 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ichthyological Bulletin No. 65, September 1995
A CONTRIBUTION TO THE TAXONOMY OF THE MARINE FISH GENUS ARGYROSOMUS
(PERCIFORMES: SCIAENIDAE), WITH DESCRIPTIONS OF TWO NEW SPECIES FROM
SOUTHERN AFRICA.
^1 Sea Fisheries Research Institute, Private Bag X2, Rogge Bay 8012, Cape
Town.
Code Number: FB95004
Size of Files:
Text: 146K
Graphics: line drawings (gif) - 220K
[ALL TABLES AT END OF PAPER]
ABSTRACT Study of the biology, anatomy and taxonomy of the sciaenid fishes of the genus Argyrosomus from South Africa and Namibia revealed that three species were confused under the name "Argyrosomus hololepidotus (Lacepede, 1801)". Comparison of morphometric and meristic data, otoliths, swim-bladders, drumming muscles, and other morphological features, of specimens from southern Africa, Madagascar, the Mediterranean Sea, the eastern Atlantic Ocean, Japan and Australia, established that the "A. hololepidotus" of recent authors is a complex of four species: A. japonicus (Temminck & Schlegel, 1843), which occurs off southern Africa, Japan and Australia; A. inodorus sp. nov., which is known from Namibia to the Kei River (32 degrees 40' S) on the east coast of South Africa; A. coronus sp. nov., which is known from central and northern Namibia and Angola, and A. hololepidotus, which appears to be endemic to Madagascar. These four species are compared with A. regius (Asso, 1801) of the Mediterranean and eastern Atlantic, A. thorpei Smith, 1977 from South Africa, Mozambique and the west coast of Madagascar, and A. beccus Sasaki, 1994 known only from Durban harbour. To promote stability in the nomenclature and to resolve the confusion in the taxonomy of Argyrosomus species, neotypes are selected for A. hololepidotus and A. japonicus. The biology, distributions and fisheries of six species are reviewed. Distribution patterns for the southern African species and a key to the seven species known from Africa and Madagascar (A. regius, A. japonicus, A. inodorus, A. coronus, A. thorpei, A. beccus, and A. hololepidotus) are provided. The composition and distinction of the genus Argyrosomus are briefly discussed. INTRODUCTION Sciaenid fishes of the genus Argyrosomus occur in the eastern Atlantic and Indo-West Pacific regions, and are important food species wherever they are found. Close similarity in the external appearance of these species has resulted in a difficult and sometimes confused taxonomy. Correct identification of Argyrosomus species is essential from a management perspective, as different species often exhibit different life history traits, and therefore require separate conservation strategies.
Since 1977 two species of Argyrosomus, "A. hololepidotus" and A. thorpei, were recognised as important recreational and commercial fishes off South Africa. In 1990, a project initiated by the first author (MHG) to study the biology of "Argyrosomus hololepidotus", revealed that two species were confused under this name. In order to establish their identity, these two species were compared with other species of Argyrosomus. The results are presented in this paper, together with descriptions of A. hololepidotus, A. regius, A. japonicus and A. thorpei. Neotypes are designated for A. hololepidotus and A. japonicus. Two new species, A. inodorus and A. coronus are also described from southern Africa, bringing the total number of Argyrosomus species to ten.
Taxonomic differences between A. inodorus and A. japonicus were studied in detail, as both species are important in recreational and commercial fisheries in South Africa. The habitats and distributions of the Southern African Argyrosomus species are discussed, and the biology and fisheries of the six African species are reviewed. MATERIALS AND METHODS The specimens examined are listed after each description. Institutional abbreviations follow Leviton et al. (1985). Sex is indicated by the letters M (male) and F (female); juveniles, too small to be sexed, are indicated by "J". In cases where additional fresh specimens were examined but not preserved, the total sample number (n) includes these fresh specimens. Except as indicated below, measurements and counts used in this study were based on the methods of Hubbs and Lagler (1964). Counts of gill-rakers and swim-bladder appendages exclude rudiments (structures wider than long). Swim-bladder appendages were counted on the left side of the swim-bladder. Orientation of swim-bladder appendages may vary depending on the amount of fat in which they are embedded. Our descriptions of appendage orientation for the species treated here are based (when possible) on specimens with little or no fat.
Vertebral counts were made on skeletons (see below) or on radiographs. Caudal vertebrae were taken as those with fused transverse processes or distinct haemal spines. Body depth was measured between the origins of the first dorsal and the pelvic fins. Caudal peduncle length was measured obliquely from the end of the second dorsal fin base to the lateral line at the base of the caudal fin. Pre-pelvic length was taken from the tip of the lower jaw to the origin of the pelvic fin. The least fleshy interorbital width was measured. As specimen treatment causing shrinkage of the eye (e.g. storage in alcohol or freezing), generally also caused shrinkage in this measurement, data for fresh specimens is provided in the key where relevant. Pectoral fin length was measured from the origin of the most dorsal ray to the tip of the longest ray. Anal and pelvic fin lengths were taken from the fin origin to the distal tip of the fin. The first lower-limb gill-raker (next to the raker at the angle), and the longest gill filament of the first gill arch were measured. The heights and lengths of scales on the body at the tip of the adpressed left pectoral fin were measured. The length of the longest urinary bladder (left or right, Fig. 1) of A. japonicus and A. inodorus was measured to the nearest millimetre. The size of the bladder was influenced by the degree of fullness at the time of death. In order to compensate for bladder fullness, measurements were taken from fresh material after the bladders were emptied and straightened. The shape of the caudal fin varies with species, specimen size and expansion. To compensate for variable expansion, the fin was fully splayed (while still wet) on a smooth surface, and then allowed to contract to the "natural" position. Variation in caudal fin shapes and terminology are illustrated in Figure 2. Notes on life colour were taken from fresh specimens. The examination of more than five hundred live A. inodorus and A. japonicus suggest that both species darken after death.
Figure 2. Argyrosomus caudal fin shapes and terminology. A) elongate pointed; B) elongate S-shaped; C) S-shaped; D) bi-concave; E) wedge-shaped; F) rounded; G) truncate.
Sonific (drumming) muscles run in a narrow longitudinal band (one on each side) along the inside of the ventrolateral wall of the posterior part of the body cavity. The muscle fibres run transversely to the long axis of the muscle and are attached to the medial surface of the peritoneum. They are not connected to the swim-bladder.
Morphometric ratios are expressed as percentages of head length (HL) (measurements on head only) and standard length (SL). Unless indicated as "TL", all fish lengths are of SL. For statistical analyses, measurements were expressed as fractions of SL and then arcsine-transformed. Meristic data and morphometric ratios (except A. hololepidotus with n = 2 or 3) were compared using ANOVAs and Tukey studentized range tests. Morphometric ratios for A. inodorus and A. japonicus were also analyzed according to 100 mm SL size classes to compare characters that show allometric growth (e.g. eye diameter) or those which grow at different rates in the two species (e.g. pectoral fin length). In addition to comparing the general morphology of the sagittae of five Argyrosomus species, the relationships between sagittal measurements (and weight) and TL for A. inodorus and A. japonicus were compared using regression analysis. Total length (TL) was used instead of SL, because it could be measured more rapidly when recording data from commercial catches. Otoliths were taken from all sizes (A. inodorus, 60 - 1378 mm TL; A. japonicus, 36 - 1750 mm TL) represented in the catch. Otolith weight (to the nearest milligramme) and measurements (to the nearest 0.01 mm, see Fig. 3) were taken from the left sagitta. Fish lengths and otolith measurements (including weight) were log-transformed and straight line regressions were calculated. The corresponding regression lines for A. inodorus and A. japonicus were tested for differences in slope and intercept (Myers, 1990).
Height, OCD = Ostium to Cauda Distance, PM = Posterior Margin, VM = Ventral Margin.
Twelve skeletons of A. inodorus, 7 of A. japonicus, 2 of A. thorpei and 1 of A. coronus were compared. Measurements of cranial features (Figs 4 & 5) were taken as follows: pre-supraoccipital crest length (PSOCCRL) was measured point-to-point along the median axis from dorsoanterior tip of mesethmoid to dorsal end of frontal/supraoccipital suture; supraoccipital crest length (SOCCRL) was measured similarly to PSOCCRL from frontal/supraoccipital suture to the posterior edge of supraoccipital crest, immediately below the dorso-posterior angle; neurocranial length (NCL) was taken from the anterior tip of the vomer to the posterior tip of the basioccipital; neurocranial width (NCW) is the distance between the anterior ends of the sphenotic bones. The left dentary was measured from the anterior tip to the posterior tip of the ventral processes (Fig. 6). The length of the left premaxilla was measured as the maximum straight line distance between anterior and posterior tips.
IOW = Interorbital Window; LLC = Lateral Line Canal; NCL = Neurocranial Length; PSOCCRL = Pre-Supraoccipital Crest Length; SOCCRL = Supraoccipital Crest Length. Scale bar = 10 mm.
Figure 5. Dorsal
view of neurocrania: A)
Argyrosomus inodorus, 365 mm; B) A. japonicus, 365 mm; C)
A. coronus, 495 mm; D) A. thorpei, 461 mm. Neurocranial width
is the distance between the anterior ends of the sphenotic bones (arrows);
Figure 6. Left
dentaries (lateral view): A)
Argyrosomus japonicus, 525 mm; B) A. thorpei, 461 mm; C)
A. inodorus, 433 mm; D) A. coronus, 495 mm.
Argyrosomus Pylaie 1835 Argyrosomus Pylaie, 1835: 532. Type species Argyrosomus procerus Pylaie, 1835 (substitute name for "Sciaena aquila Cuvier" = Cheilodipterus aquila Lacepede, 1803, = Perca regia Asso, 1801 = Argyrosomus regius) by monotypy. See Trewavas (1977) for details of synonymy. DESCRIPTION: Body fairly elongate and moderately compressed; dorsal profile more convex than ventral profile. Mouth terminal, slightly oblique; jaws equal anteriorly, or lower jaw protrudes slightly. Chin without barbel but with three pairs of mental pores: the median pair rounded, the second oval or elongate, and third pair narrow slits, each pair progressively further apart. No large canines; teeth differentiated in size; upper jaw with outer row of enlarged teeth and narrow inner band of smaller teeth; lower jaw with single inner row of enlarged teeth and outer row of smaller teeth; smaller teeth in both jaws less visible in larger specimens. Anterior nostril round to oval, the posterior one slightly larger and oval to slit-like. Preopercle serrated; operculum ending in two weak, flattened spines (often hidden by skin). Pectoral fins relatively short (15-23% SL), with a dark fleshy axillary fold at upper end of fin base. Caudal fin pointed (in small juveniles) to S-shaped, rhomboid, rounded or even truncate (in young and adults). Lateral-line scales 47-53 (usually 51) to base of caudal fin; lateral-line tubules arborescent; lateral-line scales extend to end of caudal fin.
Sagitta moderately elongate, with prominent postcentral umbo on outer surface; inner (medial) surface convex, with enlarged, spoon-shaped ostium and strongly curved, J-shaped cauda with bluntly rounded tip; otoliths of adults generally deepest within posterior third, close to tip of cauda. The sagitta of the type species (A. regius) is indistinguishable from that of A. hololepidotus or A. japonicus (Figs 11, 14 & 15). Since A. regius is the type-species of the genus, this general otolith morphology is referred to as the Argyrosomus form.
Swim-bladder typically carrot-shaped (tapering posteriorly) with 21-45 short appendages that may or may not (individual variation) be embedded in a wedge of fatty tissue; no appendages enter the head (i.e. pierce the transverse septum); most appendages arborescent, divided initially into two distinct limbs which are often orientated dorso-posteriorly and ventro-anteriorly, respectively; last 2-6 appendages reduced in size and bud-like or weakly branched (last appendage not tubular). Drumming muscles of Argyrosomus species may be present in both sexes or in males only. Generally 11 abdominal and 14 caudal vertebrae. REMARKS: In her excellent revision of the Indo-West Pacific sciaenids, Trewavas (1977) recognized five species of Argyrosomus: A. regius, hololepidotus, japonicus, miiuy and amoyensis. Mohan (1984) added Sciaena heinii Steinachner, 1902 to the genus (Trewavas [1977: 451] treated S. heinii as a "doubtful species", because she had not examined any specimens and was unsure of its genetic assignment. Mohan's (1984) description of the swim bladder, otolith, dentition and external morphology of this species agrees with those of Argyrosomus (as described above).
A comprehensive revision of the genus Argyrosomus was beyond the scope of our present work, but some comments (we hope constructive) on the currently accepted limits and definition of this genus are in order. Sasaki (1989) recognized the genus Miichthys Lin, 1983 for Sciaena miiuy Basilewsky, 1855 and put this species in a new monotypic tribe, Miichthyini, "distinguished by the combination of well curved sulcus tail [cauda of the sagitta], absence of enlarged swim-bladder appendages, absence of cephalic swim-bladder appendages, scaly dorsal fin". As discussed by Sasaki (1989: 81, 90-92), the otolith and swim-bladder features that distinguish Sciaena miiuy from A. japonicus and other members of his tribe Argyrosomini are relatively primitive (plesiomorphic) character states. The presence of enlarged anterior swim-bladder appendages is a dubious synapomorphy for the tribe Argyrosomini, because it appears only in larger specimens. Enlarged appendages are known in A. regius, A. japonicus, and A. inodorus, but they were not present in 46 cm SL specimens of A. thorpei. They are not known for A. hololepidotus, A. coronus or A. amoyensis, but this may be because the swim-bladders of larger individuals of these three species have not been examined. The two autapomorphies: ("frontal projecting downward" and "caudal fin being truncate to lunate in adults") cited by Sasaki (1989:118) for A. japonicus are also dubious as defining character states for the genus Argyrosomus. The "frontal projecting downward" refers to the increased ossification of the interorbital septum in A. japonicus (Fig. 4B), in which the frontals are fused ventrally into a median lamina that extends ventrally and fuses with similar lamina arising from the median ethmoid and parasphenoid bones. In A. coronus (Fig. 4C) the frontal and parasphenoid bones are also fused, but in A. inodorus (Fig. 4A) and A. thorpei (Fig. 4D), the interorbital septum is not so well ossified, and the median frontal lamina does not contact the parasphenoid bone. The state of the interorbital septum in A. hololepidotus, A. amoyensis, and the type-species (A. regius) are unknown. The value of this character state (increased ossification of interorbital septum) as an indication of a monophyletic genus is further diminished by its homoplastic development in more distantly related genera (e.g., Johnius and Kathala, Sasaki, 1989).
Judging from several thousand South African specimens and eight specimens from Japan (including the photograph of Masuda et al., 1988: P1. 147, Fig. E), it is not quite accurate to describe the caudal fin of A. japonicus as "truncate to lunate in adults". The caudal fin of adults is most commonly S-shaped (Fig. 2C, with the rear margin of the upper half of the fin concave and the lower half convex), less frequently hi-concave (Fig. 2D) and seldom wedge shaped (Fig. 2E) or truncate (Fig. 2G). In view of the ontogenetic variation in caudal fin shape, and the fact that other (distantly related) sciaenids also have s-shaped or truncate caudal fins, the value of the caudal fin shape as a defining character state for the genus Argyrosomus is also questionable.
Although Sasaki's (1989) evidence for recognition of Miichthys and the tribes Miichthyini and Argyrosomini is not convincing, we do not have sufficient information to contest or support these taxa.
Sasaki (1994) described Argyrosomus beccus as a new species from South Africa based on three specimens (175, 191 & 230 mm SL) collected from Durban Harbour in 1969. No further specimens of this anomalous species have been collected in South African waters. Anatomically, it differs strikingly from other species of Argyrosomus in having an unusually short beak-like snout, with a slight concavity in the dorsal head profile just before the eye (Fig. 8). The snout length (6.1-6.3% SL, 20-21% HL) is shorter than the eye diameter (7.3-8.3% SL, 24-27% HL); whereas, in the six species that we measured, the snout lengths (7.4-9.6% SL, 24-31% HL) are always longer than the eye diameters (4.7-7.9% SL, 14-24% HL). The interorbital width of A. beccus is also much narrower (4.7% SL, 15.2-15.6% HL) than in the other five species (6.0-8.5% SL, 19-29% HL).
---------------------------------------------------------------------------
1a. Snout length 20-21% HL, shorter than eye diameter (24-27% HL) . . .
. . . . . . A. beccus
1b. Snout 24-31% HL and longer than eye diameter (14-24% HL) . . . . 2
2a. Axillary skin fold above pectoral fin base scaly; anterior part of
lateral line strongly curved; peritoneum pigmented with fine black
dots; fins orange brown or dusky yellow in life; otolith (sagitta)
with tip of cauda expanded and truncate . . . . . . A. thorpei
(Algoa Bay to southern Mozambique)
2b. Axillary skin fold of pectoral fin without scales; anterior part of
lateral line slightly curved; peritoneum unpigmented; otolith cauda
J-shaped, the tip rounded and not expanded . . . . . . . . 3
3a. Caudal peduncle depth 58-74% peduncle length; ostium to cauda distance
37-63% otolith height; drumming muscles absent in females; length of
body scales at tip of pectoral fin 1.8-2.5% SL; interorbital width
20-25% HL (23-26% in fresh specimens); swim-bladder appendages 31-42
. . . . . . . . . . . . . . . . A. inodorus sp. nov.
(Namibia to Kei River)
3b. Caudal peduncle depth 68-94% peduncle length; ostium to cauda distance
23-34% otolith height; drumming muscles present in both sexes of
adults (> 27 cm SL) . . . . . . . . . . . . . . . . 4
4a. Interorbital width 24-29% HL (27-30% in fresh specimens) . . . . 5
4b. Interorbital width 20-24% HL (22-25% in fresh specimens) . . . . 6
5a. Swim-bladder appendages 36-45; caudal peduncle depth 68-82% peduncle
length; length of midlateral body scales. 1.8-2.3% SL, lower jaw
44-48% HL . . . . . . . . . . . . . . . . A. regius
(Mediterranean, eastern Atlantic, and Red Sea)
5b. Swim-bladder appendages 28-38; caudal peduncle depth 76-94% peduncle
length; length of midlateral body scales 2.2-2.7% SL, lower jaw
46-50% HL . . . . . . . . . . . . . A. coronus sp. nov.
(Namibia and Angola)
6a. Suborbital width 10.5-12% SL; swim-bladder appendages 34-36 . . . .
. . . . . . . A. hololepidotus
(Madagascar)
6b. Suborbital width 8.4-10.5% ½L; swim-bladder appendages 21-31 . . .
. . . . . . . . . A. japonicus
(Indo-West Pacific to False Bay, South Africa)
---------------------------------------------------------------------------
Argyrosomus regius (Asso, 1801)
Perca regia Asso, 1801:42 (coasts of Spain). Cheilodipterus aquila Lacepede, 1803: 685, Pl.21, Fig.3 (French Atlantic).
