line (but 5 shown in original description). Hyoid barbel longer than standard length, thread-like, with two branchings, proximalmost short, ca. 5% SL, distalmost deeper, bearing group of photophores. Measurements in percent SL (from Balushkin and Trunov, 1988): head width 43; upper jaw 60; lower jaw 73; snout 25; eye diameter 6; interorbital width 12; longest upper jaw tooth 19; longest lower jaw tooth 27; illicium 28; esca 14; barbel 125; predorsal 87; preanal 93. Counts: D 3; A 3; P 14; upper jaw teeth 22; lower jaw teeth 16; vomerine teeth 2; branchiostegal rays 5; vertebrae 20. DISTRIBUTION: Known only from the holotype, ZIN 48199 (female, 47.5 mm TL); WNW of Childs Bank; 30E38'S, 14E50'E; 0-920 m, M/V GIZHIGA; 17 Feb. 1974. Not seen. Linophryne parini Linophryne parini Bertelsen, 1980b: 234, fig. 1 (off Fish R. mouth, South Africa, 0-1220 m). Bertelsen, 1982: 87, 91, 96. Heemstra, 1991: 377, text fig. DIAGNOSIS: Distal end of esca with a short bulbous swelling, filaments absent; a pair of posterolateral escal appendages (PLA), their tips with numerous filaments; illicial length 15% SL; hyoid barbel with 3 main branches at base. DESCRIPTION (from Bertelsen, 1980b; single 64 mm SL female): Body and fins as for congeners. Fins