Sciaena umbra (non Linnaeus): Cuvier, 1814: 13, Pl. Sciaena aquila: Cuvier, 1816: 298; Risso, 1826:411 (Mediterranean); Cuvier, in Cuvier & Valenciennes, 1830: 28, Pl. 139; Day, 1880-1884: 150, Pl. L (Great Britain); Smitt, 1892: 50, Fig.13. Pseudosciaena aquila: Bleeker, 1863: 142; Bleeker, 1876: 329. Perca vanloo Risso, 1810: 298, Pl. 9, Fig. 30 (Western Meditteranean). Argyrosomus procerus Pylaie, 1835:532 (substitute name for Sciaena aquila Cuvier). Johnius hololepidotus (non Lacepede): Fowler, 1936:884 (described from Mediterranean specimens). Argyrosomus regius: Trewavas, 1966: 4; 1973: 397; 1977: 324, Fig. 8; Seret, 1981: 266. For additional references see Trewavas, 1973. Neotype: MNHN 7511, La Rochelle, French Atlantic; designated by Trewavas (1966) quoting a description by P.J.P. Whitehead (MS). DIAGNOSIS: A species of Argyrosomus with the following combination of characters: dorsal fin rays 26-30; gill-rakers 3-5 + 8-10; swim-bladder carrot shaped, with 36-45 arborescent appendages; caudal peduncle length 11-13% SL; peduncle depth 68-82% peduncle length; interorbital width 7.3-8.5% SL (24-29% HL); suborbital width 8.4-11% HL; lower jaw length 45-48% HL; midlateral body scale length 1.8-2.3% SL; drumming muscles present in both sexes; urinary bladders large; sagitta typically Argyrosomus (Fig. 10), OCD 26-32% OH; pectoral fins 18-21% SL, not reaching past vertical at tip of pelvic fins; pectoral fin axillary fold scaleless; peritoneum unpigmented; anterior part of lateral line slightly curved. DESCRIPTION: Based on 19 whole specimens (173-433 mm) plus otoliths, swim-bladders, interorbital widths, head lengths and SL of 7 others collected for us by Bernard Seret. Due to the poor condition of five specimens (MNHN 7511, MNHN 7533, MNHN 7945, MNHN 1524, and the neotype), only the interorbital width and SL measurements of these fish were included in the following description. Trewavas' (1977) data (10 specimens 135-965 mm SL) are given in parentheses. Dorsal fin X+I,26-30 (IX-X+I,26-29); anal fin II,7; pectoral-fin rays 16-18; lateral-line scales 49-53(47-51); gill-rakers 3-5 + 8-10 (2-5 + 9); swim-bladder appendages 36-45 (40-42); vertebrae 11 + 14.
Proportions as % SL: head length 28-34 (27-35); predorsal length 33-37; pre-pelvic length 29-39, usually 30-34; body depth 24-34 (25-36); caudal peduncle length 11-13; peduncle depth 7.9-10.8; snout 8.1-9.6; eye diameter 5.5-6.5 at 173-222 mm, 4.1-5.1 at 261-339 mm and 4.3 at 433 mm SL; interorbital width 7.3-8.5; suborbital width 2.7-3.5; upper jaw 12-14; lower jaw 14-16; pectoral fin 18-21 (1823); pelvic fin 19-21 at 173-198 mm, 17-19 at 261-342 mm and 17 at SL 433; anal fin 14-18; third dorsal spine 11-16; second anal spine 7.0-8.9 (6-9); gill-raker 1.6-2.4; gill-filament 2.8-3.3. Proportions as % HL: snout 26-29 (26-31); eye diameter 17-19 at 173-222 mm, 12-16 at SL 261-342 mm and 14 at 433 mm SL (15-19 at SL 193-494 mm and 10-11 at SL 950-965 mm); interorbital width 24-29 (21-26); suborbital width 8.5-11; upper jaw 38-42 (39-44); lower jaw 45-48 (42-49); gill-raker 5.0-7.2; gill-filament 9.1 - 11.
Proportions as % eye diameter: gill-raker 32-45; gill-filament 49-56 at 173-198 mm, 63-69 at 261-342 mm and 76 at 433 mm SL. Scales cycloid on snout and below eye, otherwise finely ctenoid. Lateral line evenly curved. Caudal fin elongate pointed in small specimens, S-shaped in those of medium size, and according to Trewavas (1977), bi-concave ("symmetrically concave") in very large individuals. Drumming muscles well developed in both sexes. Urinary bladders large, as in A. japonicus.
Figure 11. Swim-bladder
appendages (anterior view) from
mid-way along left side of swim-bladder: A) Argyrosomus hololepidotus,
465 mm SL, neotype;
Otoliths (Fig. 10): Description as for A. japonicus; OCD 24-34% OH.
COMPARISONS: Specific differences between A. regius and the congeners described in this paper are dealt with under the "comparisons" sections of the respective species (below) and in Tables 2-6 (see end of paper). DISTRIBUTION: A. regius occurs throughout the Mediterranean and Black Sea, northwards along the Atlantic coasts of Europe to the British Isles, southern Norway and Sweden; it has been recorded along the west coast of Africa to the Gulf of Guinea (Trewavas, 1977; Chao & Trewavas, 1981; Seret, 1981; Chao, 1986). A. regius has entered the Red Sea through the Suez canal (Steinitz, 1967; Mohan, 1984). The species occurs in inshore and shelf waters, close to the bottom or near the surface (depth range 15-200 m); it also enters estuaries and coastal lagoons (Chao, 1986). FISHERIES: A. regius is an important food fish in the Bay of Biscay (French Atlantic) and along the coasts of Senegal and Mauritania (Moal, 1957; Balguerias, 1985; Quero & Vayne, 1987). It is caught with hook and line, trawl nets, gillnets, circular nets and seines. The flesh is sold fresh, salted or dried. BIOLOGY: Although sexual maturity is attained by some individuals at 80 cm TL, most adults off the west coast of Africa are larger than 110 cm TL (Moal, 1957). Spawning occurs during May to July in the Bay of Biscay, from April to July in the southern Mediterranean (Chao, 1986) and from January to May off west Africa (Mauritania, Moal, 1957; Tixerant, 1974 in Champagnat & Domain, 1978). Both adults and juveniles are migratory, moving longshore or offshore/onshore in response to change in water temperature (Moal 1957; Champagnat & Domain, 1978; Oliver & Lafon, 1981; Quero & Vayne, 1987; Quero, 1989). The largest size recorded is 182 cm TL and 103 kg (Quero & Vayne, 1987). MATERIAL EXAMINED:
MEDITERRANEAN SEA: Israel: HUJ 6906 (F 325 mm); HUJ 14451 (J 173
mm); HUJ 16342 (F 198 mm); RUSI 30254 (M 261 mm); RUSI 30255 (M 269 mm);
RUSI 30256 (F 227 mm). Egypt: HUJ 5182 (M 255 mm & 300 mm); MNHN 4978 (F
222 mm); MNHN 7495 (J 189 mm); MNHN 7533 (M 264 mm, 286 mm [gonads
missing]); MNHN 1524 (286 mm [gonads missing]).
Argyrosomus japonicus (Temminck & Schlegel, 1843)
Sciaena antarctica Castelnau, 1872: 100, Pl. 1 (Melbourne market; Macleay, 1881: 520; Tenison-Woods, 1883: 53, Pl. 16; Stead, 1906: 113, Fig. 42; Ogilby, 1908: 66, Pl. 37; Roughley, 1916: 112, PI. 35, 195.1: 70, Pl. 28; McCullock, 1922: 58, Pl. 58; Waite, 1923: 129. (Australia). Sciaena margaritifera Haly, 1875:269 (off Natal, South Africa). Sciaena aquila (non Lacepede): Castelnau, 1878: 381.
Corvina axillaris (non Cuvier): De Vis, 1884:538 (Brisbane River; so synonymized by Ogilby, 1918). Sciaena neglecta Ramsey & Ogilby, 1886:941 (coast of New South Wales; so synonymized by Ogilby, 1918). Sciaena hololepidota antarctica: (non Lacepede); Ogilby, 1918: 70, Pl.21 (Queensland). Nibea japonica: Matsubara, 1937: 41, Figs 9 & 10 (near Tokyo); Chu, Lo & Wu, 1963: 55, Figs 31, 57, 83; Taniguchi, 1969: Figs 4 A & B; Masuda et al. 1988: Pl. 147 E. Argyrosomus japonicus: Lin, 1938: 170; Lin, 1940: 244, Fig. 1 (Hong Kong); Matsubara, 1955: 645, Pl. 67, Fig. 229 (Japan); Chu, 1956: 25, Pl. 2, Fig. 3 (Taiwan); Kamohara, 1964:50 (Japan); Fischer & Whitehead, 1974: SCIAEN Argyr 4; Sasaki, 1994.
Johnius hololepidotus (non Lacepede): Smith, 1949: 226, Fig. 552; Smith & Smith, 1966: 43.
Pseudosciaena antarctica: Ogilby & Marshall, 1954: 70, Fig. 85 (Queensland).
Argyrosomus hololepidotus (non Lacepede): Smith & Smith, 1966: 43; Fischer & Whitehead, 1974: SCIAEN Argyr 3; Trewavas, 1977: 326, Figs. 9, 12, 47 & 54 (Australia & South Africa); van der Elst, 1981: 261; Mohan, 1984: SCIAEN Argyr 3; Heemstra, 1986: 616, Pl. 71; Grant, 1987: 221, Fig. 493; Kailola et al. 1993: 318; Kuiter 1993: 199; Gomon, 1994; Branch et al. 1994: 250. Johnius antarctica: Grant, 1978: 474, Pl. 201 (Australia). Argyrosomus sp.: Sainsbury et al., 1984: 230, fig. Neotype: RUSI 44704, 499 mm SL, Meitsu, Nango-Cho, Miyazaki Prefecture, southeast coast of Kyushu, Japan, depth 50 m; 21st October 1993. Collected by Yukio Iwatsuki. DIAGNOSIS: A species of Argyrosomus with the following combination of characters: dorsal fin rays 25-30; gill-rakers 4-5 + 10-12; swim-bladder carrot shaped, with 21-31 arborescent appendages; caudal peduncle length 11-13 % SL; peduncle depth 70-92% peduncle length; interorbital width 6.4-7.4% SL (21-24% HL); suborbital width 8.4-10.5% HL; lower jaw length 48-52% HL; midlateral body scale length 2.4-3.1% SL; drumming muscles present in both sexes; urinary bladder length 2.0-9.6% SL; otolith shape like A. regius (Figs 13-15; see description below), OCD 23-41% OH; pectoral fins 17-21% SL, usually not reaching vertical at tips of pelvic fins; pectoral axillary fold scaleless; peritoneum unpigmented; anterior part of lateral line slightly curved. DESCRIPTION: Based on 66 preserved specimens (119-638 mm), seven skeletons (318-590 mm) and the otoliths of 543 fish that were not preserved. Data from the neotype are in parenthesis. Dorsal fin X+I,25-30 (X,29); anal fin II,7; pectoral-fin rays 15-17 (17); lateral-line scales 50-53 (51); gill-rakers 4-5 + 9-12 (4 + 9); swim-bladder appendages 21-31 (30); vertebrae 11 + 14. Proportions as % SL: head length 28-33 (28.5); pre-dorsal length 32-36 (33); pre-pelvic length 31-36 (31); body depth 25-30 (25); caudal peduncle length 11-13 (13); peduncle depth 8.8-10.6 (9.0); snout length 7.7-9.0 (8.3); eye diameter 5.2-7.0 at 119-298 mm, 4.4-5.2 at 301-387 mm and 3.9-5.0 (3.9) at 403-638 mm SL; interorbital width 6.4-7.4 (6.8); suborbital width 2.6-3.2 (2.6); upper jaw 12-13.5 (12); lower jaw 13.5-16 (14.0); pectoral fin 17-21 (17); pelvic fin 17-22 at 119-298 mm, 17-20 at 301-387 mm and 15-18 (16) at 403-634 mm SL; anal fin 15-19 at 119-298 mm, 15-17 at 301-387 mm and 12-15 (14) at 403-638 mm SL; third dorsal spine 11-15 (12); second anal spine 7.5-12 at 119-298 mm and 5.7-7.9 at 301-638 mm SL; gill-raker 1.8-2.8 at 119-298 mm, 1.8-2.2 at 301-387 mm and 1.6-2.1 (2.0) at 403-638 mm SL; gill-filament 2.6-3.4 (2.9).
Proportions as % HL: snout 26-30 (29); eye diameter 16-23 at 119-298 mm, 15-18 at 301-387 mm and 13-16 (13.6) at 403-638 mm SL; interorbital width 21-24 (24); suborbital width 8.5-10.4 (9.2); upper jaw 40-45 (42.5); lower jaw length 48-52 (49); gill-raker 6.2-8.9 at 119-298 mm, 6.3-7.1 at 301-387 mm and 5.3-9.0 (6.9) at 407-664 mm SL; gill-filament 8.5-11.4 (10.1). Proportions as % eye diameter: gill-raker 32-57 (51); gill-filament 45-63 at 130-298 mm, 55-68 at 301-387 mm and 57-80 (74) at 407-664 mm SL. Dorsal profile generally has a hump or "shoulder" behind the head. Axillary fold at base of pectoral fin scaleless. Scales weakly ctenoid on body (including belly and chest, nape, interorbital region, preoperculum and below eye; those on body not easily lost; length of midlateral body scales 2.4-3.1% SL. Lateral line evenly curved. Pectoral fin of most fish less than 100 cut not reaching vertical at tip of pelvic fins, but on some fish the pectoral and pelvic tips are equal. Caudal fin typically pointed (Fig. 2A) in fish less than 10 cm, becoming elongate S-shaped (Fig. 2B) in fish 15-35 cm SL and S-shaped (Fig. 2C) in fish larger than 40 cm SL; the caudal fin of large adults is occasionally bi-concave, wedge shaped or slightly rounded (Fig. 2F). Drumming muscles well developed in males and females but thicker in mature males than in mature females; although poorly developed in small individuals, drumming muscles were observed in all specimens greater than 27 cm. Peritoneum not pigmented. Urinary bladders large (Fig. 1). Pyloric caeca 7-10 (n= 64).
DEA = Disproportionately Enlarged Appendage. Scale bar = 10 Figure 14. Lateral view of swim-bladders and anterior view of middle appendage: A) Argyrosomus inodorus, 640 mm, RUSI 38480; B) A. japonicus, 638 mm, RUSI 38452. DEA = Disproportionately Enlarged Appendage. Scale bars = 10 mm.
Figure 16. Lateral and ventral views of some sagittae illustrated in Figure 15. Scale bar Figure 17. Intraspecific variation of sagittae from South African Argyrosomus japonicus and similar-sized specimens from Japan and Australia.
Neurocranium (Figs 4 & 5): Length of neurocranium (NCL) 20-22% SL. Pre-supraocciptal length 10-11% SL and 49-53% NCL. Supraoccipital crest length 9.2-10.2% SL and 43-49% NCL (318-560 mm). Neurocranial width 7.3-8.4% SL and 34-39% NCL. Premaxilla 8.1-8.9% SL and 39-42% NCL. Dentary 9.5-10.1% SL and 44-48% NCL. Third and fourth neural spines 7.5-8.3 and 8.9-9.9% SL respectively.
Colour: In life, silvery grey becoming darker above and lighter below; dorsal surface with a bluish bronze sheen that may become coppery on head. Larger fish (>80 cm) are golden brown; dorsal and caudal fins grey/brown, becoming darker with age, often with a reddish hue - due to vascular distension or haemorrhaging (possibly during capture). Pectoral and anal fins white in fish less than 10 cm, changing to yellow and finally to grey or grey/brown in fish greater than 35 cm, also becoming darker with size. Mouth lining of juveniles white, becoming yellow and finally yellowish grey in large adults. Axillary fold of pectoral fin grey to black, generally with an irregular coppery-bronze patch. Preserved specimens vary in colour according to treatment; usually grey but sometimes brown; darker above and lighter below. Dark patches on scales may give appearance of oblique stripes. Fins grey, sometimes with one or two longitudinal stripes on dorsal fin; the pelvic and pectoral fins usually pale.
REMARKS: The original description of Temminck and Schlegel (1843: 58, Pl. 24, Fig. 1) was based on the description and plate in Burger's unpublished manuscript. According to Boeseman (1947: 62) "there never was a specimen in Burger's or Von Siebold's collection,". Boeseman (1947: 62) also pointed out "that Burger's descriptions without exception proved to be rather inexact. Temminck and Schlegel generally corrected and completed Burger's data with characters from the specimen(s) [from Burger's collection] ... Burger's plates too often show inaccuracies, but these also sometimes have been corrected by the authors [Temminck & Schlegel] ...In this case, however, there was no way to verify Burger' s data and plate. On account of this, it is very difficult, if not impossible, to make a final statement as to the identity and synonymy of Temminck & Schlegel's species." In view of the uncertainty of the identity of Sciaena japonica from Temminck & Schlegel's original description and illustration, the absence of a type specimen, and the presently confused taxonomy of Argyrosomus species, we believe that designation of a neotype for Sciaena japonica will promote stability of nomenclature and help to resolve the taxonomic confusion associated with these species.
Trewavas (1977: 330) commented: "As Lin (1940) pointed out, the resemblance [of A. japonicus] to A. hololepidotus is striking. The apparent differences are mostly correlated with size and are due to allometry." Trewavas' concept of "A. hololepidotus" was based on South African and Australian specimens of A. japonicus, as she did not examine any specimens from Madagascar. Her account of A. japonicus was compiled from published descriptions of specimens from Japan and China, and she went on (1977: 331) to observe that "there is no inherent improbability in Lin's suggestion (1940) that the relationship between A. japonicus and A. hololepidotus may be that of races of one species".
Our comparison of specimens from South Africa, Japan and Australia confirms that these populations are conspecific. Previous recognition of "A. hololepidotus" and A. japonicus as separate species was due to taxonomic inertia and the fact that none of the previous authors had directly compared specimens from Japan with those from South Africa or Australia. Although the South African, Japanese and Australian populations of A. japonicus seem to be well isolated geographically (see below), and the smaller size of Australian specimens (see below) implies some genetic differences between South African and Australian populations, the absence of any significant morphological (including branching patterns in the swim- bladder appendages) or meristic differences in the specimens that we have examined from these populations requires that we recognize them as single species. Examination of molecular characters (proteins, DNA, etc) or some morphological features that we may have overlooked may show significant differences between these allopatric populations; but until such evidence is discovered, we accept the null hypothesis of conspecific identity of these populations by default (i.e., the burden of proof for recognition of separate species requires some tangible evidence other than apparent allopatry). DISTRIBUTION: A. japonicus is found on the African southeast coast from the Cape of Good Hope to southern Mozambique; in Australia it is known along the entire southern seaboard from North West Cape (north of Shark Bay) in Western Australia to the Burnett River (north of Brisbane) in Queensland (Trewavas, 1977; Grant, 1978; Sainsbury et al., 1984; Kailola et al., 1993; Starling, 1993); in the northern Indian Ocean, it occurs off Pakistan and the northwest coast of India (Mohan, 1984) (as " A. hololepidotus"); and in the Northern Pacific it has been reported from Hong Kong northwards along the Chinese coast to southern Korea and Japan (Trewavas, 1977).