unpigmented except for small spot near tip of each pectoral ray. Esca globular, pigmented along base and laterally along light organ; no true distal escal appendage (DA), instead, a low, rounded, unpigmented swelling present. Pair of posterolateral escal appendages (PLA), with tapering stalks and filamentous tips. Barbel pigmented at base only, of 3 main branches and a small, unbranching filament proximally; each main branch ramifying into numerous secondary and tertiary filaments, with single photophore distally. Measurements in percent SL: head length 55; head width 20; upper jaw 48; lower jaw 50; snout 25; eye diameter 5; longest upper jaw tooth 8.5; longest lower jaw tooth 12.5; illicium 15; barbel 44. Counts: D 3; A 3; P 14-15; upper jaw teeth 45; lower jaw teeth 32; vomerine teeth 3; branchiostegal rays 5; vertebrae 21. DISTRIBUTION: Known only from the holotype, IOAN uncat. (female, 64 mm SL); off Fish R. mouth, South Africa; 33E41.6'S, 27E26.0'E; BT; 0-1220 m; FIOLENT cr. 3, sta. 126. Not seen. ACKNOWLEDGEMENTS We thank the staff of the South African Museum and Sea Fisheries for help in the collection and curation of the material studied here, especially P. A. Hulley, L. J. V. Compagno and M. G. van der Merwe. Other data and