Along the South African coast A. japonicus occurs abundantly from Cape Agulhas to northern Natal (Table 7 - see end of paper). It is found in estuaries (preferring those that are turbid), in the surf zone, and in the nearshore zone to depths of about 50 metres. In the nearshore environment, the species rarely ventures onto the "soft" (flat) substrates exploited by trawlers, but prefers "linefish grounds" (especially wrecks or high profile reefs that are associated with sand). West of Cape Agulhas, the relative proportion of A. japonicus found in Argyrosomus catches declines markedly, and the species is not known in the cold waters from the Cape of Good Hope to Namibia north of Luderitz (Table 7). FISHERIES: Due to its large size, palatability and abundance, A. japonicus is perhaps the premier angling species of the inshore environment (estuaries and surf zone) along the South African east coast (Cape Agulhas to Natal). Commercial line boat fishermen operating in the nearshore environment catch an average of 197 tons per annum (1988-1992). Recreational boat-fishermen often target this species and are believed to catch at least as much, and possibly more than the commercial fishermen. The flesh is generally sold fresh, but sometimes frozen.
In Australia A. japonicus (previously identified as Sciaena antarctica, Johnius antarctica, or more recently as "Argyrosomus hololepidotus") is an important commercial and recreational angling species known as the "mulloway". It is caught in estuaries, in the surf and offshore, using hook and line, gill nets, seine nets and trawl nets (Roughley, 1951; Ogilby, 1954; Grant, 1978; Hall, 1985; Jones, 1991; Kallola et al. 1993; Starling, 1993). From 1964 to 1990, the annual Australian catch has fluctuated between 175 and 575 tons (Kailola et al., 1993). BIOLOGY: In South African waters, 50% sexual maturity for A. japonicus is attained at 110 cm TL for females and at 95 cm TL for males (Griffiths & Hecht, 1993 [species A]). Although there is strong evidence for a spawning migration of adult fish to Natal in spring, reproductive activity is also observed in the Cape Province (as far west as Cape Infanta). Adult fish are found mainly in the nearshore environment (beyond the surf zone) but at times visit inshore habitats. Juvenile fish (<100 cm TL) remain exclusively in the inshore habitats (surf and estuaries) with small juveniles (< 15 cm TL) found only in estuaries. Juvenile recruitment to estuaries occurs at about 30 mm TL. The maximum size recorded is 75 kg (181 cm TL). Fish of 45 kg are common, and those between 50-60 kg are not unusual.
In Australia, A. japonicus attains sexual maturity at 75 cm TL, and spawning apparently occurs in the summer months (Hall, 1986). According to Kailola et al. (1993), the maximum size in Australia is 43 kg, but Gomon (1994) states that the mulloway attains a length of over 2 metres and a weight of nearly 60 kg. MATERIAL EXAMINED:
JAPAN: BMNH 1987.5.7.1 (M 297 mm); BMNH 1987.5.7.2 (M 339 mm); BMNH
1987.5.7.3 (F 268 mm). Argyrosomus hololepidotus (Lacepede, 1801)
Sciaena aquila (non Lacepede): Pellegrin 1914:225 (Madagascar). Argyrosomus hololepidotus: Bauchot & Bianchi, 1984: 88; Mohan, 1984, SCIAEN Argyr 3 (in part). Holotype: none; Iconotype: Lacepede, 1801, Pl. 21, Fig. 2 (from Philibert Commerson's drawing). Neotype: RUSI 41433, female 465 mm, Fort Dauphin, Madagascar; collected by Marc H. Griffiths, 3rd April 1993. According to Cuvier and Valenciennes (1830), Commerson's original specimen from Fort Dauphin, Madagascar and his description of the species were lost. Because sciaenid taxonomy depends heavily on internal anatomy (Trewavas, 1977), the iconotype (an engraving of Commerson's figure) contains too little detail to differentiate between Argyrosomus species. It is therefore necessary to designate a neotype to resolve the taxomomic confusion that besets this name. ADDITIONAL MATERIAL: RUSI 41432, J 204 mm, and a fresh specimen (M 615 mm) examined at the Fort Dauphin fish market. Meristic data from the third specimen were recorded, and the otoliths were collected; otoliths of four larger specimens were also collected. DIAGNOSIS: A species of Argyrosomus with the following combination of characters: dorsal fin rays 27-28; gill-rakers 4-5 + 9; swim-bladder carrot shaped, with 34-36 arborescent appendages with lateroanterior limbs throughout length of bladder; caudal peduncle length 12-13% SL; peduncle depth 75-77% peduncle length; interorbital width 6.3-7.3% SL (22% HL); suborbital width 11-12% HL; lower jaw length 50% HL; midlateral body scale length 2.5-2.8% SL; drumming muscles present in both sexes; sagitta similar to that of A. regius, OCD 28-34% OH; pectoral fins 17-18% SL, not reaching vertical at tip of adpressed pelvic fins; pectoral fin axillary fold scaleless; peritoneum unpigmented; anterior part of lateral line moderately curved. DESCRIPTION: Data for the neotype in parentheses. Dorsal fin X+1,27-(28); anal fin 11,7; pectoral fin rays 16-(17); lateral-line scales 50 - (51); gill-rakers 4-(5) + 9; swim bladder appendages (34)-36; vertebrae 11 + 14.
Proportions as % SL: head length (29)-30; pre-dorsal length 34; pre-pelvic length 32; body depth (26)-27; caudal peduncle length (12)-13; peduncle depth (9.0)-9.7; snout length (7.4)-7.8; eye diameter (4.7)-5.5; interorbital width (6.3)-6.7; suborbital width (3.0)-3.4; upper jaw (12)-12.5; lower jaw (14)-15; pectoral fin (17)-18; pelvic fin (18)-20; anal fin (14)-16; third dorsal spine (13); second anal spine (6.7)-8.5; gill-raker (1.8)-1.9; gill-filament 2.9-(3.2); scale length (2.5)-2.8. Proportions as % HL: snout (26)-27; eye diameter (16.5)-18; interorbital width (22)-23; suborbital width (10.5)-12; upper jaw 42-(43.5); lower jaw 50; gill-raker (4.2)-6.3; gill-filament 10-(11).
Proportions as % eye diameter: gill-raker 34-(39); gill-filament 54-(68). Dorsal profile of A. hololepidotus includes a hump or "shoulder" behind the head. Pectoral fins not reaching vertical at tip of pelvic fins (for fish 204-615 mm). Scales ctenoid on body, belly, nape, interorbital region, operculum and below eye. Caudal fin elongate S-shaped (Fig. 2B) in smallest fish and S-shaped (Fig. 2C) in two larger specimens examined; it is probable (as in A. japonicus) that the caudal fins of fish less than 10 cm SL are pointed. Gill-raker at angle shorter than outer gill-filaments (at angle) in larger fish, and equal to gill-filaments in 204 mm specimen. Drumming muscles present in both sexes. Urinary bladders large, as in A. japonicus. Pyloric caeca 8 or 9.
Otoliths: Comparisons of sagittae from A. hololepidotus, A. japonicus (Figs 15 & 16) and A. regius (Fig. 11 and the illustrations of Chaine, 1938 and Schwarzhans, 1993) revealed no significant differences. The denticles observed along the dorsal margin of the smallest otolith figured for A. hololepidotus (Figs 15 & 16) are also sometimes found in small specimens of A. japonicus and A. regius. The OCD is 28-34% OH for A. hololepidotus.
Colour: Fresh colour of neotype silvery grey becoming darker and bluish above and white below; flanks and dorsal surface with a distinctly bronze sheen. Pectoral, dorsal and caudal fins brownish grey; pelvic and anal fins almost white with streaks of grey. Fleshy fold at axil of pectoral fin black and scaleless. After fixation the neotype darkened, becoming distinctly brown; this dark brown colour is possibly due to the use of paraformaldehyde (to prepare the formalin solution). Mouth lining pale grey; inner aspect of operculum dark grey. Peritoneum unpigmented. COMPARISONS (Tables 2 - 4): A. hololepidotus differs from A. japonicus by having a wider suborbital, more swim-bladder appendages and fewer lower gill-rakers. Furthermore, the branches arising from the ventral limbs along the posterior half of the swim-bladder (Fig. 20), project laterally from the bladder and do not lie against it as in A. japonicus (Figs 13 & 14), and in specimens with little or no appendage fat, the ventral limb is orientated lateroanteriorly (vs ventroanteriorly) in the posterior half of the bladder. A. hololepidotus differs from A. regius by having fewer and smaller swim-bladder appendages (Fig. 11), which have ventral limbs (in specimens with little or no appendate fat) that are orientated anterolaterally (vs anteroventrally) and branches arising from these which project at 90 degrees from the bladder (vs posteriorly and lying against the bladder); wider suborbital; shorter pectoral fin; longer scales; narrower interorbital; and longer lower jaw.
A. hololepidotus was also compared with data and illustrations of A. amoyensis from Trewavas (1977), Mohan (1984) and the original description of Nibea miichthioides Chu et al. 1963. A. hololepidotus differs in having 9 (vs 8) lower gill-rakers, interorbital width 22-23% HL (vs 19% HL), and suborbital width 11-12% HL (vs 9% HL). The swim-bladder of A. amoyensis is spindle shaped (tapering anteriorly and posteriorly), there are only 22-29 appendages, and they are not obviously divided into dorsal and ventral limbs. The sagitta of A. hololepidotus (Figs 15 & 16) is similar to that of A. amoyensis (Trewavas, 1977: Fig. 9b; Chu et al., 1963: Fig. 82), but it lacks the protuberance found on the posterodorsal corner in A. amoyensis. DISTRIBUTION: A. hololepidotus is known only from Madagascar. The neotype, Commerson's original specimen (Cuvier & Valenciennes, 1830) and the specimens listed by Pellegrin (1914) were all collected from Fort Dauphin on the south east coast of Madagascar. Although Mohan (1984) indicated that A. hololepidotus is found along the entire cast coast and northern parts of the island, fisheries personnel in Madagascar report that it is common only on the southeast coast. FISHERY: A. hololepidotus or the "fyandava" (Malagasy name) is generally caught by artisanal fishermen using dugout canoes and handlines in 20 - 40 m of water. BIOLOGY: All three specimens (204-615 mm) examined in this study were immature, suggesting a large size at sexual maturity (similar to A. japonicus and A. regius). The maximum size, as measured from marks on the oars and gaffs of fishermen, was 135 cm TL.
Argyrosomus thorpei Smith, 1977
Argyrosomus thorpei Smith 1977: 561, Pls. I & II; van der Elst, 1981: 262; Mohan, 1984: SCIAEN Argyr 7; Heemstra, 1986: 617, PI. 71. Afroscion thorpei: Trewavas, 1977: 337; Schwarzhans, 1993: 134, Figs. 244-245. Holotype: RUSI 674 (320 mm SL from Natal) Paratypes: RUSI nos 675-680, 2651A & B, BMNH 1976.10.18.1 DIAGNOSIS: A species of Argyrosomus with the following combination of characters: dorsal fin rays 26-28; gill-rakers 4-6 + 10-12; swim-bladder carrot shaped, with 25-33 arborescent appendages; caudal peduncle length 12-14% SL; peduncle depth 65-79% peduncle length; interorbital width 6.0-7.9% SL (19-26% HE); suborbital width 10-12% HL; lower jaw length 47-52% HL; midlateral body scale length 2.6-3.3% SL; drumming muscles present in both sexes; urinary bladders large; sagitta with anterior two-thirds expanded, posterodorsal angle >90 degrees, posterior margin straight, cauda with distal portion expanded and tip truncate, OCD 34-44% OH; pectoral fins 20-23% SL, reaching past vertical at tip of pelvic fins; pectoral fin axillary fold scaly; peritoneum pigmented; anterior half of lateral line strongly curved. DESCRIPTION: Based on 16 preserved specimens (166-474 mm), two fresh specimens, two skeletons (452 & 461 mm) and otoliths of 53 other fish. Dorsal fin X+I,26-30; anal fin II,7; pectoral-fin rays 15-17; lateral-line scales 49-52; vertebrae 11 + 14. Proportions as % SL: HL 29-32; pre-dorsal length 33-37; pre-pelvic length 32-34; body depth 24-30; caudal peduncle length 12-14; peduncle depth 8.4-10.2; snout 8.1-8.9; eye diameter 6.8-7.9 at 166-23 8 mm, 4.8-5.4 at 315 -390 mm and 4.0-4.2 at 441-474 mm SL; interorbital width 6.0-7.9; suborbital width 3.0-3.7; upper jaw 12-14; lower jaw 14-16; pectoral fin 20-23; pelvic fin 16-21 at 166-238 mm, 17-18 at 315-390 mm and 16-17 at 403-634 mm SL; anal fin 15-17 at 166-238 mm, 14 at 315-390 mm and 13 at 460 mm SL; third dorsal spine 10-15; second anal spine 4.4-7.9 at 166238 mm and 6.7-7.7 at 315-474 mm SL; gill-raker 2.7-3.0 at 166-238 mm, 2.4-3.0 at 315-390 mm and 2.3-2.5 at 441-474 mm SL; gill-filament 2.6-3.4. Proportions as % HL: snout 26-29; eye diameter 19-21 at 166-238 mm, 17-19 at 315-390 mm and 15-17 at 403-474 mm SL; interorbital width 19-26; suborbital width 10-12; upper jaw 39-44; lower jaw 47-52; gill-raker 9.0-9.8 at 166-238 mm, 8.1-10.0 at 315-390 mm and 8.1-8.5 at 441474 mm SL; gill-filament 9.1-11.6.
Proportions as % eye diameter: gill-raker 42-56; gill filament 51-55 at 166-238 mm, 53-68 at 315-390 mm and 66-74 at 441-474 mm. Scales cycloid on snout, below eye, on interorbital region and on extreme anterior part of chest; ctenoid on rest of body. Anterior half of lateral line strongly curved. Caudal fin elongate S-shaped in juveniles, becoming slightly S-shaped and occasionally truncate in adults (Fig. 2). The correction made to the caudal fin in the photograph of the holotype (Smith, 1977) is probably erroneous. Gill-raker at angle of first arch at least equal to outer (shorter) gill-filaments in fish less than 46 cm, but gill-raker shorter than gill-filaments in larger fish. Smith (1977) reported no sonific muscles for A. thorpei, but we found drumming muscles in both sexes (n=48) in specimens larger than 23 cm (including the paratypes listed). Urinary bladders large (as in A. japonicus).
Swim-bladder carrot shaped, with 25-33 appendages (for illustration see Smith 1977, P1.2); the ventral limb is orientated lateroanteriorly, rather than venteroanteriorly and the branches arising from it project laterally and at no stage lie against the bladder The dorsal limb is orientated dorsoposteriorly, but along the posterior three quarters of the bladder it curves through 150 degrees so that the tip points anteriorly. The branches on this limb project posteriorly to dorsally; and lie against the swim-bladder; last 1-3 appendages are bud-like and the 1-3 preceding appendages weakly branched. Disproportionate swim-bladder enlargement not observed in any A. thorpei that we examined.
Otoliths: Sagittae of A. thorpei (Figs 15 & 16) very distinctive, moderately elongate, the anterior two-thirds expanded, resulting in a broadly rounded apex; dorsal and posterior margins relatively straight; posterodorsal angle >90 degrees; ventral margin convex in smaller fish, becoming concave with growth; cauda strongly curved with distal portion expanded and tip truncate.
Neurocranium (Figs 4 & 5) and skeleton: Length of neurocranium (NCL) 20-21% SL; pre-supraoccipital crest length 10-11% SL and 49-51% NCL; supraoccipital crest length 8.5-8.9% SL and 41-44% NCL (452-461 mm SL); neurocranial width 8.1-8.4% SL and 40-41% NCL. Premaxilla 8.6-8.7% SL and 42-43% NCL; dentary 9.8-10% SL and 48% NCL; interorbital septum relatively unossified. Third and fourth neural spines 8.9% and 8.5-8.7% respectively.
Colour: Silvery grey becoming darker and blue (with a coppery sheen) above and lighter below; fins yellow grey to orange brown, occasionally reddish (particularly caudal) as a result of vascular distension and haemorrhaging; mouth lining pale yellow; inside of operculum grey/black with yellow tinge; peritoneum pigmented with grey/black dots. After preservation the colour depends on treatment, and may vary from dark brown to grey, or even silver. A dark spot on each dorsal body scale may give the appearance of oblique stripes; fins greyish or brown; peritoneum and ventral surface of swim-bladder with fine black/grey dots. COMPARISONS (Tables 2-6): The body of Argyrosomus thorpei is slightly more compressed than the other species described in this paper. Teeth of lower jaw with Argyrosomus pattern of differentiation, but smaller than in equal- sized specimens of the other species (Fig. 6). Differences between A. thorpei and A. japonicus include: a wider suborbital, scaly pectoral fin axillary fold, anterior half of lateral line more strongly curved, longer caudal peduncle, longer gill-rakers; sagitta with a more obtuse posterodorsal corner, anterior two-thirds of otolith and distal portion of the cauda expanded (Fig. 15); neurocranium wider, with wider lateral line canals (Fig. 5) and larger inter-orbital window (Fig. 4).
A. thorpei differs from A. hololepidotus in having fewer swim-bladder appendages, more lower gill-rakers, longer pectoral fin, anterior half of lateral line more strongly curved, scaly pectoral fin axillary fold, pigmented peritoneum; sagitta with a more obtuse posterodorsal corner and anterior two-thirds of otolith and distal portion of the cauda expanded (Fig. 15). REMARKS: Based on otolith shape, Trewavas (1977) established the monotypic genus, Afroscion, for Argyrosomus thorpei Smith, 1977. This genus was not recognized by Mohan (1984), Heemstra (1986) or Sasaki & Kailola (1988). The recent discovery of Afroscion trewavasae Schwarzhans 1993, based on otoliths from the Lower Pliocene of Morocco, led Schwarzhans to reinstate the genus Afroscion, arguing that it represents a lineage separated from Argyrosomus for a considerable period of time. According to him, "The shape of the dorsal rim and the absence of a concavity at the ventral rim (of the otolith) are the most obvious diagnostic differences" for the genus. Schwarzhans (1993), however, described the dorsal margins ("rims") of both genera as being flat, and we found that the ventral margin of Argyrosomus sagittae becomes concave with growth (Fig. 15). Although A. thorpei differs from other species of Argyrosomus by its scaly pectoral axillary fold, pigmented peritoneum, strongly curved lateral line, and distinctive otolith morphology, we believe these differences are insufficient to justify a separate generic status. DISTRIBUTION: A. thorpei is known from the southeast African coast from Port Elizabeth to Mozambique and also from the west coast of Madagascar. In South Africa it is mostly caught beyond the surf zone (to depths of 80 m) and seldom found south of Natal (Table 7). FISHERIES: Natal commercial lineboat fishermen catch an average of 107.1 tons and the recreational line-boat sector land an estimated 23 tons (ORI unpublished data) per annum (1988-1992). Shore anglers occasionally catch this species at selected sites on the Natal north coast. A. thorpei is also caught in large numbers by line-boat fishermen operating in Mozambique. BIOLOGY: The size at 50% sexual maturity is 33 cm TL, and spawning occurs during winter (van der Elst et al., 1990). Juveniles are found on sand or mud substrata and the adults predominantly on reef (Fennessy, 1994). Maximum size recorded is 12.0 kg. MATERIAL EXAMINED:
SOUTH AFRICA: Natal: RUSI 675 (M 365 mm); RUSI 676 (F 365 mm); RUSI
677 (M 474 mm); RUSI 678 (M 375 mm); RUSI 680 (F 315 mm); RUSI 2651 (M 390
mm); RUSI 10264 (F 385 mm); RUSI 10694 (J, 166 mm and 2F, 171 & 185 mm);
RUSI 11613 (F, 238 mm); RUSI 13065 (1 M 460 mm and 2 F, 379 mm & 441 mm);
RUSI 28382 (M 23 1 mm). Plus otoliths from 30 fish (14 - 110 cm TL).
Eastern Cape Province: Algoa Bay: RUSI 10264 (M 385 mm).
Argyrosomus inodorus sp. nov.
Johnius hololepidotus (non Lacepede): Smith, 1949:226 (in part, not Fig. 552, which is A. japonicus). Argyrosomus hololepidotus (non Lacepede): Trewavas, 1977:327 (in part, not figs); Heemstra, 1986:616 (in part, not figs); Bianchi et al., 1993: 164; Schwarzhans, 1993: 138, Fig. 255. Holotype: RUSI 44705, male, 447 mm SL, South Africa, False Bay, depth 10 m; collected by M.H. Griffiths, 5 June 1994. Paratypes: 78 specimens (131-860 mm). NAMIBIA: Walvis Bay: RUSI 15257 (M 605 mm). Swakopmund: RUSI NP (M 392 mm); RUSI 48475 (F 415 mm); RUSI 48476 (F 424 mm); RUSI 48477 (M 338 mm); RUSI 48478 (F 332 mm); RUSI 48479 (M 315 mm). SOUTH AFRICA: Eastern Cape Province: AMS I. 34965-001 (F 462 mm, M 308 mm); BMNH 1994.11.30:2 (M 305 mm); BMNH 1994.11.30:3 (F 266 mm); MNHN 1994-0568 (M 558 mm); MNHN 1994-0569 (F 307 mm); NSMT-P 46548 (M 348 mm); NSMT-P 46549 (F 521 mm); RUSI 11924 (F 470 mm); RUSI 38464 (F 306 mm); RUSI 38466 (F 274 mm); RUSI 38467 (F 330 mm); RUSI 38469 (F 283 mm); RUSI 38471 CF 304 mm); RUSI 38474 (F 374 mm); RUSI 38476 (F 258 mm); RUSI 38478 (F 289); RUSI 38479 (M 299 mm); RUSI 38480 (F 325 mm); RUSI 38481 (F 265 mm); RUSI 41138 (F 368 mm); RUSI 41141 (F 293 mm); SAM 33473 (F 276 mm) ; SAM 33474 (M 381 mm); USNM 331636 (F 357 mm, M 296 mm). Western Cape Province: RUSI 37390 (J 131-157 mm); RUSI 38475 (F 275 mm); RUSI 39836 (5 M 211-287 mm & 3 F 212-227 mm) ; RUSI 41129 (F 301 mm); RUSI 41132 (M 308 mm); RUSI 41133 (F 362 mm); RUSI 41134 240 mm) ; RUSI 41136 (F 369); RUSI 41139 (J 154 mm) ; RUSI 41140 (M 366 mm); RUSI 41143 (M 306 mm); RUSI 41145 (M 364 mm) ; RUSI 41147 (F 267 mm); RUSI 41148 (F 219 mm); RUSI 41149 (F 267 mm) ; RUSI 41150 (F 202 mm) ; RUSI 41151 (F 235 mm) ; RUSI 41153 (F 234 mm); RUSI 41154 (J 167 mm) ; RUSI 41156 (F 407 mm); RUSI 41160 (J 179 mm) ; RUSI 41161 (M 595 mm); RUSI 41162 (J 161 mm); RUSI 41163 (F 227 mm) ; RUSI 41167 (F 234 mm); RUSI 41435 (J 141 mm); RUSI 42241 (M 660 mm) ; RUSI 42242 (F 510 mm) ; RUSI 42246 (M 664 mm) ; RUSI 42240 (F 640 mm) ; RUSI 42243 (F 527 mm) ; RUSI 42244 CM 387 mm); RUSI 42245 (M 383 mm) ; RUSI 42247 (F 650 mm); RUSI 42248 (F 860 mm) ; RUSI 44706 (453 mm); RUSI 44707 (435 mm). DIAGNOSIS: A species of A. argyrosomus with the following combination of characters: dorsal fin rays 25-29; gill-rakers 4-6 + 10-12; swim-bladder carrot shaped with 31-42 arborescent appendages, the latero-anterior limb of appendages present throughout length of bladder; caudal peduncle length 12-15% SL; peduncle depth 58-74% peduncle length; interorbital width 6.3-7.8% SL (20-25% HL); suborbital width 8.6-10.9% HL; lower jaw length 47-51% HL; midlateral body scale length 1.8-2.5% SL; drumming muscles present only in males; urinary bladders rudimentary, length 0.1-3.3% SL; sagittae (Figs 15, 16 & 22) elongate, with pronounced bulge in posterior third of ventral margin; OCD 37-63% OH; pectoral fins 19-23% SL, reaching to or beyond vertical at tip of adpressed pelvic fins of fish 25-90 cm SL (in South African specimens); pectoral fin axillary fold naked; peritoneum unpigmented; anterior part of lateral line slightly curved. DESCRIPTION: Based on 79 preserved specimens (131-860 mm), 12 skeletons (300-517 mm) and otoliths and total lengths of 623 fish from South Africa, and 85 from Namibia, that were not preserved. Counts and measurements for the holotype are in parentheses. Dorsal fin X+I,25-29 (27); anal fin 11,7 [one specimen from Walvis Bay with 8 rays]; pectoral fin rays 16-17 (17); lateral-line scales 50-53 (51); gill-rakers 4-6 + 10-12; swim-bladder appendages 31-41; vertebrae 11 abdominal and 14 caudal. Proportions as % SL: HL 28-33 (30); pre-dorsal length 32-36 (33); pre-pelvic length 30-36 (34); body depth 24-28 (25.5); caudal peduncle length 12-15 (13.5); peduncle depth 7.7-9.6 (9.2); snout 7.1-9 (8.5); eye diameter 5.8-7.3 at 131-299 mm, 4.9-6.2 at 301-387 mm, 4.0-5.0 (5.0) at 404664 mm, and 3.3 at 860 mm SL; interorbital width 6.3-7.8 (7.2); suborbital width 2.6-3.3 (3.0); upper jaw 12-14 (12); lower jaw 14-18 (14.5); pectoral fin 19-23 (21.6); pelvic fin 16-20 at 131-387 mm, 15-17 (17) at 407-664 mm, and 14 at 860 mm SL; anal fin 14-18 at 131-299 mm, 13-15 at 301-387 mm, 12-14 (13.9) at 407-664 mm and 12 at 860 mm SL; third dorsal spine 11-15 (12); second anal spine 5.1-11.7 (7.2); gill-raker 2.0-2.9 at 131-299 mm, 1.8-2.6 at 301-387 mm, 1.6-2.1 at 407-664 mm and 1.2 at 860 mm SL; gill-filament 2.6-3.6.
Proportions as % of HL: snout 24-30 (28.5); eye diameter 18-24 at 131-299 mm, 16-20 at 301-387 mm, 13-17 (17) at 407-664 mm SL, and 11 at 860 mm SL; interorbital width 20-25 (24); suborbital width 8.6-10.9 (10.1); upper jaw 39-44 (40); lower jaw 47-51 (49); gill-raker 6.7-9.1 at 131-299 mm, 6.0-8.8 at 301-387 mm, 5.3-7.0 at 407-664 mm and 3.9 at 860 mm SL; gill-filament 8.9-13.3.
Proportions as % eye diameter: gill-raker 3246; gill-filament 37-53 at 131-299 mm, 46-77 at 301-3 87 mm, 5 5-88 at 407-664 mm and 107 at 860 mm SL.
Dorsal profile evenly rounded. Eye obliquely egg shaped. Nostrils as for A. japonicus. Scales deciduous, weakly ctenoid on body (including belly and chest, nape of neck, interorbital region, operculum and below eye. Skin fold at base of pectoral fin scaleless. Pectoral fins reach to or beyond vertical at tip of adpressed pelvic fins for South African specimens 25-90 cm SL, but on smaller fish the pelvic fins extend past tip of pectorals. Caudal fin shape changes with growth; it may be pointed (<10 cm SL, Fig. 2A), elongate and S-shaped (10-22 cm SL, Fig. 2B), or strongly to slightly S-shaped (>30 cm, Fig. 2C). Caudal fin of larger specimens occasionally wedge-shaped to slightly rounded or truncate (Figs 2 E, F & G). Gill-raker at angle of first arch equal to or longer than outer gill-filaments at angle for specimens less than 55 cm, but gill-filaments longer than gill-raker at angle in larger individuals. Drumming muscles present only in larger males, and are poorly developed (or even absent) in most males less than 25 cm SL (n= 8350). Urinary bladders (Fig. 1) rudimentary or sometimes absent in South African specimens, but possibly larger in those from Namibia. Pyloric caeca 7-10 (n= 335).
Swim-bladder (Figs 13A & C & 14A) carrot-shaped, tapering posteriorly, with 3142 appendages. Most appendages arborescent, basally divided into dorsoposterior and lateroanterior (rather than ventroanterior) limbs, none of which significantly overlap the bladder. Branches arising from anterior limb project at 90 degrees to bladder throughout its length and at no stage lie against it. The last 1-3 pairs of appendages are bud-like and the 1-3 pairs preceding these, weakly branched. Disproportionate appendage enlargement (see A. japonicus) was observed only in the 860 mm specimen, which was the largest of 86 A. inodorus examined. Ventral surface of swim-bladder with a sparse pattern of small black-grey dots that do not appear to increase in size or number, but disperse, with growth.
Neurocranium (Figs 4 & 5) and skeleton: Length of neurocranium (NCL) 20-22% SL; pre-supraoccipital crest length 10-12% SL and 51-56% NCL; supraocciptal crest length 8.0-8.8% SL and 39-43% NCL (300-546 mm SL); neurocranial width 8.1-9.2% SL and 3945% NCL. Premaxilia 8.1-9.1% SL and 4042 % NCL; dentary 9.3 - 10. 3 % SL and 46-49% NCL; interorbital septum relatively unossified. Third and fourth neural spines 6.9-8.0 and 8.2-9.3% SL respectively. Colour: In life, body silvery becoming green/brown above and white below. Dorsal surface with a copper to bronze sheen which may be very pronounced on head; dorsal and caudal fins pale yellow grey to pale grey/brown; anal, pelvic and pectoral fins from almost white to brown/grey; fins (particularly caudal) occasionally reddish as a result of vascular distension and haemorrhaging. Pectoral fin axillary fold black with a copper/bronze patch. Mouth lining varies from pale yellow to yellow/grey; inner aspect of operculum grey to black, sometimes with a yellow tinge. In alcohol, body silvery to grey, with a dark spot on each dorsal body scale, sometimes giving rise to oblique stripes; fins pale to dark grey. One or two longitudinal stripes may be visible on dorsal fins. COMPARISONS (Tables 1-6 & 8)(see end of paper): A. inodorus differs from A. japonicus in having a more even dorsal profile, larger eye (Fig. 23, Table 8) longer pectoral fins (Fig. 24; significant for fish >30 cm SL, Table 8), which reach to or beyond vertical at tip of pelvic fins in fish 25-90 cm SL, shorter scales (Fig. 25), which are more easily lost; a longer and shallower caudal peduncle (Figs 26-28); no drumming muscles in females; rudimentary urinary bladders (Figs 1 & 29); and a higher modal number of lower gill-rakers (Tables 4 & 5). A. inodorus also has more swim-bladder appendages (Tables 2 & 3), the appendages are larger (particularly along anterior part of bladder), with the lower limbs orientated anterolaterally (vs anteroventrally) throughout length of bladder Wigs 13 & 14), and the branches arising from the dorsal and ventral limbs of the appendages along the posterior half of the swim-bladder project at right angles from the bladder (in A. japonicus these branches extend posteriorly and lie against the bladder, Figs 13 & 14).
Figure 24. Relationship between pectoral fm length and SL for Argyrosomus inodorus (squares) and A. japonicus (circles).
Figure 25. Relationship between midlateral body scale length and SL for Argyrosomus inodorus (squares) and A. japonicus (circles).
The sagitta of A. inodorus (Figs 15, 16 & 22) is longer than that of A. japonicus (Fig. 30, Table 1), it has a more obtuse posterodorsal angle, rounded (vs straight) posterior margin, pronounced bulge on ventral margin, a greater OCD (Fig. 31, Table 1), and lower OCD:OH ratio (Fig. 32, Table 2).
The neurocranium is wider than that of A. japonicus (Fig. 33), it also has an enlarged lateral-line canal system (Figs 4 & 5), a larger interorbital window with frontal and parasphenoid bones not connected (Fig. 4), and shorter supraoccipital crest (Fig. 34, Table 2).
The fish weight/TL and otolith weight/TL relationships for A. inodorus and A. japonicus are also significantly different (P< 0.0001, Table 1). The OCD:OH ratio was not allometric for either A. inodorus or A. japonicus (see Fig. 32) and consequently should assist in the identification of these two species when only otoliths are available (e.g. studies on the feeding of Argyrosomus predators or on the middens of ancient southern African man).
Differences between A. inodorus and A. hololepidotus (Tables 2-4) include: more lower gill-rakers; narrower suborbital width; longer pectoral fin and caudal peduncle; shorter scales; reduced urinary bladders; and drumming muscles in males only. The sagitta of A. inodorus (Figs 15, 16 & 22) has a more obtuse posterodorsal angle, rounded (vs straight) posterior margin, pronounced bulge in posterior portion of ventral margin, greater OCD (Fig. 15), and a lower OCD:OH ratio (Table 2).
A. inodorus differs from A. regius (Tables 2 & 3-6) in having more lower-limb gill-rakers, narrower interorbital, longer pectoral fin, caudal peduncle and lower jaw, smaller urinary bladders, drumming muscles in males only, and lower limb of arborescent swim-bladder appendages orientated anterolaterally (vs ventroanteriorly), with branches projecting at 90 degrees to the bladder (vs projecting posteriorly and against the bladder). The sagitta of A. inodorus (Figs 15, 16 & 22) has a more obtuse posterodorsal angle, rounded (vs straight) posterior margin, pronounced bulge in posterior portion of ventral margin, greater OCD (Fig. 15), and lower OCD:OH ratio (Table 2).
A. inodorus was compared with data and illustrations of A. amoyensis from Trewavas (1977), Mohan (1984) and the original description of Nibea miichthioides Chu et al. (1963). A. inodorus has more lower gill-rakers (10-12 vs 8); a carrot-shaped (vs spindle-shaped) swim-bladder with more appendages (31-41 vs 22-29); wider interorbital (width 20-25% vs 19% ILL); and longer pectoral fin (1923% vs 17% SL). The sagitta of A. inodorus (Figs 15, 16 & 22) is also distinct from that of A. amoyensis (Trewavas, 1977: Fig. 9b; Chu et al., 1963: Fig. 82); it is more elongate, with a prominent bulge on posterior sector of ventral margin, rounded (vs straight) posterior margin, greater OCD, more elongate ostium with concave ventral and dorsal margins, and it lacks the protuberance on the posterodorsal corner in A. amoyensis.
Figure 28. Relationship between CPD (caudal peduncle depth) as % CPL (caudal peduncle length) and SL for Argyrosomus inodorus (squares) and A. japonicus (circles).
Based on descriptions of A. heinii by Steindachner (1907) and Mohan, (1984), A. inodorus can be distinguished by lower limb gill-rakers 10-12 (vs 9); dorsal-fin rays 25-29 (vs 32-33) and an S-shaped (vs emarginate) caudal fin in adults.
Figure 30. Relationship between otolith length and TL for Argyrosomus inodorus (squares) and A. japonicus (circles).
Based on the description of A. miiuy by Trewavas (1977), A. inodorus has lower limb gill-rakers 10-12 (vs 9-10), snout length 24-30% HL (vs 22-24% HL), interorbital width 20-25% HL (vs 19-21% HL), upper jaw length 39-44% HL (vs 44-46% HL) and lower jaw length 47-51% HL (vs 53-55% HL). The swim-bladder appendages are much longer in A. miiuy, and at about 27 cm SL, almost completely enwrap the bladder (Trewavas, 1977). The caudal fin is also more elongate in A. miiuy than in A. inodorus, and at 305 mm SL is pointed (Chu et al., 1963) as opposed to S-shaped. Comparisons of the sagitta of A. inodorus (Figs 15, 16 & 22) and A. miiuy (Chu et al., 1963: Fig. 90; Schwarzhans 1993: Figs 239-240) reveal that the otolith of A. miiuy is more slender, with concave to straight (vs straight or convex) dorsal margin, posterodorsal corner more obtuse, posterior margin sharply rounded to pointed (vs rounded), the cauda is less curved, and the postcentral umbo on the lateral surface of the otolith is about 65% of the otolith length (less than 50% in A. inodorus). ETYMOLOGY: Inodorus, Latin for "without smell". A. inodorus was first suspected of being a different species because it lacked the strong brassy/metallic smell that is typical of A. japonicus found in South African waters. REMARKS: For many years A. inodorus has been confused with A. japonicus (as "A. hololepidotus"). However, based on the distribution patterns of the two species (this study), the catch localities, the sizes of the specimens examined and the descriptions of the caudal fin, it is almost certain that all of the authors mentioned in the synonymy (above) were inadvertently referring to A. inodorus (in part or in full.
Figure 32. Relationship between OCD (ostium to cauda distance) as a ratio of OH (otolith height) and TL for Argyrosomus inodorus (squares) and A. japonicus (circles).