illustrative materials were facilitated with the assistance of J. G. Nielsen, Zoologisk Museum, Copenhagen, G. Schultze, Zoologisches Museum, Hamburg, and T. W. Pietsch, University of Washington. Mrs. E. Heemstra rendered the fine original illustrations here. REFERENCES
The following images related to this document are available:Photo images[fb01002e.jpg] [fb01002h.jpg]Line drawing images[fb01002k.gif] [fb01002i.gif] [fb01002c.gif] [fb01002f.gif] [fb01002a.gif] [fb01002n.gif] [fb01002l.gif] [fb01002b.gif] [fb01002j.gif] [fb01002m.gif] [fb01002g.gif] [fb01002o.gif] [fb01002d.gif] |
| |||||||||

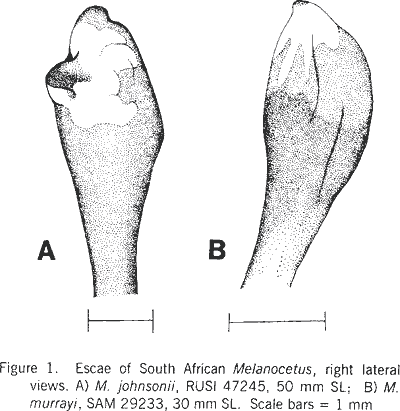{kind=link}
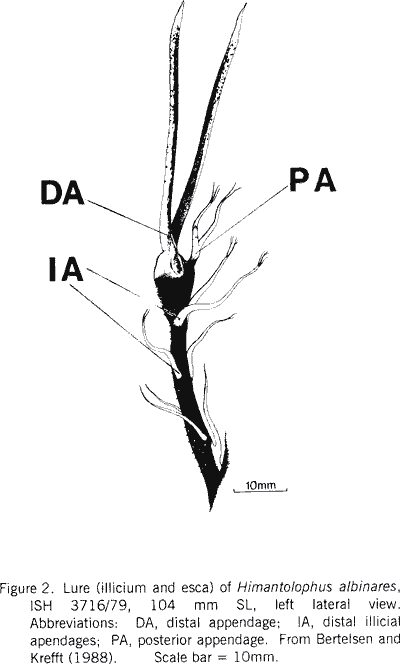{kind=link}
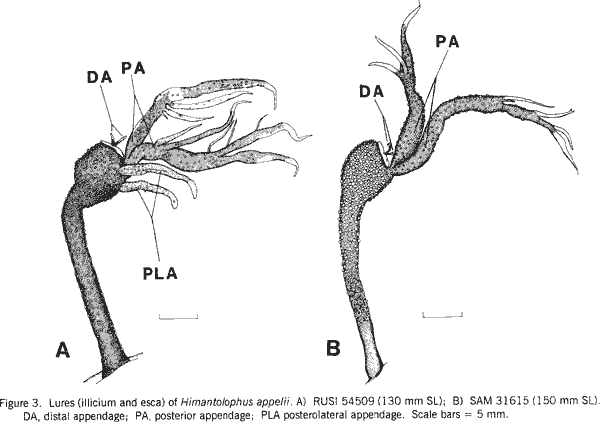{kind=link}
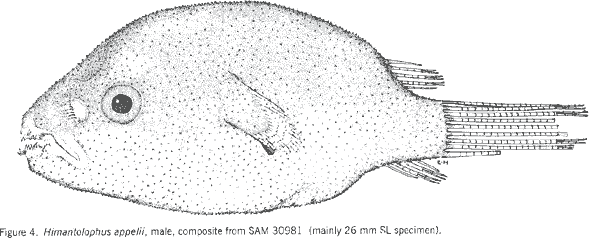{kind=link}
{kind=link}
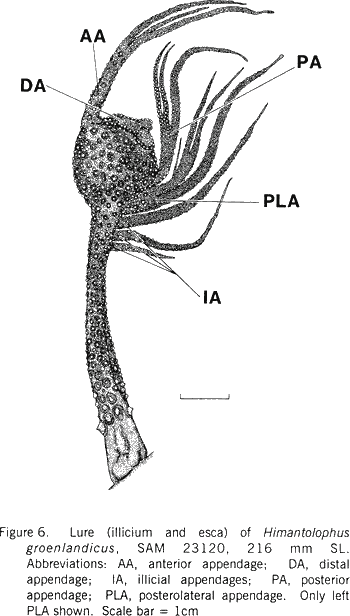{kind=link}
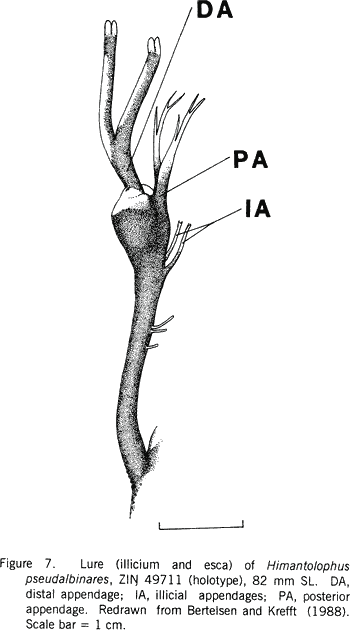{kind=link}
{kind=link}
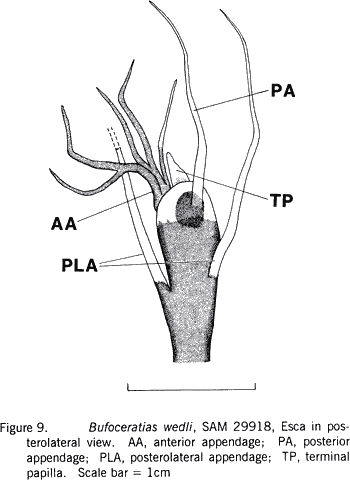{kind=link}
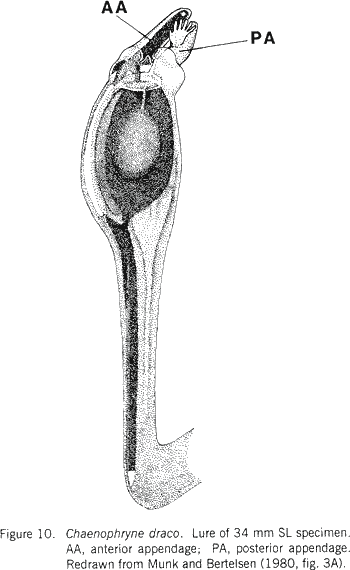{kind=link}
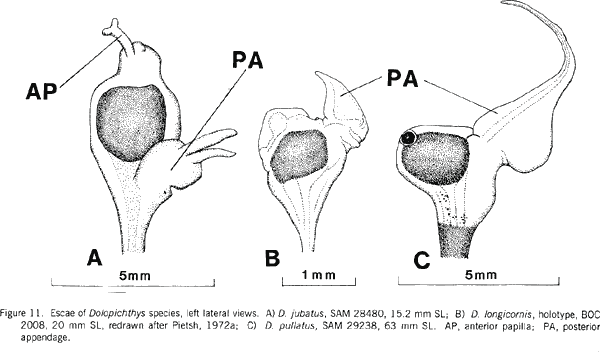{kind=link}
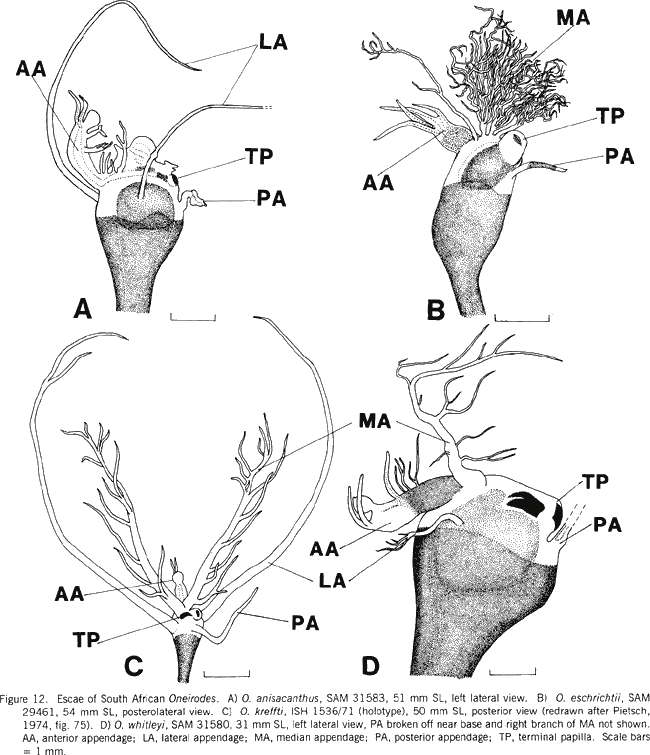{kind=link}
{kind=link}
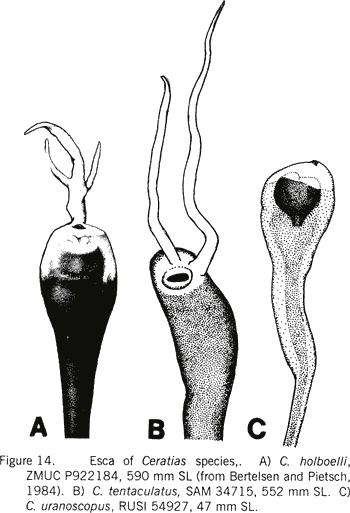{kind=link}
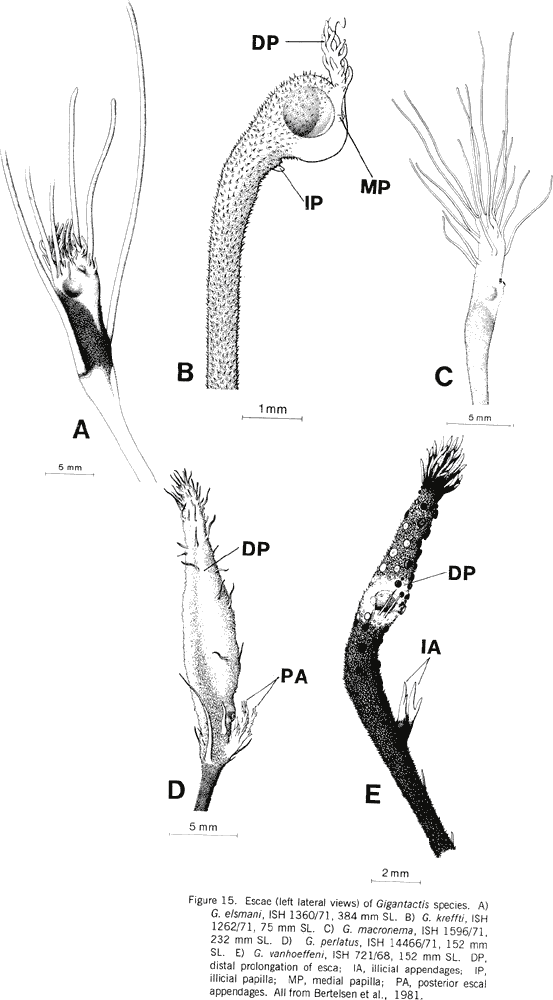{kind=link}