A. inodorus occurs from Namibia southwards around the Cape of Good Hope and northwards at least as far as the Kei River (Fig. 7). In the area between the Kei River and Cape Agulhas, A. inodorus rarely enters estuaries or the surf zone, and is caught mostly by skiboat fishermen and trawlers at depths of 10-100 metres (Table 7). However, as one rounds Cape Agulhas and enters the cooler waters of the west coast, the species becomes more abundant in the surf zone. The proportion of A. inodorus in Argyrosomus catches declines in northern Namibia, where water temperatures are higher, and the species is not known in Angola. FISHERIES: A. inodorus is highly regarded as a table fish (mostly sold fresh, but sometimes frozen) and it is an important commercial and recreational species wherever it is found. The majority of the South African catch is made between the Cape of Good Hope and the Kei River. Commercial line-fishermen operating from vessels of between 5 and 15 m in length, catch an average of 835 tons per annum (1988-1992). Although no statistics are available for recreational boat fishermen, their catches are expected to be of a similar magnitude. The South African trawl fishery is responsible for an average by-catch of 217 tons per annum, and Western Cape shore angling and beach seine catches are estimated at 26 and 4 tons respectively (Lamberth et al. 1994).
In Namibia A. inodorus is most abundant north of Meob (200 km south of Walvis Bay). Line-boat fishermen catch an average (1989/93) of 413 tons per annum (unpublished data, Namibian Ministry of Sea Fisheries), and shore anglers land in excess of 40 000 fish (approx. 130 tons) a year (Lenssen et al. 1991). Namibian A. inodorus are restricted to a narrow coastal band of 1-20 m, apparently as a result of an anoxic zone which extends beyond this depth (Hart & Curry, 1960; Boyd, 1983; Bailey et al., 1985).
BIOLOGY: In South Africa A. inodorus attains 50% sexual maturity at 31 cm TL (males) and 34 cm TL (females), with spawning activity observed throughout its distribution, from the Cape of Good Hope to the Kei River Mouth, during spring/summer (Griffiths & Hecht, 1993 [species B]). Nursery grounds consist of soft-bottom bay areas at depths less than 50 m (Wallace et al., 1984; Smale & Badenhorst, 1991; as "A. hololepidotus"). Maximum size recorded was 145 cm TL (36.3 kg). Common to 115 cm TL (15 kg).
Argyrosomus coronus sp. nov.
Argyrosomus hololepidotus (non Lacepede): Trewavas, 1977:327 (in part not figs); Chao & Trewavas, 1981: SCIAEN Argyr 3. Bianchi et al. 1993:164 (in part). Holotype: RUSI 48468, female, 430 mm SL, northern Namibia, 4 km south of Hoarusib River, surf zone, collected by M.H. Griffiths, 15 March 1995.
Paratypes: 18 specimens (147-595 mm) . DIAGNOSIS: A species of Argyrosomus with the following combination of characters: dorsal fin rays 26-30; gill-rakers 4-5 + 8-10; swim-bladder carrot shaped, with 28-38 arborescent appendages; caudal peduncle length 10-12% SL; peduncle depth 76-94% peduncle length; interorbital width 7.5-8.3% SL (24-27% HL); suborbital width 8.210.2% HL; lower jaw length 46-50% HL; midlateral body scale length 2.2-2.7% SL; drumming muscles present in both sexes; urinary bladders large; sagitta typically Argyrosomus (Fig. 11), OCD 26-34% OH; pectoral fins 18-21% SL, not reaching past vertical at tip of pelvic lugs; pectoral fin axillary fold scaleless; peritoneum unpigmented; anterior part of lateral line slightly curved. DESCRIPTION: Based on 19 preserved (147-595 mm), 7 fresh specimens (486-598 mm), one skeleton (495 mm) and 107 otolith pairs (281-1110 mm). Counts and measurements for the holotype are in parentheses. Dorsal fin X+I,26-30 (27); anal fin II,7; pectoral-fin rays 16-18 (18); lateral-line scales 50-53 (51); gill-rakers 4-5 + 8-10; swim-bladder appendages 28-38; vertebrae 11 + 14. Proportions as % SL: head length 31-32 (31); pre-dorsal length 33-36 (35); pre-pelvic length 33-36 (34); body depth 25-28 (27); caudal peduncle length 1 O- 12 ( 12); peduncle depth 8.6-10 (10); snout 8.4-9.3 (9.2); eye diameter 6.7 at 147 mm, 5.2-6.0 at 202-281 mm, 4.7-5.0 at 312-385 mm, 4.2-4.7 (4.5) at 408-471 mm and 3.8-4.4 at 523-595 mm SL; interorbital width 7.5-8.3 (8.2); suborbital width 2.6-3.2 (2.9); upper jaw 12-14 (13); lower jaw 15-16 (15); pectoral fin 18-21 (20); pelvic fin 21 at 147 mm, 19-20 at 202-281 mm, 17-18 at 312-385 mm, 16-18 (17) at 408-471 mm and 15-18 at 523-595 mm SL; anal fin length 18 at 147 mm, 15-17 at 202-281 mm, 15-16 at 312-385 mm, 14-15 (15) at 408-471 mm and 13-14 at 523-595 mm SL; third dorsal spine 13-15 at 202-281 mm, 11-13 at 312-385 mm, 11-13 (13) at 408-471 mm and 11 at 595 mm SL; second anal spine 9.7 at 147 mm, 7.5-8.7 at 202-281 mm, 6.4-8.3 at 312-385 mm, 6.8-8.3 (7.2) at 408-471 mm and 5 at 595 mm SL; gill-raker 1.6-2.3; gill-filament 3.0-3.8.
Proportions as % HL: snout 27-31 (30); eye diameter 22 at 147 mm, 17-19 at 202-281 mm, 15-16 at 312-385 mm, 14-15 (14) at 408-471 mm and 14 at 523-595 mm SL; interorbital width 24-27 (27); suborbital width 8.2-10.4 (9.2); upper jaw 38-43 (43); lower jaw 46-50 (49); gill-raker 5.0-7.4; gill-filament 9.9-12.1.
Proportions as % eye diameter: gill-raker 31-53; gill-filament 56 at 147 mm, 60-62 at 202-281 mm, 65-73 at 312-385 mm, 67-78 at 408-471 mm and 83 at 523-595 mm SL. Dorsal profile of A. coronus includes a hump or "shoulder" behind the head. Pectoral fins not reaching past vertical at tip of pelvic fins (for fish 147-595 mm). Nostrils as for A. japonicus. Caudal fin mostly S-shaped (281-595 mm), but sometimes wedge shaped, bi-concave or rounded. Lateral line evenly curved, axillary skin fold at base of pectoral fin scaleless. Scales cycloid below eye, but ctenoid on body (including belly and chest), nape of neck, interorbital region, and operculum. Gill-rakers at angle shorter than outer gill-filaments at angle (281-593 mm). Drumming muscles well developed in both sexes. Pyloric caeca 8-10. Urinary bladders large, as in A. japonicus.
Figure 34. Relationship between SOCCRL (supraoccipital crest length) and SL for Argyrosomus inodorus (squares), A. japonicus (circles), A. coronus (triangles) and A. thorpei (crosses). Figure 36. Swim-bladder of Argyrosomus coronus, 360 mm SL: A) lateral view and B) ventral (left half) views.
Otoliths (Fig. 10): Description as for A. japonicus, but cauda more strongly curved (less angular) and posterior margin of cauda more rounded; OCD 26-34% OH.
Neurocranium (Figs 4 & 5) and skeleton: Length of neurocranium (NCL) 21% SL; pre-supraoccipital crest length 11% SL and 52% NCL; supraoccipital crest length 9.1% SL and 44% NCL (495 mm) ; neurocranial Width 8.4% SL and 41% NCL. Interorbital septum relatively well ossified with frontal fused with medial ethmoid and parasphenoid bones. Premaxilla 7.7% SL and 37% NCL; dentary 9.2% SL and 44% NCL. Third and fourth neural spines 8.9 and 8.3% SL.
Colour: In life, silvery grey/brown becoming darker above and lighter below; dorsal surface with a bluish bronze sheen that may become coppery on head. Some individuals darker brown than others; fins dark grey/brown; mouth lining yellow, some with a touch of grey; inside of operculum yellow and dark grey. Axillary fold of pectoral fin grey to black, generally with an irregular coppery-bronze patch.
Preserved specimens vary, usually grey, darker above and lighter below. Dark patches on scales may give appearance of oblique stripes. Fins grey, sometimes with one or two longitudinal stripes on dorsal fin. COMPARISONS (Tables 2-6): A. coronus generally has a more pointed head (resulting from a less convex snout, Fig. 35) than the other species described in this paper, including A. regius (Figs 9, 12, 19 & 21).
A. coronus differs from A. regius in having a higher modal number of lower limb gill-rakers; a slightly longer lower jaw; longer midlateral scales (Fig. 37); a shorter and deeper caudal peduncle (Fig. 38); fewer arborescent swim-bladder appendages, in which the lower limb is orientated latero-anteriorly (vs ventro-anteriorly) with branches that project at 90 degrees to the bladder (vs project posteriorly and lie against the bladder). The otoliths of A. coronus are more elongate (Figs 10 & 39) than those of A. regius; the cauda is more curved, with a more rounded posterior margin (Fig. 40); and the curvature of the posterior margin of the ostium more acute.
Figure 38. Relationship between CPD (caudal peduncle depth) as % CPL (caudal peduncle length) and SL for Argyrosomus regius (circles) and A. coronus (squares). Figure 39. Relationship between otolith length and SL for Argyrosomus regius (circles) and A. coronus (squares). Figure 40. Superimposed sagittae of Argyrosomus regius, 516 mm TL (solid line) and A. coronus, 512 mm TL, (broken line).
A. coronus differs from A. hololepidotus in having a wider interorbital (Fig. 18), a narrower suborbital, and a shorter and deeper caudal peduncle. The sagitta of A. coronus (Fig. 10) has a more strongly curved cauda, which is less angular, and has a more rounded posterior margin than that of A. hololepidotus (Fig. 15).
Differences between A. coronus and A. thorpei include fewer lower limb gill-rakers, a higher modal number of pectoral fin rays and swim-bladder appendages, a wider interorbital, a narrower suborbital, a naked (vs scaled) pectoral axillary fold, a shorter caudal peduncle, shorter midlateral body scales, and a slight (vs strong) anterior curve of the lateral line. The sagitta of A. coronus is not expanded in the anterior two-thirds, it has a more acute posterodorsal angle, a cauda which is neither expanded in the distal portion nor truncated at the tip, and a shorter OCD (Figs 11 & 15).
A. coronus differs from A. inodorus in having lower modal numbers of upper and lower gill-rakers, a lower modal number of swim-bladder appendages, a wider interorbital (Fig. 41 and Table 2), a shorter and deeper caudal peduncle (Fig. 42 and Table 2), a smaller orbital diameter (Fig. 43), larger urinary bladders, drumming muscles in both sexes (vs in males only), a longer supraoccipital crest (Fig. 34 and Table 2), a smaller interorbital window (frontal and parasphenoid bones connected, Fig. 4) and shorter premaxilia (7.7 vs 8.1-9.1% SL) and dentary (9.2 vs 9.3-10.2% SL). The sagitta of A. coronus (Fig. 10) has a more acute posterodorsal angle, a smaller OCD (Fig. 44), a lower OCD:OH ratio (Fig. 45), a straight (vs rounded) posterior margin, and. it lacks the prominent bulge found on the ventral margin in A. inodorus (Fig. 15). A. coronus is generally darker and more brown in colour (body) than A. inodorus, and the fins are dark grey/brown (vs yellow/grey).
Figure 42. Relationship between CPD (caudal peduncle depth) as % CPL (caudal peduncle length) and SL for Argyrosomus coronus (circles) and A. inodorus (squares).
Figure 43. Relationship between orbital diameter and HL for Argyrosomus coronus (circles) and A. inodorus (squares). Figure 44. Relationship between OCD (ostium to cauda distance) and TL for Argyrosomus coronus (circles) and Namibian A. inodorus (squares) Figure 45. Relationship between OCD (ostum to cauda distance) as a ratio of OH (otolith height) and TL for Argyrosomus coronas (circles) and Namibian A. inodorus (squares).
A. coronus differs from A. beccus Sasaki (1994) in having swim-bladder appendages 28-38 (vs 23); longer snout (8.4-9.3 % vs 6.1-6.3 % SL), snout convex (concave in A. beccus); eye diameter 5.2-6.7% SL (vs 7.3-8.3% SL, for fish of comparable size); wider interorbital 7.5-8.3% SL (vs 4.7% SL), and pelvic fin length 19-21% SL (vs 22-23% SL, for fish of comparable length). The sagitta of A. coronus (Fig. 10) is distinguished from that of A. beccus (Sasaki, 1994: Fig. 2B); in that it is more elongate: with a longer ostium, a more strongly curved cauda, and the postcentral umbo on the lateral surface of the otolith is less than 50% of the otolith length (about 75% in A. beccus).
Based on descriptions of A. heinii by Steindachner (1907) and Mohan (1984), A. coronus can be distinguished by dorsal-fin rays 26-30 (vs 32-33) and an S-shaped (vs emarginate) caudal fin in medium sized fish.
Based on the description of A. miiuy by Trewavas (1977), A. coronus has snout length 27-31% HL (vs 22-24% HL), interorbital width 24-27% HL (vs 19-21% HL), upper jaw length 38-43% HL (vs 44-46% HL) and lower jaw length 46-50% HL (vs 53-55% HL). The swim-bladder appendages are much longer in A. miiuy, and at about 27 cm SL, almost completely enwrap the bladder (Trewavas, 1977). The caudal fin is also more elongate in A. miiuy than in A. coronus, and at 305 mm SL is pointed (Chu et al., 1963) as opposed to S-shaped. Comparisons of the sagitta of A. coronus (Fig. 10) and A. miiuy (Chu et al., 1963: Fig. 90; Schwarzhans, 1993: Figs 239-240) reveal that the otolith of A. miiuy is more slender, with concave or straight (vs straight or convex) dorsal margin, sharply rounded or pointed (vs straight) posterior margin, posterodorsal corner more obtuse, cauda less curved, and the postcentral umbo on the lateral surface of the otolith about 65% of the otolith length (less than 50% in A. coronus). ETYMOLOGY: This species is named Argyrosomus coronus in honour of Corona Griffiths, wife of the first author, in appreciation of her contributions to the illustrations of this paper and her help with field work. The specific name is treated as a noun in apposition. REMARKS: Although morphometric differences between A. coronus and A. regius are subtle, and possibly would not justify species recognition, we have placed much weight on swim-bladder appendage counts, and on differences in otolith morphology for the following reasons: Swim-bladders of A. regius specimens from the Mediterranean, French Atlantic and north Africa provided no evidence for a cline in appendage counts. Counts of A. coronus specimens from Namibia and Angola were also similar. In addition the numbers of appendages of other Argyrosomus species were found to be consistent over wide distributional ranges e.g. A. japonicus from South Africa, Australia and Japan; and A. inodorus from the eastern seaboard of South Africa and from central Namibia. Differences in the morphology of the otoliths of specimens from the northern and southern hemispheres were also regarded as important because these structures often do not differ between recognized Argyrosomus species e.g. A. regius, A. japonicus and A. hololepidotus.
The recognition of A. coronus as a separate species also assists with species identification in Namibia. This species has a shorter and deeper caudal peduncle and a lower number of swim-bladder appendages than A. regius, and together with interorbital width is therefore more easily separated from A. inodorus. DISTRIBUTION: Known from Namibia and Angola. It becomes more abundant in Argyrosomus catches as one moves northwards (Table 7) indicating that it prefers warmer waters. Found in estuaries, the surf zone and further offshore. Reports of "A. hololepidotus" from the west coast of Africa, south of the equator (Trewavas, 1977; Mohan, 1984; Heemstra, 1986), are probably based on specimens of A. coronus. FISHERIES: In Namibia A. coronus is caught by commercial and recreational fishermen in fairly low numbers. Areas further north are closed to the public. In southern Angola it is caught with hand lines, gillnets, trapnets and occasionally purse seines; mostly in depths of 20-40 m, but also in the surf zone and in depths to 100 m (B.Bennett pers comm). BIOLOGY: The biology of this species has not been studied. Personnel of the Namibian Ministry of Sea Fisheries found fish up to 13 kg to be immature, suggesting a large size at maturity, similar to that of A. japonicus and A. regius. Adults are rarely found in Namibia but according to Bruce Bennett (pers comm) are abundant in southern Angola (common to 50 kg). Maximum size 77 kg. The lack of permanent estuaries in southern Angola suggests that this habitat type does not play an important role in the life history of A. coronus.
Table 1. Regression equations for data from Argyrosomus japonicus (J) and A. inodorus (I) from South African waters for weight/total length (W/TL), otolith dimension/TL, and standard length/total length (SL/TL) relationships. OL = otolith length, OH = otolith height, OC = ostium to cauda distance and OW = otolith weight. The asterisk (*) denotes a significant difference (P< 0.01) between the slopes and/or intercepts of the corresponding relationships, after log-transformation.
---------------------------------------------------------------------------
Relationship r n Range (TL) F-ratio P
---------------------------------------------------------------------------
J W(g)=8.391x10^-6TL(mm)^3.025 0.999 1215 37-1750 mm 29.6 <0.0001*
I W(g)=6.832x10^-6TL(mm)^3.055 0.997 3146 60-1378 mm
J OL(mm)=TL(mm)^0.706/5.678 0.995 543 37-1750 mm 1238.3 <0.0001*
I OL(mm)=TL(mm)^0.699/4.691 0.993 623 60-1310 mm
J OH(mm)=TL(mm)^0.630/5.931 0.989 542 37-1750 mm 19.0 <0.0001*
I OH(mm)=TL(mm)^0.603/4.932 0.990 623 60-1310 mm
J OC(mm)=TL(mm)^0.589/14.821 0.969 539 50-1750 mm 3251.2 <0.0001*
I OC(mm)=TL(mm)^0.607/10.717 0.972 616 60-1310 mm
J OW(g)=TL(mm)^2.008/479004 0.989 542 37-1750 mm 138.6 <0.0001*
I OW(g)=TL(mm)^1.896/195344 0.989 623 60-1310 mm
J SL=0.877 TL - 10.882 0.999 167 35-1790 mm - -
I SL=0.886 TL - 14.882 0.999 408 140-970 mm - -
---------------------------------------------------------------------------
Table 2. Diagnostic characters of Argyrosomus species. CPD = caudal peduncle depth, CPL = caudal peduncle length, F = female, M = male, OCD = ostium-cauda distance, OH = otolith height, Argyr. = as for A. regius.
---------------------------------------------------------------------------
hololepidotus inodorus japonicus regius coronus thorpei
---------------------------------------------------------------------------
Gill-rakers 9 10-12 9-11 8-10 8-10 10-11
(lower limb)
Swim-bladder 34-36 3241 21-31 36-45 28-38 25-33
appendages
Interorbital width 22-23 20-2.5 21-24 24-29 24-27 20-21
Suborbital width 10.5-12 8.6-10.9 8.4-10.5 8.5-11 8.2-10.2 10-11.5
(% HL)
Lower jaw length 50 46-51 48-52 4448 46-50 49-53
(% HL)
CPD as %CP 75-77 58-74 70-92 68-82 76-94 65-79
CPL as %SL 12-13 12-15 11-13 11-13 11-12 12-14
Pectoral fin length 17-18 19-23 17-21 18-21 17-21 20-23
(%SL)
Midlateral body 2.5-2.8 1.8-2.5 2.4-3.1 1.8-2.3 2.2-2.7 2.6-3.3
scale - Length (%SL)
Supraoccipital crest - 8.0-8.8 9.2-10.2 - 9.1 -
length (%SL)
Urinary bladder large 0.1-3.3 2.0-9.6 large large large
length (%TL)
OCD as %OH 28-34 37-63 2341 26-32 34-44
Otolith morphology Argyr. distinct Argyr. Argyr. Argyr. distinct
Anterior curve of slight slight slight slight slight strong
lateral line
Pectoral fin scaleless scaleless scaleless scaleless scaleless scaly
axillary fold
Drumming muscles M & F M only M & F M & F M & F M & F
---------------------------------------------------------------------------
Table 3. Frequency distributions for swim-bladder appendage counts in Argyrosomus hololepidotus (H), A. inodorus (I), A. japonicus (J), A. coronus (C), A. regius (R), and A. thorpei (T).
--------------------------------------------------------------------------
Swim-bladder appendages
n x 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
--------------------------------------------------------------------------
H 3 35 1 1 1
I 77 35.8 1 3 9 9 15 8
J 63 26.3 1 2 6 6 12 5 15 5 4 6 1
C 24 39.9 3 4 2 1 4 4 2 1
R 23 39.3 3
T 18 29.4 1 1 3 3 4 4 2
---------------------------------------------------------------------------
Swim-bladder appendages
-----------------------------------------------------------
n x 37 38 39 40 41 42 43 44 45
H 3 35
I 77 35.8 10 7 ? 4 3 2
J 63 26.3
C 24 39.9 1 1
R 23 39.3 4 2 1 5 3 2 1 1 1
T 18 29.4
------------------------------------------------------------Table 4. Frequency distributions for meristic characters in Argyrosomus hololepidotus (H), A. inodorus (I), A. japonicus (J), A. coronus (C), A. regius (R) and A. thorpei (T).
---------------------------------------------------------------------------
Upper Gill-rakers Lower Gill-rakers
-------------------------------- ---------------------------------------
n x 3 4 5 6 n x 8 9 10 11 12
------------------ -----------------------
H 3 4.7 1 2 H 3 9 3
I 76 5.2 1 59 16 I 76 11 17 48 11
J 66 4.6 24 42 J 66 9.9 11 51 13
C 25 4.1 23 2 C 25 9.5 1 11 12 1
R 17 4.3 1 10 6 R 19 9.1 1 16 2
T 18 5.1 1 15 2 T 18 11.1 1 15 2
---------------------------------------------------------------------------
n x 15 16 17 18 n x 25 26 27 28 29 30
------------------ --------------------------
H 2 16.5 1 1 H 3 27.3 2 1
I 393 16.5 11 193 185 4 I 381 27.1 10 85 160 99 26 1
J 68 16.1 3 50 12 J 61 26.7 4 28 23 2 2
C 23 16.7 1 5 16 1 C 24 27.2 5 12 5 1 1
R 19 16.9 5 11 3 R 18 27.6 1 1 6 6 3 1
T 18 16 3 12 3 T 18 27.1 3 11 3 1
---------------------------------------------------------------------------
Table 5. ANOVA: Multiple range tests of meristic characters that show significant differences (P< 0.0001). Mean values increase from left to right; * indicates a significant difference at the 1% level and # a difference at the 5% level. Argyrosomus inodorus (I), A. japonicus (J), A. regius (R), and A. thorpei (T).
---------------------------------------------------
Character Species
---------------------------------------------------
Pectoral fin-rays T J * I # R
-----
Gill-rakers (upper) R * J * T I
------
Gill-rakers (lower) R * J * I T
------
Swim-bladder appendages J * T * I * R
---------------------------------------------------Table 6. ANOVA: Multiple range tests of arc-sine transformed morphometric ratios that show significant differences (P< 0.0001). Mean values increase from left to right; * signifies a difference significant at the 1% level and # a difference at the 5% level. Argyrosomus inodorus (I), A. japonicus (J), A. regius (R), and A. thorpei (T).
------------------------------------------------------
Character Species
------------------------------------------------------
Suborbital width J * I R * T
------
Interorbital width J T I * R
-----------
Pre-dorsal length I * T J R
------------
Body depth I R * J # T
Caudal peduncle length J # R * T I
-----
Caudal peduncle depth I R * T J
----- -----
Scale height R * I * J * T
Scale length R I * J T
----- -----
Gill-raker length R J I * T
----------
-----------------------------------------------------Table 7. The distribution of South African Argyrosomus species, based on relative contributions (% by number) to catches made in four habitat types of 10 regions (see Fig. 7 for regional boundaries). A. inodorus (I), A. japonicus (J), A. coronus (C), A. thorpei (T).
---------------------------------------------------------------------------
Estuary Surf zone Nearshore reef Nearshore sand/mud
---------------------------------------------------------------------------
S. Mozambique - - 99.8%T:O.2%J -
(n=1021)
N. Natal 100%J 35%T:65%J 95%T:5%J 100%T
(n=45) (n=32) (n=557955) (n=832)
S. Natal 100%J 100%J 85%T:15%J -
(n=13) (n=36) (n=87456)
Transkei 100%J 100%J - -
(n=410) (n=16)
E. Cape 100%J 98%J:2%I 1.3%J:98.7%I 100%I
(n=670) (n=590) (n=2629) (n=1050)S.
S. Cape 100%J 100%J 1.5%J:98.5%I 100%I
(n=168) (n=SS) (n=22128) (n=1473)
S.W. Cape Rare 3.3%J:96.7%I 100%I 100%I
(n=333) (n=558) (n=33)
W.Cape Rare 100%I 100%I Rare
(n=9) (n=42)
S. Namibia no 100%I - -
estuaries (n=229)
C. Namibia no 93%I:7%C 100%I 100%I
estuaries (n=740) (n=225) (n=52)
N. Namibia no 50%I:50%C 100%I -
estuaries (n=408) (n=220)
S. Angola - 100%C 100%C 100%C
(n=30) (n=10) (n=6)
--------------------------------------------------------------------------
Table 8. Arcsine-transformed morphometric ratios for A. inodorus and A. japonicus that showed significant difference (ANOVA) only when analysed according to size class. 1 = 100-199 (n= 7+10), 2 = 200-299 (n= 28+13), 3 = 300-399 (n= 21+13), 4 = 400-499 (n= 6+11), 5 = 500-599 (n= 4+5) * = 5% level (p<0.05), ** = 1% level (p<0.01). Size class ranges are of mm SL.
-------------------------------------------
Character Size classes (100 mm)
------------------------
1 2 3 4 5
Eye diameter * ** ** * *
Pectoral fin length ** * *
Anal fin length ** **
-------------------------------------------ACKNOWLEDGEMENTS We are grateful to: Dr Bernard Seret for donating specimens of Argyrosomus regius; to Dr Yukio Iwatsaki for donating specimens of A. japonicus from Japan; and Dr Johan Bell and Mr Mat Broadhurst who donated whole specimens and otoliths of this species from Australia. The South African Rock & Surf Anglers Union contributed 14 large specimens of A. japonicus from the eastern Cape. Many of the whole A. inodorus specimens were collected during research cruises aboard the RV AFRICANA of the Department of Environmental Affairs and Tourism. Otoliths were provided by Barry Clarke, Dr Bruce Bennett, Steven Lamberth, Dr Theresa Lasiak, Prof Hannes Marais, Dr Emil Plumstead, Koos Venter, Jurg Walters, the Oceanographic Research Institute (Durban) and the Linefish Section of the Namibian Ministry of Fisheries and Marine Resources. Specimens were loaned by the British Museum (Dr Nigel Merret), Museum National d' Histoire Natarelle, Paris (Dr Marie-Louise Bauchot, Guy Duhamel & Bernard Seret), Institut Royale des Sciences Naturales de Belgique (Dr J.-P. Gosse), the Australian Museum (Mr Mark McGrouther), and the Hebrew University of Jerusalem (Dr D. Golani). Mrs S. Radloff from the Department of Mathematics and Statistics at Rhodes University, Grahamstown, assisted with the statistical analyses. Ms Joan Wright and Marion Baxter produced camera-ready copy of the text. Mr Robin Stobbs of the JLB Smith Institute of Ichthyology in Grahamstown, radiographed specimens. The illustrations were done by Marc Griffiths and Tony Van Dalsen (otoliths), Corona Griffiths (skulls, jaws & swim-bladders) and Elaine Heemstra (whole specimens). We would also like to thank Drs Humphry Greenwood, Kunio Sasaki, Eric Anderson and John Paxton for constructive comments on the manuscript. This work was partially funded by the Sea Fisheries Fund of South Africa. LITERATURE CITED ASSO, L DE. 1801. Introduction a la ichthyologia oriental de Espania. An. Cienc. nat. Madrid. 4: 28-52. BAILEY, G.W., BEYERS, C.J. DE B. & S.R. LIPSCHITZ. 1985. Seasonal variation of oxygen deficiency in waters off southern South West Africa in 1975 and 1976 and its relation to the catchability and distribution of the Cape rock lobster Jasus lalandii. S. Afr. J. Mar. Sci. 3:197-214 BALGUERIAS, E. 1985. Actividad de la flota artesanal canaria que faeno en la costa noroccidental africana de 1975 a 1982. (pp 851-871) In: C. Bas, R. Margalef & P. Rubies (eds), Simposia internacional sobre las areas de afloramiento mas importantes del oeste africano (Cabo Blanco Y Benguela). vol II. Instituto de Investigaciones Pesqueras, Barcelona. BARNARD, K.H. 1927. A Monograph of the Marine Fishes of South Africa. Part 2. Neill and Co., Ltd., Edinburgh. pp 419-1065. BAUCHOT, M.-L. & G. BIANCHI. 1984. Fiches FAO d'identification des especes pour les besoins de la peche. Guide des poissons commerciaux de Madagascar (especes marines et d'eaux saumatres). Avec le support du Programme des Nations Unies pour le Development (Project RAF/79/065). Rome, FAO, 135 pp. BIANCHI, G.K.F. CARPENTER, J.P. ROUX, F.J. MALLOY, D. BOYER, & H.J. BOYER, H.J. 1993. FAO species identification field guide for fishery purposes. The living marine resources of Namibia. Rome, FAO, 250 pp. BLEEKER, P. 1860. Over enige vischsoorten van de Kaap de Goede Hoop. Natuur. Tijdschr. Ned.-Indie. 21: 49-80. --- 1863. Descriptions des quelues especes de poissons nouvelles ou peu connues de Chine envoyees au Musee de Leide par M. G. Schlegel. Ned. Tijdschr. Dierk. 1: 135-150. --- 1876. Systema Percarum Revisum. Archs neerl. Sci. Nat. 11: 247 -340. BOESEMAN, M. 1947. Revision of the fishes collected by Burger and von Sicbold in Japan. Zool. Meded. Leiden, 28: 1-242. BOYD, A.J. 1983. Intensive study of the currents winds and hydrology at a coastal site off central South West Africa, June/July 1978. Invest. Rep. Sea Fish. Res. Inst. S. Afr. 126: 37pp BRANCH, G.M., C.L. GRIFFITHS, M.L. BRANCH, & L.E. BECKLEY. 1994. Two Oceans, a Guide to the Marine Life of Southern Africa. David Philip, Cape Town. 360 pp. BRONNICH, M.T., 1768. Ichthyologia Massiliensis, sistens piscium descriptiones eorumque apud incolas nomina. Accedunt Spolia Maris Adriatici. Hafniae et Lipsiae: xvi+110 pp. CASTELNAU, F.L. de L. 1872. Contribution to the ichthyology of Australia. Proc. zool. acclm. Soc. Vici. 1: 29-247. --- 1878. Essay on the ichthyology of Port Jackson. Proc. Linn. Soc. N.S.W. 3: 347-402. CHAINE, J. 1938. Recherches sur les otolithes des poissons. Etude descriptive et comparative de la sagitta des Teleosteens (suite). Act. Soc. finn. Bordeaux. 90: 5-258. CHAMPAGNAT, C. & F. DOMAIN. 1978. Migrations des poissons demersaux le long des cotes Ouest-Africaines de 10 a 240 de latitude nord. Cah. O.R.S.T.O.M. ser. Ocdanogr. 16(3-4): 239-261. C HAO, L.N. 1986. Sciaenidae. (pp 865-874)In: P.J.P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielson & E. Tortonese (eds), Fishes of the North-eastern Atlantic and the Mediterranean, vol. 2: 515-1007. UNESCO, Paris. CHAO, L.N. & E. TREWAVAS. 1981. Sciaenidae. (40 pp) In: W. Fischer, G. Bianchi & W.B. Scoa (eds). FAO species identification sheets for fishery purposes. Eastern Central Atlantic; fishing areas 34, 47 (in part). Ottawa, Department of Fisheries and Oceans Canada by arrangment with the Food and Agriculture Organization of the United Nations, vol. 3: pag. var. CHU, K.Y. 1956. A review of the sciaenoid fishes of Taiwan. Rep. Inst. Fish. Taipeh. 1: 13-47. CHU, Y.T., Y.L. LO & H.L. WU. 1963. A study on the classification of sciaenoid fishes of China, with description of new genera and species. Science and Technology Press of Shanghai. 100 pp.(In Chinese with English summary). CUVIER, G. 1814. Sur un poisson celebre, et cependant presque inconnu des auteurs systematiques, appele sur nos cotes de l'Ocean, Aigle ou Maigre, et sur celles de la Mediterranee, Umbra, Fegaro et Poisson Royal; avec une description abrebgee de sa vessie natatoire. Mere. Mus. Hist. nat. Paris. 1:1-21 (for date see Sherborn 1914, in Trewavas 1977). --- 1816. Le Regne Animal. 4 vol. Paris. Poissons, 2: 104-351. CUVIER, G. & A. VALENCIENNES. 1830. Histoire Naturelie des Poissons. F.G. Levrault, Paris. 6:374 pp. DAY, F. 1880-84. The Fishes of Great Britain and Ireland. Vol.I. Williams & Norgate, London: 336 pp.+92 Pls. DE VIS, C.W. 1884. New fishes in the Queensland Museum No. 3. Proc. finn. Soc. N.S.W. 9: 537-547. FENNESSY, S .T. 1994. The impact of commercial prawn trawlers on line fish catches on the north coast of Natal. S. Afr. J. Mar. Sci. 14: 263-279. FISCHER, W. & P.J.P. WHITHEAD (eds). 1974. FAO species identification sheets for fishery purposes, Eastern Indian Ocean (fishing area 57) and Western Central Pacific (fishing area 71). Rome: F.A.O. 1-4. FOWLER, H.W. 1936. The marine fishes of West Africa. Bull. Am. Mus. nat. Hist. 70: 1-1493. GOMON, M.F. 1994. Family Sciaenidae. (pp 602-603) In: M.F. Gomon, C.J.M. Glover & R.H. Kuiter (eds), The Fishes of Australia's South Coast. State Printer, Adelaide, 992 pp. GRANT, E.M. 1972. Guide to Fishes. Queensl. Dep. Primary Ind., Brisbane, (2nd edn). 472 pp. --- 1978. Grant's Guide to Fishes. Department of Harbours and Marine, Brisbane. 767 pp. --- 1987. Fishes of Australia. E.M. Grant PTY Limited, Queensland. 480 pp. GRIFFITHS, M.H. & T. HECHT. 1993. Two South African Argyrosomus hololepidotus species: implications for management. In: L.E. Beckly & R.P. van der Elst, (eds), Fish, fishers and fisheries-Proceedings of the second South African marine linefish symposium. Oceanographic Research Institute, Special Publication (2): 19-22. HALL, D.A. 1986. An assessment of the mulloway (Argyrosomus hololepidotus) fishery in Australia with particular reference to the Coorong lagoon. South Australian Fisheries Discussion Paper. HALY, A. 1875. Description of new species of fish in the collection of the British Museum. Ann. Mag. nat. Hist. 15(4): 268-270. HART, T.J. & R.I. CURRY. 1960. The Benguela Current. "Discovery" Rep. 30: 123-297. HEEMSTRA, P.C. 1986. Family Sciaenidae. (pp 616-619) In: M.M. Smith & P.C. Heemstra, (eds), Smiths' Sea Fishes. Macmillan Publishers, Johannesburg. 1047 pp. HUBBS, C.L. & K.F. LAGLER. 1964. Fishes of the Great Lakes Region. Univ. Michigan Press, Ann Arbor. 213 pp. JONES, K. 1991. Fin fishery considerations: In the management of the proposed Great Australian Bight National Park. Safish. 15(4): 11-12,15. KAILOLA, P.J., M.J. WILLIAMS, P.C. STEWART, R.E. REICHELT, A. MCNEE & C. GRIEVE. 1993. Australian Fisheries Resources. Bureau of Resource Sciences, Department of Primary Industries and Energy, and the Fisheries Research and Development Corporation. Canberra. 422 pp. KAMOHARA, T. 1964. Revised catalogue of fishes of Kochi Prefecture, Japan. Rep. Usa Mar. biol. Stn. 11:1-99. KUITER, R.H. 1993. Coastal Fishes of South-eastern Australia. Crawford House Press, Bathurst. 437 pp. LACEPEDE, B .G.E. 1801. Histoire naturelie des Poissons. Paris. 3:558 pp. --- 1803. Histoire naturelie des Poissons. Paris. 4:728 pp. LAMBERTH, S.J., B.A. BENNETT & B, CLARK. 1994. Catch composition of the commercial beach seine fishery in False Bay, South Africa. S. Afr. J. Mar. Sct. 14: 69-78. LENSDA, J., P. TARR & H. BERRY. 1991. An assessment of visitor statistics and line fishing along the Sandwich shoreline, Namib-Naukluft Park, Namibia. Madoqua 18(1): 33-36. LEVITON, A.E., R.H. GIBBS Jr., E. HEAL & C.E. DAWSON. 1985. Standards in herpetology and ichthyology: Part I. Standard symbolic codes for institutional resource collections in herpetology and Ichthyology. Copeia (3): 802-832. LIN, S.Y. 1938. Further notes on the sciaenid fishes of China. Lingnan Sci. J. 17: 161-173. --- 1940. Croakers of the South China Sea. J. Hong Kong Fish. Res. Stn 1: 243-254. MACLEAY, W.S. 1881. Descriptive catalogue of the fishes of Australia. Partii. Proc. Linn. Soc. N.S.W. 5: 510-629. MATSUBARA, K. 1937. Sciaenoid fishes found in Japan and adjacent waters. J. Imp. Fish. Inst. Tokyo. 32: 27-92. McCULLOCH, A.R. 1922. The fishes of New South Wales. Roy. Zool. Soc. of New South Wales, Sydney. 104 pp. + 43 Pls. MOAL, A.R. 1957. Biology of economically important species "courbine" or "Meagre". C.C,T.A./ A.C.S.A. Sea Fisheries 1. Africa (57) O.C.W. 3 1957.30 pp. (mimeogr.). MOHAN, R.S. LAL. 1984. Sciaenidae. 34 pp. In: W. Fischer & G. Bianchi (eds). FAO species identification sheets for fishery purposes, Western Indian Ocean (Fishing Area 51). vol. 4: Food and Agriculture Organization of the United Nations, Rome. MYERS, R.H. 1990. Classical and Modern Regression with Applications. P.W.S. Kent, Boston. (2 edn). 488 pp. OGILBY, J.D. 1918. Edible fishes of Queensland. Parts X-XIV. Mem. Qd Mus. 6: 45-90. OGILBY, J.D. & T.C. MARSHALL. 1954. The Commercial Fish and Fisheries of Queensland. Department of Harbours & Marine, Brisbane. 121 pp. OKAMURA, O. 1988. Sciaenidae. pp. 161-163. In: H. Masuda, K. Amaoka, C. Azaga, T. Uyeno & T. Yoshino (eds). Fishes of the Japanese Archipelago. Tokai University Press, Tokyo (2 edn). 456 pp.+ 378 Pls. OLIVER, G. & A. LAFON. 1981. Variations saisonnieres des Sciaenidae (Pisces, Teleostei, Perciformes) dans le Golfe de Gascogne (Atlantic Oriental). Cybium (serie 3e). 5(3): 35-43. PAPPE, L. 1866. Synopsis of the Edible Fishes of the Cape of Good Hope. W. Brittain, Cape Town.(2d edn). 23 pp. PELLEGRIN, J. 1914. Sur une collection de poissons de Madagascar. Bull. Soc. zool. Fr. 39: 221-234. POLL, M. 1954. Poissons. IV. Teleostoens Acanthopterygiens Sud. (1948-1949). Inst. R. Sci. Nat. Beige. 4(3A): 1-390.( Premiere partie). Rs. Sci. Exped. oceanogr. belg. Eaux. cot. PYLAIE, DE LA. 1835. Recherches, en France, dur les poissons de l'Ocean, pendant les annees 1832 et 1833. C.R. Congres Sci. Fri. Poitiers. 1835: 532. QUERO, J.-C. 1989. Sur lapiste des maigres Argyrosomus regius (Pisces, Siaenidae) du Golfe de Gascogne et de Mauritanie. In: Facteurs externes et internes du determinisme des migrations: seminaire 15 et 22 Decembre 1988. Oceanis Doc. Oceanogr. 15(2): 161-170. QUERO, J.-C. & J.-J. VAYNE. 1987. Le maigre, Argyrosomus regius (Asso 1801)(Pisces, Perciformes, Sciaenidae) du Golfe de Gascogne et des eaux plus septentrionales. Rev. Trav. Peches marit. 49(1 et 2): 35-66. RAMSEY, E.P. & J.D. OGILBY. 1886. On an undescribed Sciaena from the New South Wales coast. Proc. finn. Soc. N.S.W. 11: 941. ROUGHLEY, T.C. 1916. Fishes of Australia and their Technology. New South Wales Department of Education, Sydney. 296 pp. --- 1951. Fish and Fisheries of Australia. Angus & Robertson, Sydney. 343 pp. RISSO, A. 1810. Ichthyology de Nice. Paris. 388 pp. + 11 pls --- 1826. Histoire Naturelie des Principales Productions de l'Europe Meridionale. Paris. 3:486 pp. + 16 pls. SAINSBURY, K.J., P.J. KAILOLA & G.G. LEYLAND. 1984. Continental Shell Fishes of Northern and North-western Australia. Clouston & Hall and Peter Pownall Fisheries Information Service, Canberra. 375 pp. SASAKI, K. 1989. Phylogeny of the family Sciaenidae, with notes on its zoogeography (Teleostei, Perciformes). Mem. Fac. Fish. Hokkaido Univ. 36(1/2): 1-137. --- 1994. Argyrosomus beccus, a new sciaenid from South Africa. Japan. J. Ichthyol., 41(1): 35-38. SASAKI, K. & P.J. KAILOLA. 1988. Three new Indo-Australian species of the sciaenid genus Atrobucca, with a reevaluation of the generic limit. Japan. J. Ichthyol., 35(3): 261-277. SCHWARZHANS, W. 1993. A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). Piscium Catalogus: Part Otolithi piscium. 1: 1-245. SERET, B. 1981. Poissons de Mer de l'ouest Africain Tropical. ORSTOM, Paris. 416 pp. SHERBORN, C.D. & F.A. JENTINK. 1895. On the dates of the parts of Siebald's 'Fauna Japonica'and Giebel's 'Algerniene Zoologie' (1st edn). Proc. zool. Soc. Land. 1895: 149-150. SMALE, M.J. & A. BADENHORST. 1991. The distribution and abundance of secondary trawl fish on the Cape south coast of South Africa, 1986-1990. S. Afr. J. Mar. Sci. 11: 395-407. SMITH, A. 1849. Illustrations of the Zoology of South Africa (Pisces). Smith, EIder & Co, London. 75 pp. SMITH, J.L.B. 1949. The Sea Fishes of Southern Africa. Central News Agency, Cape Town, South Africa. 550 pp. + 111 pls SMITH, J.L.B. & M.M. SMITH. 1966. Fishes of the Tsitsikama Coastal National Park. National Parks Board of Trustees of the Republic of South Africa. 161 pp. SMITH, M.M. 1977. A new species of Argyrosomus (Pisces, Sciaenidae) from Natal, South Africa J. Zool., Lond. 181: 561-566. SMITT, F.A. 1982. Skandinaviens Fiskar. Vol. 1. P.A. Norstedt & Soners, Stockholm. 566 pp. STARLING, S. 1993. Fishing Australia: Fishing from Rocks and Beaches. Harper Collins, Australia. 96 pp. STEAD, D.G. 1906. Fishes of Australia. William Brooks & CO, Sydney. 278 pp. --- 1908. The Edible Fishes of New South Wales, their Present Importance and their Potentialities. New South Wales State Government, Sydney. 123 pp. STEINITZ, H. 1967. A tentative list of immigrants via the Suez Canal. Israel J. ZooI. 16: 166-169; STEINDACHNER, F. 1907. Fische aus Sudarabien und Sokotra. Denkschr. Akad. Wiss. Wien 48: 316-318. TANIGUCHI, N. 1969. Comparative osteology of the sciaenid fishes from Japan and adjacent waters-I. Neurocranium. Japan. J. Ichthyol. 16(2): 55-67. TANIGUCHI, N. & Y. OKADA. 1984. Identification of young of giant sciaenid and its morphological changes with growth. Japan. J. Ichthyol. 31(2): 181-187. TEMMINCK, C.J. & H. SCHLEGEL. 1842-1850. Pisces. In: P.F. von Siebold (ed.) Fauna Japonica. Lugduni Batavorum, Batavia. 323 pp. (for dates of publication, see Sherborn & Jentink, 1895). TENISON-WOODS, J.E. 1883. Fish and Fisheries of New South Wales. Thomas Richards, Sydney. 213 pp. TREWAVAS, E. 1966. Comments on the type-species of Sciaena Linnaeus, 1758. Z.N. (S.) 850. Bull. zool. Nora. 23: 4-5. 1973. Sciaenidae. pp. 396-401. In: J.C. Hureau & T. Monod (eds) Checklist of the Fishes of the North eastern Atlantic and of the Mediterranean. vol. 1: 1-677. UNESCO. Paris. --- 1977. The sciaenid fishes (croakers or drums) of the IndoWest-Pacific. Trans. Zool. Soc. Lond. 33(4): 253-541. VAN DER ELST, R.P. 1981. A Guide to the Common Sea Fishes of Southern Africa. C. Struik, Cape Town. 367 pp. VAN DER ELST, R.P., A. GOVENDA & N. DENTON. 1990. Biology and stock assessment of the squaretail kob Argyrosomus thorpei. In: Report on research during the period April 1989 to March 1990. Unpublished Report. Oceanographic Research Institute (Durban). 66: 49-59. WAITE, E.R. 1923. Fishes of South Australia. State Government, Adelaide. 243 pp. WALLACE, J.H., H.M. KOK & L.E. BECKLEY. 1984. Inshore small-mesh trawling survey of the Cape south coast. Part 2. Occurrence of estuarine related fishes. S. Afr. J. Zool. 19(3): 165-169. 40 Copyright 1995 J.L.B. Smith Institute of Ichthyology The following images related to this document are available:Line drawing images[fb95004t.gif] [fb95004q.gif] [fb95004w.gif] [fb95004a.gif] [fb95004z.gif] [fb95004r.gif] [fb95004d.gif] [fb95004x.gif] [fb95004k.gif] [fb95004v.gif] [fb95004o.gif] [fb95004l.gif] [fb95004b.gif] [fb95004e.gif] [fb95004u.gif] [fb95004i.gif] [fb95004p.gif] [fb95004j.gif] [fb95004c.gif] [fb95004f.gif] [fb95004h.gif] [fb95004y.gif] [fb95004n.gif] [fb95004g.gif] [fb95004m.gif] [fb95004s.gif] |
| |||||||||

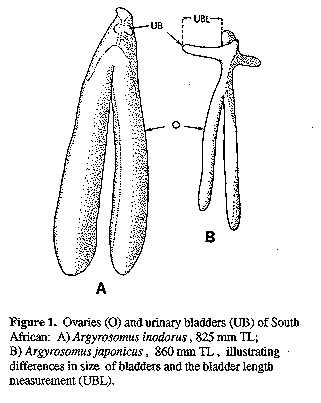{kind=link}
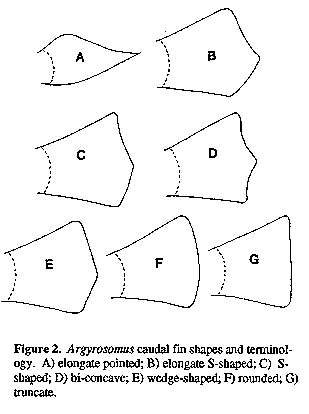{kind=link}
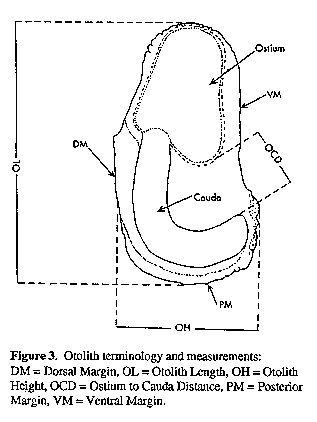{kind=link}
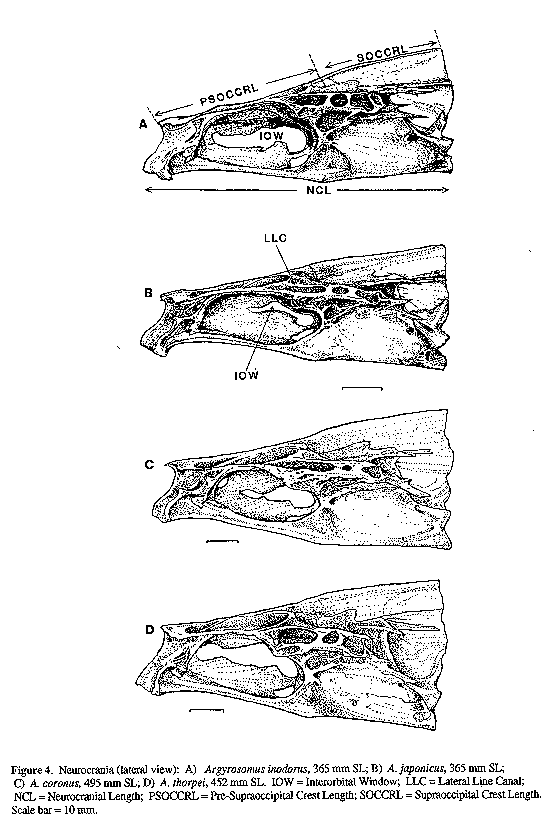{kind=link}
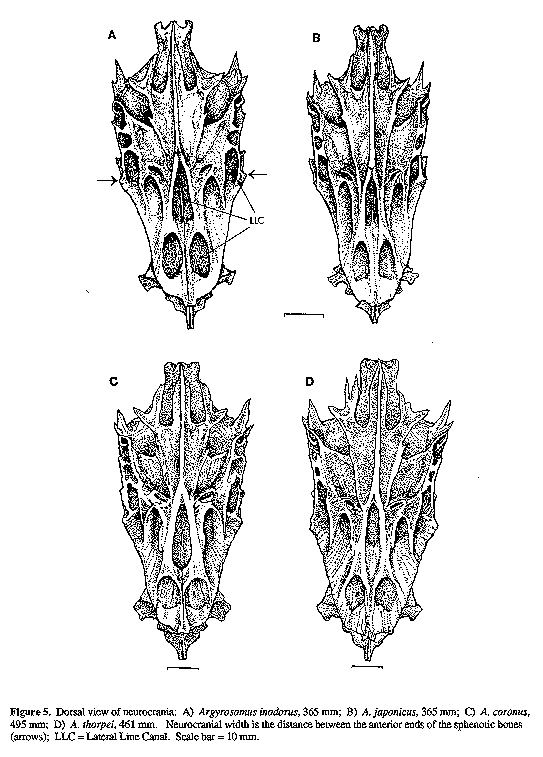{kind=link}
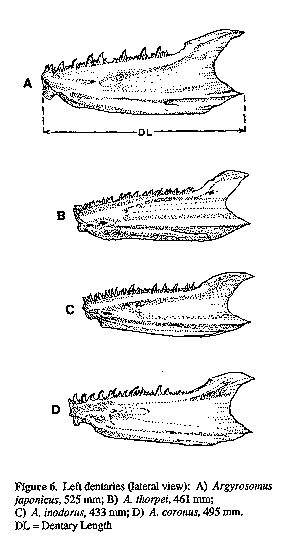{kind=link}
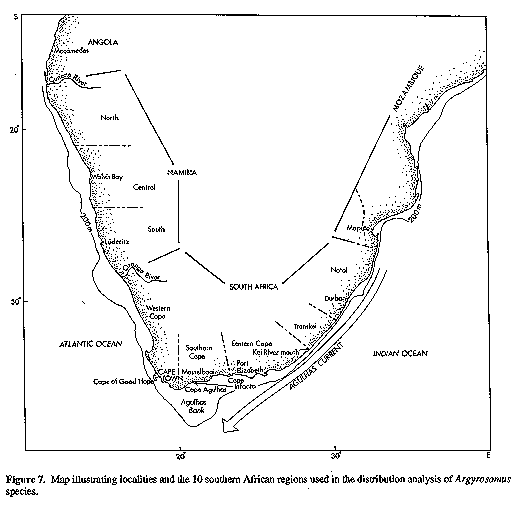{kind=link}
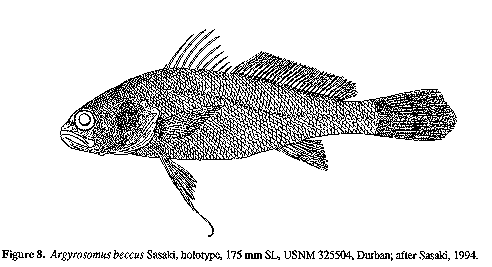{kind=link}
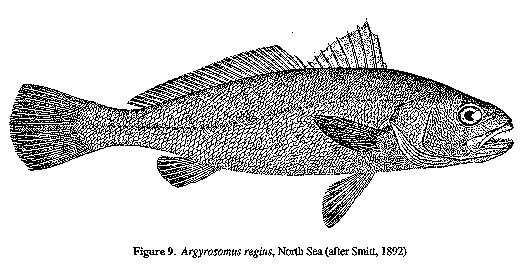{kind=link}
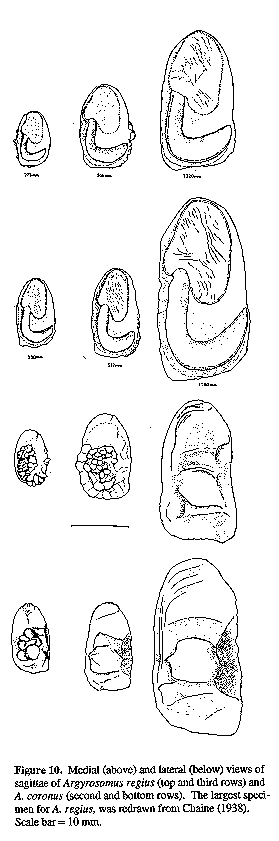{kind=link}
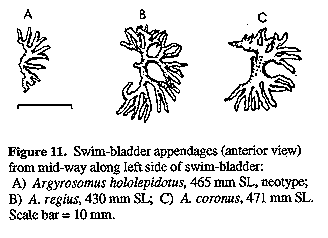{kind=link}
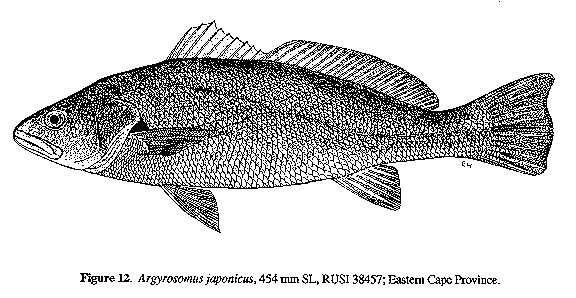{kind=link}
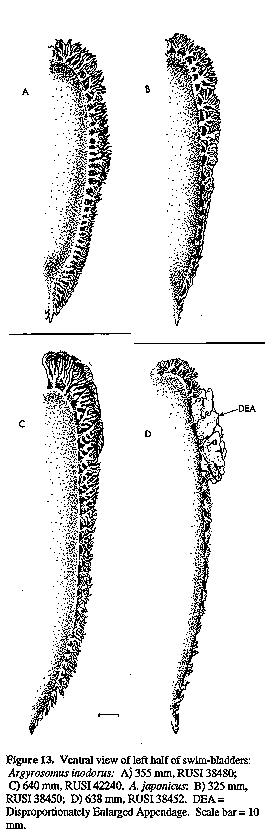{kind=link}
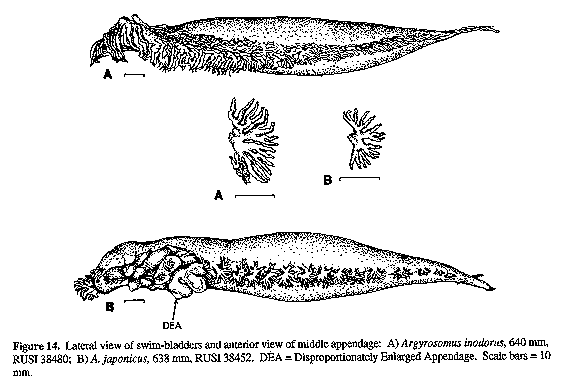{kind=link}
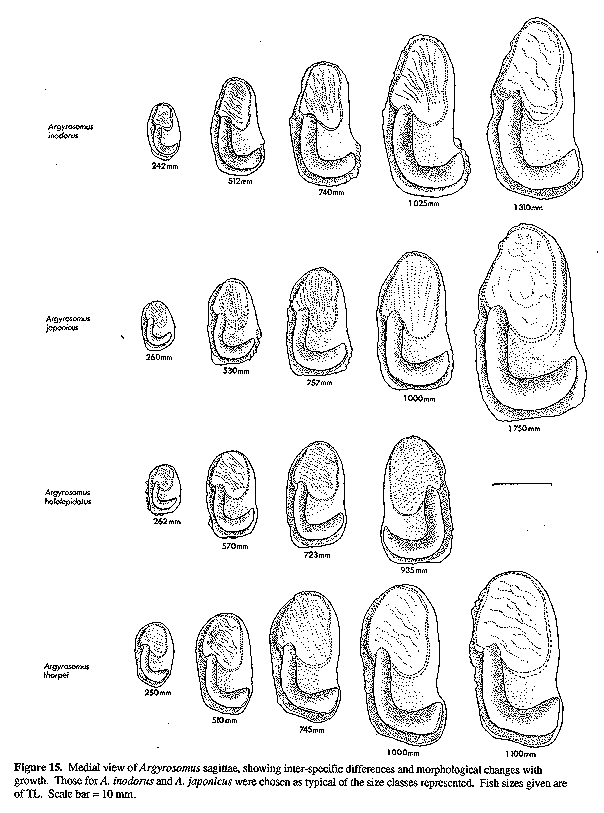{kind=link}
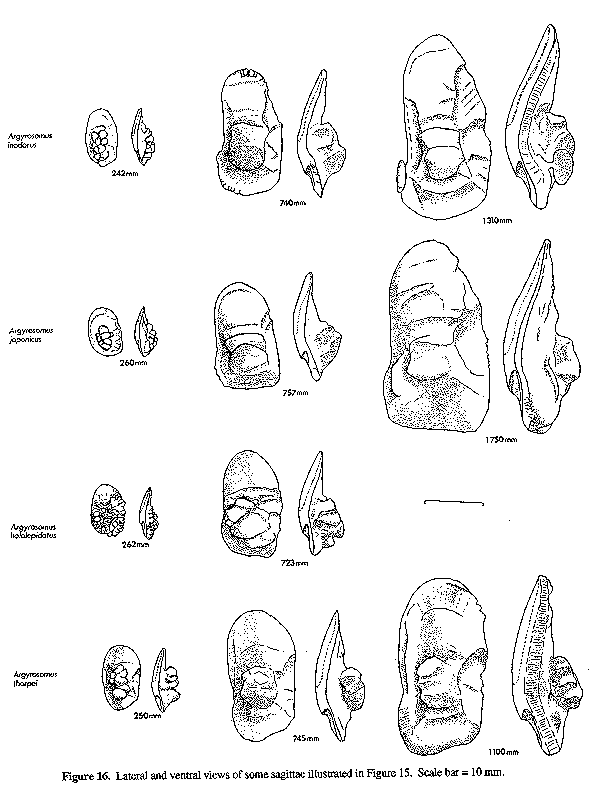{kind=link}
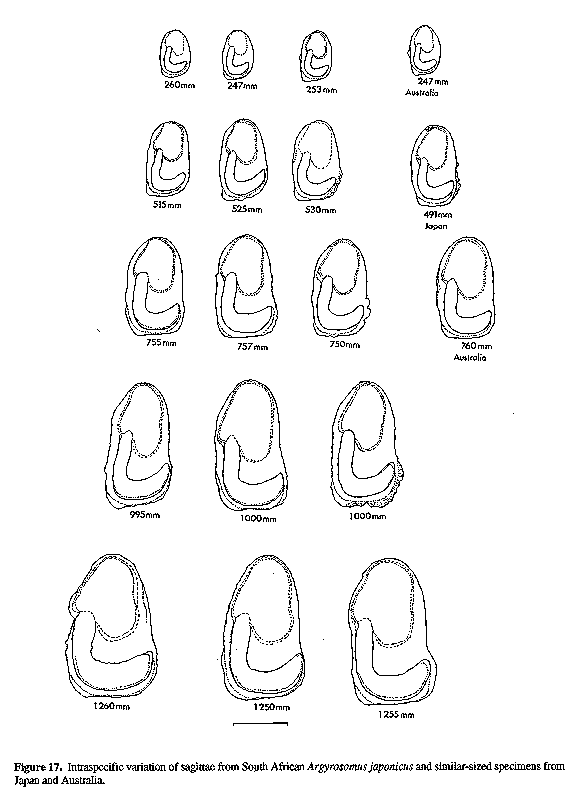{kind=link}
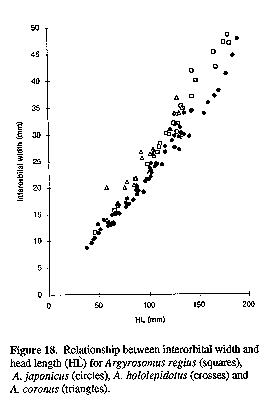{kind=link}
{kind=link}
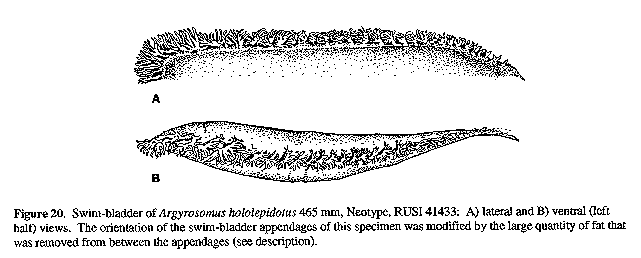{kind=link}
{kind=link}
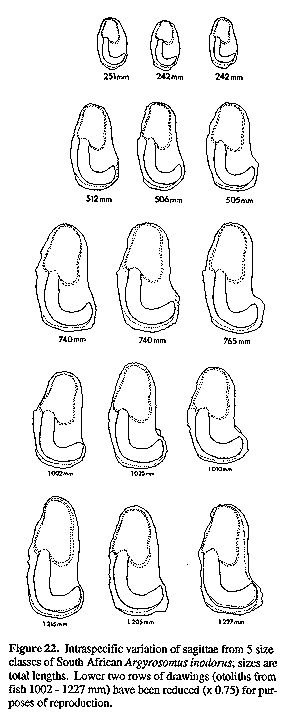{kind=link}
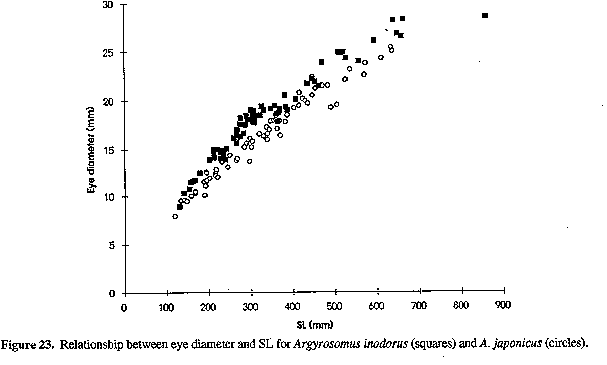{kind=link}
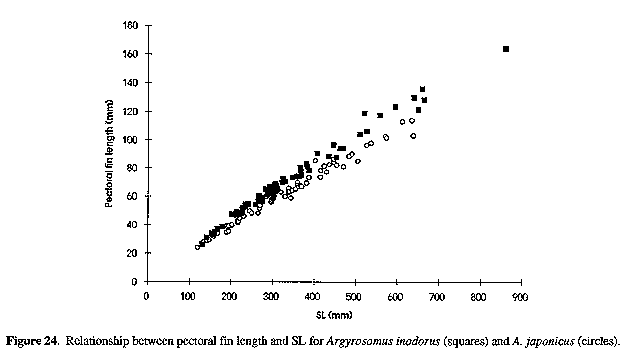{kind=link}
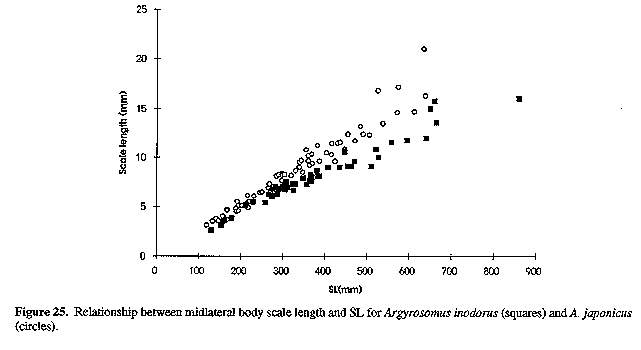{kind=link}
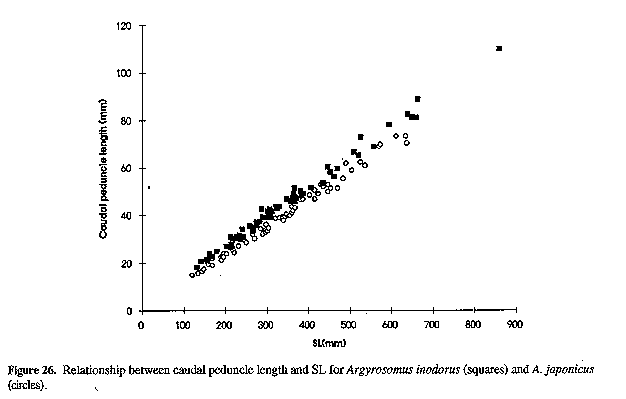{kind=link}
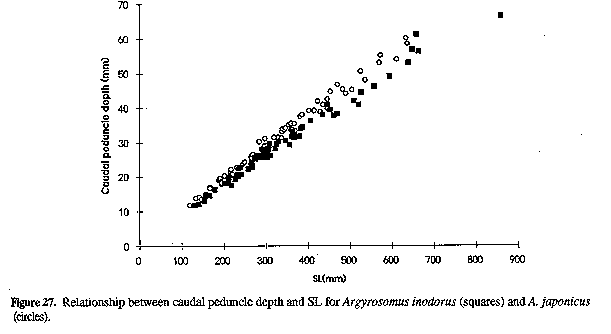{kind=link}
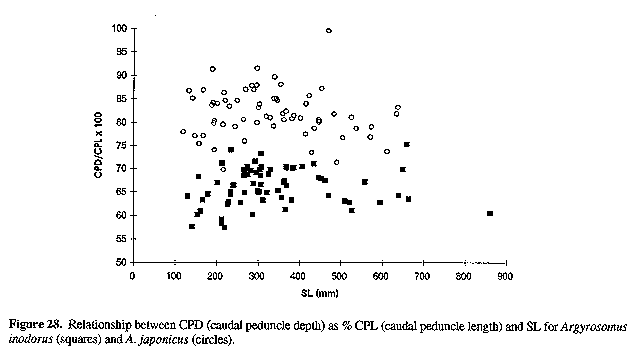{kind=link}
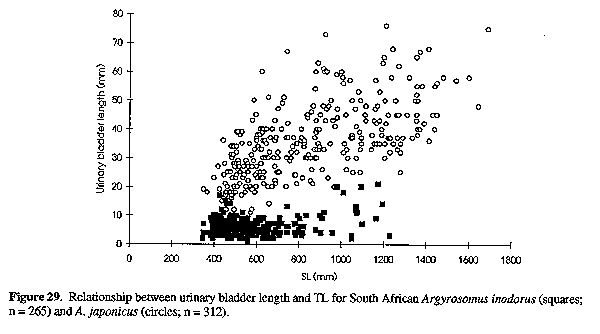{kind=link}
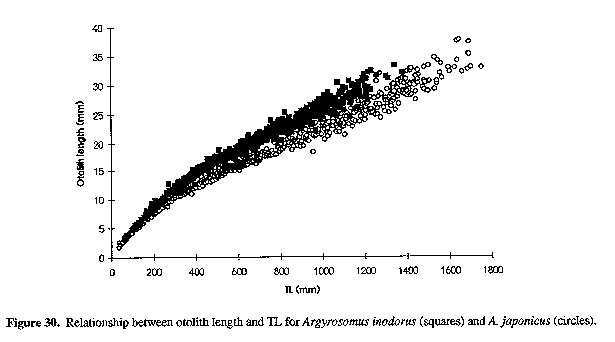{kind=link}
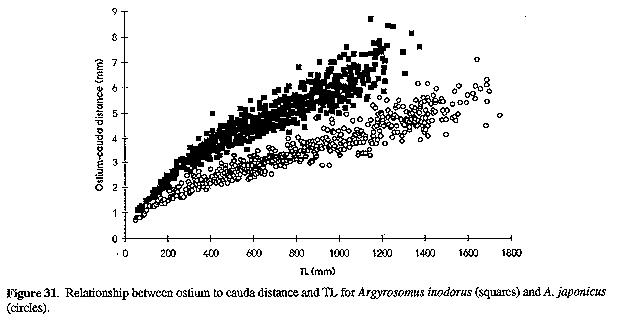{kind=link}
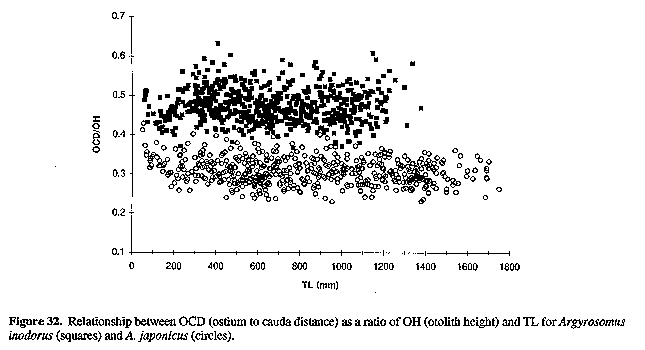{kind=link}
{kind=link}
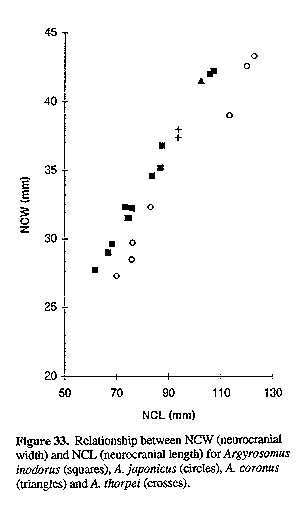{kind=link}
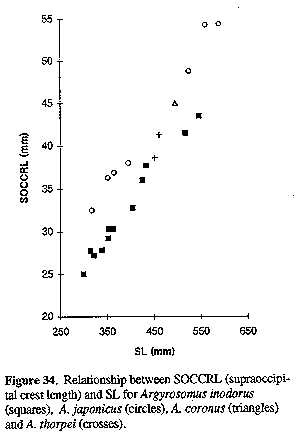{kind=link}
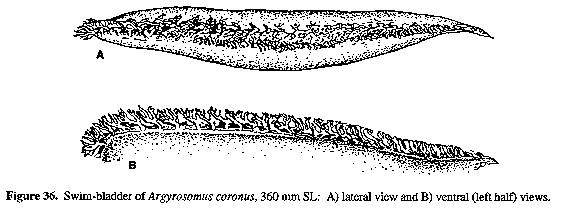{kind=link}
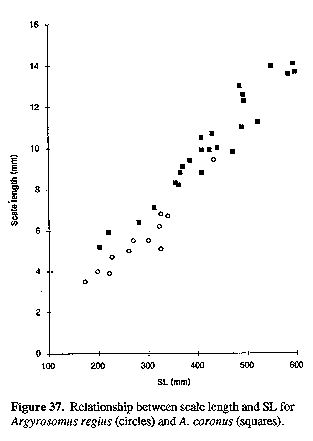{kind=link}
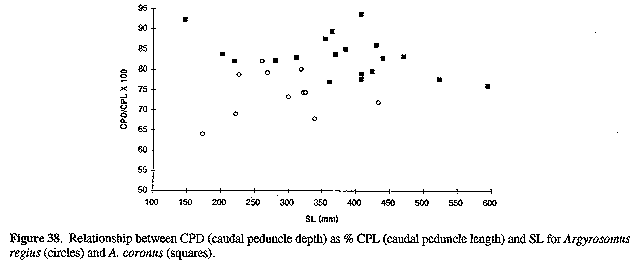{kind=link}
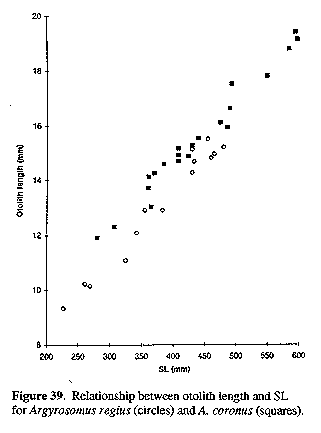{kind=link}
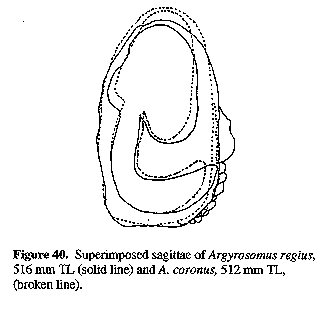{kind=link}
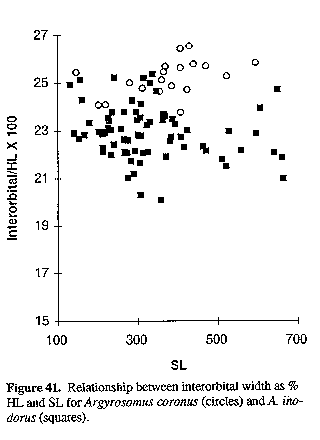{kind=link}
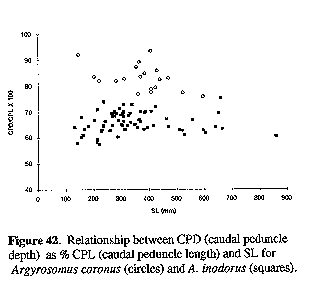{kind=link}
{kind=link}
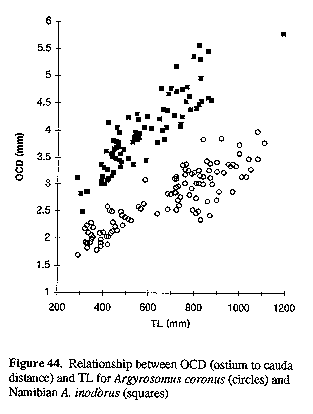{kind=link}
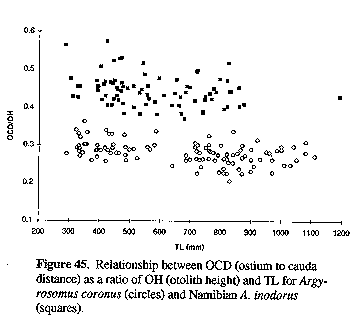{kind=link}